
Abstract
Infection of professional antigen presenting cells by viruses can have a marked effect on these cells and important consequences for the generation of subsequent immune responses. In this study, we demonstrate that different strains of bovine viral diarrhea virus (BVDV) infect bovine dendritic cells differentiated from nonadherent peripheral monocytes (moDCs). BVDV did not cause apoptosis in these cells. Infection of moDC was prevented by incubating the virus with anti-E2 antibodies or by pretreating the cells with recombinant E2 protein before BVDV contact, suggesting that BVDV infects moDC through an E2-mediated mechanism. Virus entry was not reduced by incubating moDC with Mannan or ethylenediaminetetraacetic acid (EDTA) before infection, suggesting that Ca(2+) and mannose receptor-dependent pathways are not mediating BVDV entry to moDC. Infected moDC did not completely upregulate maturation surface markers. Infection, but not treatment with inactivated virus, prevented moDC to present a third-party antigen to primed CD4+ T cells within the first 24 hours postinfection (hpi). Antigen-presenting capacity was recovered when viral replication diminished at 48 hpi, suggesting that active infection may interfere with moDC maturation. Altogether, our results suggest an important role of infected DCs in BVDV-induced immunopathogenesis.
Introduction
B
Infection of APC by viruses can have a marked effect on these cells and important consequences for the generation of the subsequent immune responses. Because maturation is essential for dendritic cell (DC) functionality, most viruses interfere with it. Experiments performed on human monocyte-derived DC (moDC) show that different viruses downregulate distinct stages and events of the maturation process (12). Thus, studying the interaction of DC with specific viruses is critical for understanding disease pathogenesis and immunity to infection.
In the early nineties, Sopp et al. proposed that the capacity of ncpBVDV to cause immunosuppression could be related to the tropism of the virus for the APC (42). There are only two reports on the interaction of BVDV with moDC, which differ in the type of cells used and the time-points analyzed. Recently, Rajput et al. reported that the ability of moDC to produce infectious virus was reduced with its differentiation from adherent monocytes to CD14−/CD205hi moDC (33) and a type 2 ncpBVDV downregulated, while a type-2 cpBVDV upregulated the expression of moDC cell surface markers major histocompatibility complex (MHC)-I, MHC-II, and CD86, but antigen presentation in these infected cells was not assessed. In 2003, Glew et al. demonstrated that monocyte-derived DCs are not susceptible to the cytopathic effect caused by cpBVDV, whereas monocytes (mo) are killed after infection (13). These authors showed that cpBVDV-infected moDCs (derived from nonadherent CD14+ monocytes) were not compromised in their capacity to stimulate proliferative responses in CD4+ T cells against a third-party antigen, 2 days after infection.
There is no information on the APC activity of moDC infected with BVDV at shorter time points. Early events of BVDV infection are important, as this virus rapidly gains access to lymph nodes draining the entry site (32) and deplete them from B and T cells and monocytes. Thus, evaluating the effect of BVDV on DC shortly after viral contact can help in understanding the role of these cells in defining the virus immunopathogenesis.
In this study, we studied the early interaction between BVDV and bovine moDCs. We demonstrate that BVDV infects moDC using the envelope glycoprotein E2 and produces new viral particles with a rapid kinetics, while infection impairs moDC maturation during the first 24 hours postinfection (hpi). Altogether, our results provide evidence on the role of infected DCs in BVDV-induced immunopathogenesis.
Materials and Methods
Cells and viruses
Madin-Darby bovine kidney (MDBK) cell line from the American Type Cell Collection (ATCC) was provided by the Institute of Virology (INTA) Argentina. Cells were cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% of gamma globulin-free, mycoplasma-tested, and virus-screened fetal calf serum (FCS) (HI FBS Qualified; Gibco). We also tested the serum for the presence of BVDV-RNA by RT-PCR and performed an ELISA for NS3 antigen and anti-NS3 antibodies (Priocheck BVDV-PI Plus; Prionics). Before use, the serum was heat inactivated at 56°C for 30 min. To control absence of live virus in the FCS, MDBK cells were cultured using 25% FCS (27). After three passages in MDBK cells, detection of BVDV antigens was performed using an FITC-conjugated polyclonal commercial antibody against BVDV (VMRD).
A cpBVDV SINGER Argentina strain genotype 1a [DQ088995] was used for all the experiments. To produce inactivated BVDV-1a-SINGER stocks (iBVDV), infection supernatants from cpBVDV were exposed to ultraviolet (UV) light at a distance of 20 cm for 30 min in an uncapped Petri-dish (41) and then stored at −80°C until used. Two other strains were used: a ncp 1b Argentinean field isolate 98/204 [JX848359] (31) and reference BVDV type 2 NY93 high-virulent strain [AF502399]. BVDV stocks were produced by infecting MDBK cells, following standard procedures (29). Titers were 1.45 × 107 for BVDV-1a-SINGER; 1.64 × 106 for BVDV-1b-98/204, and 6.55 × 106 for BVDV-2-NY93 stock.
Animals
Healthy adult bovines from INTA experimental farm, which were negative for BVDV, were used (tested with Priocheck BVDV-PI Plus; Prionics). Blood samples (∼150–200 mL) were obtained following national animal welfare regulations.
Antibodies
Anti-E2 monoclonal antibody (MAb) was provided by ICT Milstein-CONICET, Argentina (11). Anti-NS3 MAb was kindly provided by Dr. Gerrit Keizer (Prionics). Goat anti-mouse IgG conjugated to Alexa Fluor® 568 (Invitrogen) was used as secondary antibody in immunofluorescence assays. Serum form a BVDV-free bovine was used as negative control. For direct immunofluorescent staining of BVDV-infected cells, a polyclonal goat IgG anti-BVDV conjugated to FITC was used (VMRD). Anti-E2 sera from bovine and guinea pig were prepared using a recombinant purified E2 protein as reported before (11).
The following fluorochrome-conjugated monoclonal antibodies (AbDSerotec®) were applied in flow cytometry studies: anti-bovine CD11b-FITC (clone CC126), CD205-RPE (clone interleukin [IL]-A114); CD45-FITC (clone CC1), CD40-PE (clone IL-A156), CD86-FITC (clone IL-A190), CD1w2-PE (clone CC20); and anti-sheep MHC class II DQ DR polymorphic-PE (clone 28.1). Anti-bovine CD4-RPE and FITC (clone CC8), CD8-RPE and FITC (clone CC63), WC1-FITC (clone CC15), anti CD25-FITC (clone L-A111), and anti interferon-γ (IFN-γ) Alexa 647 (clone CC302) were used for the antigen presentation assay.
Differentiation of moDC
Immature moDCs were generated by culturing bovine monocytes as previously described (9,13). Briefly, peripheral blood mononuclear cells (PBMC) were isolated from BVDV-negative bovines by density gradient centrifugation as previously described (24). PBMC were then incubated with anti-human CD14-labeled superparamagnetic particles (Miltenyi-Biotech). Labeled cells were isolated from a Midimacs® column (Miltenyi-Biotech) according to the manufacturer's instructions. The purity in terms of CD14+ cells was evaluated by flow cytometry (≥92%), viability was over 95%. Cell suspensions were adjusted to 1 × 106 cells/mL, and nonadherent cells were incubated for 7 days (37°C, 5% CO2) in RPMI containing 10% BVDV-free FCS (already controlled for BVDV antigen, genome, and antibodies, as stated above), supplemented with 4 μL of bovine rGM-CSF/mL and 80 ng bovine rIL-4/mL (both cytokines purchased from AbD Serotec). Phenotype of adherent moDC was determined by flow cytometry using mouse anti-bovine CD11b-FITC and anti-CD205-RPE. More than 90% of the cells were CD11b+/CD205Low. Expression of CD14 was downregulated in differentiated cells (median fluorescence intensity ∼14; Table 1). When needed, moDCs were harvested with phosphate-buffered saline/ethylenediaminetetraacetic acid (PBS/EDTA) or treated in the culture plate, depending on the experiment. Alternatively, we infected adherent and nonadherent CD14+ cells and also differentiated and infected moDC from adherent CD14+-sorted monocytes following a published protocol (33).
moDC were infected, mock infected, or treated with iBVDV for 6 h (results were similar 12 hpi). After infection, cells were washed and left untreated or incubated ON in the presence of poly I:C or LPS. Cells were stained with different MAbs and analysis was performed on a FACS Calibur cytometer. Cells expressing CD11b were gated and the MFI of surface molecules CD1w2, MHC-II, CD40, CD86, CD14, and CD205 was determined. The average value from four separate experiments is shown.
Concentration of IL-10 in supernatants of moDC 6 h after each indicated treatment was estimated using a commercial ELISA. Values are expressed as mean ± SD from three experiments.
Significantly superior to nontreated cells within the same group (p < 0.05).
Significantly superior to noninfected untreated cells (p < 0.05).
BVDV, bovine viral diarrhea virus; hpi, hours postinfection; IL, interleukin; LPS, lipopolysaccharide; MAb, monoclonal antibody; moDC, monocyte-derived dendritic cell; MFI, median fluorescence intensity; MHC, major histocompatibility complex; ND, not done; Poly I:C, polyinosinic polycytidylic acid sodium salt, toll-like receptor ligand; SD, standard deviation.
Indirect immunofluorescent staining
moDCs were cultured in eight-well chamber slides (Lab-Tek; Nunc) at a concentration of 2 × 105 cells per well. Cells were incubated with infectious or inactivated BVDV-1a-SINGER at a multiplicity of infection (m.o.i.) of 4 for 3, 6, 12, and 24 h at 37°C, 5% CO2. After one wash with PBS, cells were fixed with PBS-3% formaldehyde for 20 min and permeabilized with PBS-0.5% Triton X-100 for 3 min. After a blocking step, anti E2 MAb 2.9.H (1:100 dilution) or anti NS3 MAb (1:400 dilution) was added followed by an Alexa Fluor 568 conjugate (1:2,000). Wells treated only with primary or secondary antibodies were included as controls. Cell nuclei were stained with 4′6-diamidino-2-phenylindole (DAPI; Sigma). After the staining, slides were assembled adding an antifade reagent (Vectashield® Mounting medium; Vector Laboratories). Images were captured using a fluorescence microscope (Leica™ Model TCS SP5).
BVDV titration on MDBK cells
Titers of extracellular virus were obtained by microtiter assay as previously described (4,11,29) with some modifications. Briefly, MDBK cells were grown in 96-well plates (Corning® Costar®) to a confluence of 80% (104 cells per well). Serial fourfold dilutions of the tested samples were made in DMEM (Gibco) and 50 μL added to duplicate wells. Plates were incubated for 5 days at 37°C, 5% CO2. Then, cells were fixed with 3% PBS-formaldehyde. Each well was treated with 0.1 M glycine in PBS followed by 0.2% Triton X-100. Presence of BVDV was revealed with an anti-BVDV polyclonal antiserum conjugated to FITC (VMRD). Infected cells were observed in an inverted fluorescence microscope (Olympus IX71). Titers were calculated as median tissue 50% culture-infective dose per milliliter (TCID50%/mL).
Virus infection studies
Virus growth curves were assayed at the same m.o.i. of 4 for the three virus strains. moDCs were plated by quadruplicates in 96-wells plates (1 × 105 per well) and infected or mock infected with 100 μL of BVDV in culture medium (RPMI) without FCS for 1 h following standard procedures. After adsorption of the virus, the inoculum was removed by washing the cells thrice with PBS 1× prior the addition of RPMI with 10% FCS. The last wash (performed with RPMI-10% FCS) was taken as time 0 postinfection. Supernatants were removed from cultures of MDBK and moDC at 0, 3, 6, 18, and 24 h time intervals up to 4 days. Paired monolayers were collected, pelleted, and frozen at −80°C. Samples were then processed for RNA extraction or NS3 ELISA.
Adherent and nonadherent CD14+-sorted monocytes were cultured in 96-well plates (1 × 105 per well) with GM-CSF and IL-4 (AbD Serotec) and infection with BVDV-1a-SINGER or NY-93 (m.o.i. = 4) was performed at different times of cytokine culture. Cells were infected 0, 48, and 96 h, and at 120 or 168 h in culture for adherent and nonadherent monocytes, respectively. Cells were harvested at 18 and 48 hpi and processed for NS3 ELISA (see below).
To evaluate possible virus entry mechanisms, moDC cultures were treated with mannan (10 mg/mL), truncated E2 glycoprotein expressed by recombinant baculovirus (rBtE2) (0–10 μg/mL), and EDTA (5 mM) or mock treated (RPMI), and virus was pretreated with bovine normal serum (1:30), anti E2 serum from bovine or guinea pigs (1:30), or mock-treated for 1 h at 37°C. These treatments have been previously done to viral infection with BVDV-1a-SINGER or BVDV-1b-98/204 strain. NS3 detection by ELISA or titer of extracellular virus was determined.
NS3 ELISA
For NS3 determination, MDBK and moDC were harvested, washed, and mixed with the lysis buffer (PrioCHECK® BVDV-Ag PI focus; Prionics AG). Total protein content was quantified using the Protein Assay Kit (Pierce® BCA; Thermo Scientific). Suspensions containing 0.22 μg of total proteins in a volume of 100 μL were used for NS3 detection by PrioCHECK® BVDV-Ag PI focus ELISA following the manufacturer's instructions. Plates were read in a MultisKan FC (Thermo Scientific) ELISA reader. Results were expressed as OD values (450 nm).
Flow cytometry analysis
Expression of activation markers in stimulated moDC was determined by flow cytometry as previously described (13). Briefly, CD11b+ moDCs were stimulated with infectious or iBVDV (m.o.i. = 4). Cells stimulated with 1 μg lipopolysaccharide (LPS)/mL (Escherichia coli O55:B5 LPS; Sigma) or 5 μg poly I:C/mL (Sigma) and nonstimulated cells were used as positive and negative controls, respectively. Treatments and controls were incubated at 37°C, 5% CO2 for 6 h. Cells were blocked with PBS-2% FCS at 4°C for 30 min and then stained for 30 min at room temperature with the corresponding antibodies. Cells were washed thrice with PBS-2% FCS and then fixed with PBS-4% paraformaldehyde. Stained cell populations (gated in CD11b; 10,000 events) were analyzed using a FACScalibur™ flow Cytometer and CellQuest™ Software v3.1 (BD Biosciences).
Annexin V apoptosis assay
After treatment with infectious or inactivated BVDV, cell viability in moDC was determined using a PE-Annexin V Apoptosis Detection Kit I (BD BioSciences) according to the manufacturer's instructions. Briefly, stimulated moDC were harvested, washed twice with cold PBS, and the concentration was adjusted to 1 × 106 cells/mL. Nonstimulated cells were used as control. Aliquots of 100 μL (1 × 105 cells) were stained with 5 μL PE-Annexin V and 5 μL 7-Aminoactinomycin D (7-AAD), mixed, and incubated at room temperature for 15 min in the dark. Before the cells were analyzed by flow cytometry, 400 μL binding buffer (0.1 M HEPES/NaOH [pH 7.4], 1.4 M NaCl, and 25 mM CaCl2) diluted at 1:10 was added to the cell samples.
Assessment of IL-10
IL-10 production was measured in BVDV-treated and mock-treated moDC at 6 and 24 hpi. Total mRNA was extracted from moDC using the Qiagen RNeasy Mini Kit (Qiagen) and IL-10 mRNA levels were semiquantified by qRT-PCR as described before (24). Concentration of IL-10 was also determined in supernatants (1 μg of total proteins) of cultured moDC using a commercial ELISA (CUSABIO Bovine Interleukin 10 ELISA Kit) following the instructions of the manufacturer. Plates were read in a MultisKan FC ELISA reader (OD = 450 nm).
Antigen presentation assays
Two antigen-specific assays were performed. PBMC for both moDC generation and lymphocyte isolation were obtained from BVDV-negative cattle from INTA's experimental field. The animals enrolled in these experiments had been vaccinated at least thrice with a tetravalent foot and mouth disease virus (FMDV) commercial vaccine and had received the last vaccine dose about 30 days before our experiments were performed.
In a first experiment, moDC were differentiated from CD14+ cells as detailed above and incubated with BVDV (live or UV-inactivated) (m.o.i. = 4) for 6 h and washed as described above. These cells were then treated or not with 10 μg of inactivated sucrose gradient-purified FMDV 140S particles (A 24/Cruzeiro strain), for 18 h (23). These moDCs were then treated with 25 μg/mL of mitomycin C following standard protocols (16) and cocultured with a T-cell-enriched PBMC suspension obtained from the same animal. This preparation was obtained by isolation of PBMC from whole blood by density gradient centrifugation (Histopaque, density 1.083 g/mL; Sigma), depletion of CD14+ cells by magnetic separation, and exclusion of plastic-adherent cells. Cell suspensions were identified as 95% T cells by flow cytometry staining with anti CD25 and also CD8, CD4, and WC1 antibodies. The syngeneic coculture also included a control with pokeweed mitogen (PWM–Phytolacca americana; Sigma). After 18 h of incubation, CD4+/IFN-γ+ cells were determined by flow cytometry as already described (24).
In a second experiment, moDC from three different bovines were treated with live BVDV for 6, 24, and 48 h before the FMDV uptake and coculture with T-cell-enriched PBMC. T-cell activation was determined by measuring IFN-γ production using a commercial ELISA (ID Screen® Ruminant IFN-γ, ID Vet) following the manufacturer's instructions.
Statistical analysis
Results between different experimental groups were compared by ANOVA two-factor repeated measures followed by Bonferroni multiple comparisons test. When comparing two groups, Mann–Whitney test was used. The confidence interval was 95% or 99%, depending on the experiment. Statistical analyses were carried out using GraphPad Prism v5.0 (GraphPad Software).
Results
BVDV infects and replicates in differentiated moDC
moDC differentiated from nonadherent CD14+ monocytes were treated with infectious (BVDV) or UV-inactivated cpBVDV (iBVDV) at high multiplicities of infection (m.o.i. = 4), and the presence of intracellular viral particles was determined by immunofluorescence staining using a MAb against the virus envelope glycoprotein E2 and nonstructural protein 3 (NS3), used as a marker of infection (13). Results are shown in Figure 1. We verified that almost all the infected moDC stained positive for E2 (80% per field) and about 40% per field stained positive for NS3 at 6 hpi (Fig. 1A). Staining with the anti-NS3 MAb at 6 hpi indicated that the viral RNA was rapidly translated in the infected cells (Fig. 1B). Cells treated with the UV-inactivated virus stained positive with the anti-E2 MAb, demonstrating that the moDC incorporated the inactivated viral particles, and stained negative with anti-NS3 MAb, as expected (Fig. 1B).
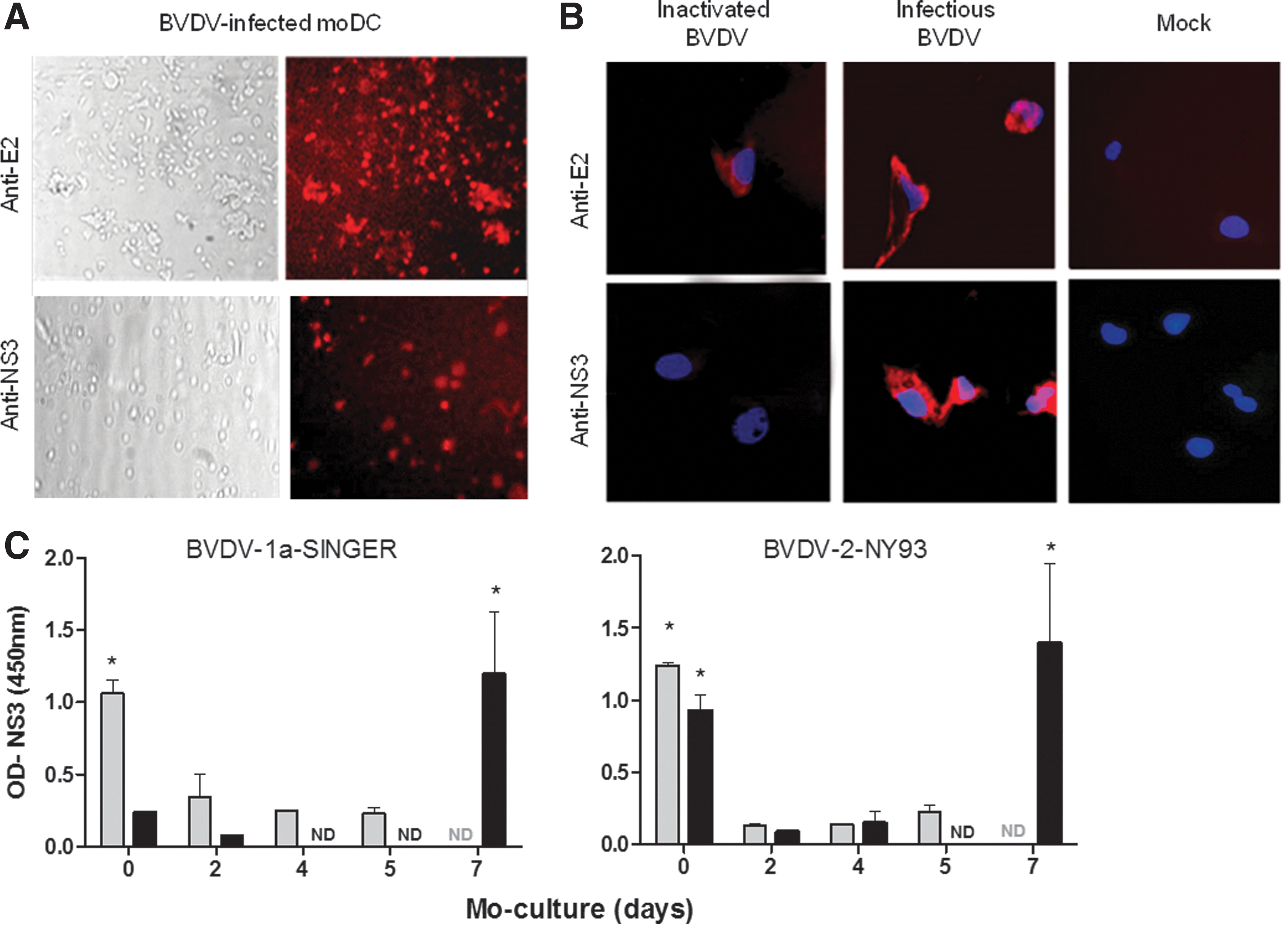
A previous report by Rajput et al. (33) demonstrated a reduced capacity of moDC to produce infectious virus compared to monocytes. These authors used adherent CD14+ cells sorted from PBMC, while we differentiated moDC from nonadherent precursors according to the protocol reported by Glew et al. (13). Thus, in an approach to figure out if there is a difference in BVDV infectivity in these cells, we evaluated both adherent and nonadherent CD14+ precursors in terms of susceptibility to BVDV infection along the differentiation process from monocytes to moDC.
CD14+ PBMC-sorted precursors were cultured with GM-CSF and IL-4 and infected with BVDV-1a-SINGER or BVDV-2-NY93 strains (m.o.i. = 4) at different times along the differentiation process (Fig. 1C). Cells were harvested at 18 hpi and tested for NS3 by ELISA. Adherent monocytes supported replication of both strains (0 days of culture), while nonadherent monocytes were infected only with the NY93 strain. Adherent monocytes, however, lost infectivity with both strains as they differentiated to moDC. Conversely, moDC differentiated from nonadherent monocytes yielded high NS3 levels at 7 days of culture for both virus strains. These results indicate that moDC derived from either monocyte population may correspond to different lineages and they do not behave equally in terms of supporting BVDV replication.
Replication kinetics of different BVDV strains in moDC
We then studied the replication kinetics (one cycle; m.o.i. = 4) of BVDV-1a-SINGER, BVDV-2-NY93, and a field ncp isolate (BVDV-1b-98/204) in moDC derived from nonadherent CD14+ PBMC. Supernatants were harvested at different time points between 0 to 144 hpi and evaluated by titration on MDBK cells (Fig. 2A). Pellets were lysed to obtain total proteins and NS3 was detected by ELISA (Fig. 2B). The experiment was run four times, using four independent moDC preparations. To confirm the absence of viral particles in the supernatant after removing the virus inoculum, cells were washed thoroughly, and the media recovered after the last washing step was used as the inoculum for time 0 dpi. qRT-PCR performed on this sample indicated that we had undetectable levels of viral RNA (not shown).
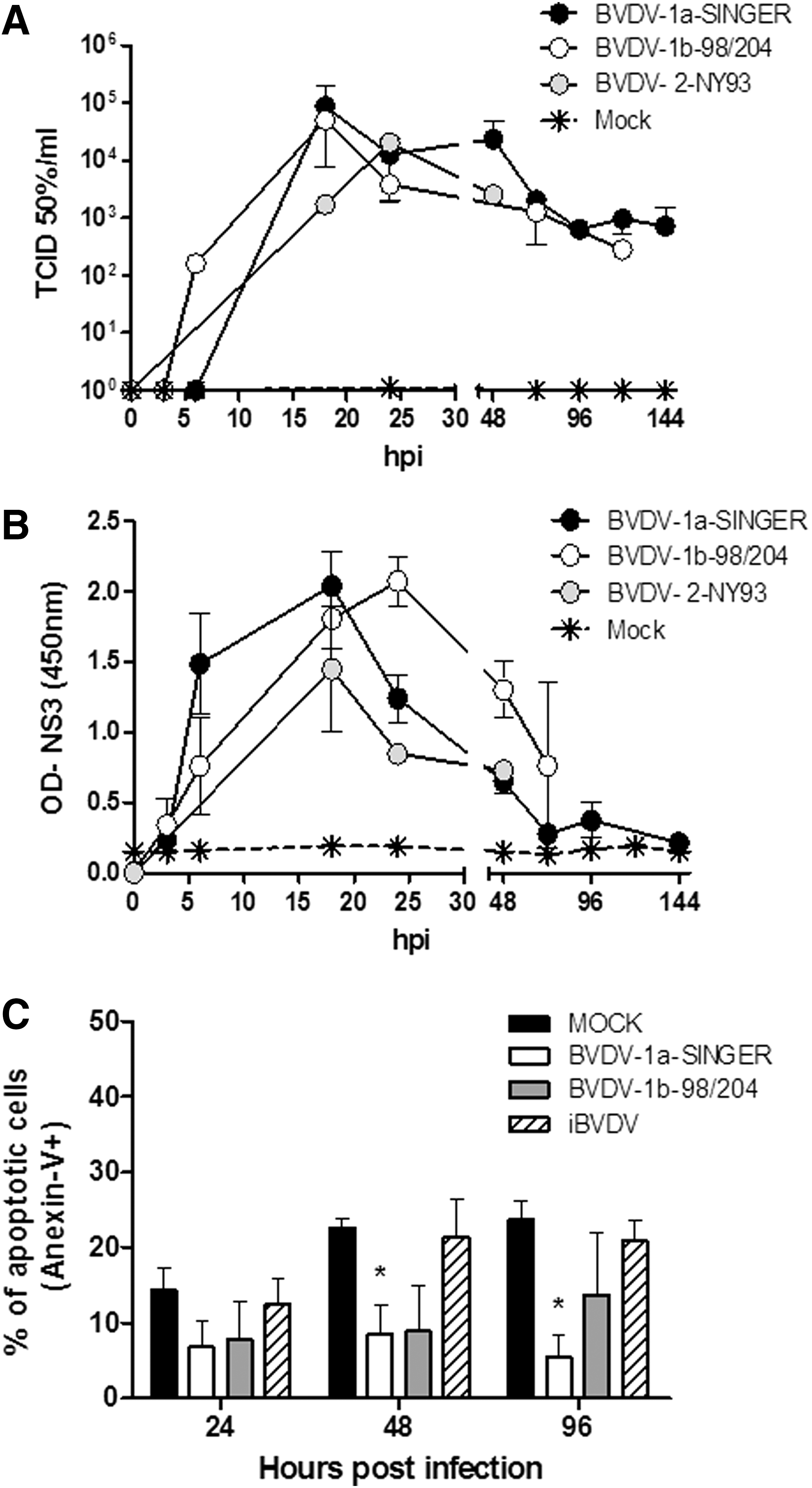
New viral particles collected from the supernatants were produced in moDC as soon as 6–18 hpi, with the highest titers (measured in MDBK cell culture) at 18 hpi for both type 1 strains. This is an accelerated kinetics compared to one-cycle infection of MDBK cells, on which similar titer levels (∼105 TCID50%/mL) are obtained at 72 hpi (data not shown). Peak titers for BVDV-2-NY93 strain were detected at 24 hpi. Titers ranging ∼103–104 TCID50%/mL were maintained from then on within similar levels until the end of the experiment (144 hpi) for the three strains. No cytopathic effect was visualized in ncp or cpBVDV-infected moDC cultures along the experiment.
We also followed the replication kinetics by detection of intracellular NS3 in moDC infected with the three strains (Fig. 2B). A sharp increase in NS3 values was detected at 6 hpi in BVDV-1a-SINGER-infected moDC that peaked at 18 hpi. Cells infected with BVDV-2-NY93 strain also showed peak NS3 production at 18 hpi. Conversely, BVDV-1b-98/204 strain had a delayed peak at 24 hpi. NS3 values decayed gradually after the peak for the three strains.
BVDV-infected moDC do not undergo apoptosis
No morphological differences were visualized between BVDV-infected, iBVDV-treated, or noninfected moDC, even when a cytopathic strain was tested. When these virus stocks were tested in PBMC, they all induced apoptosis (not shown) and the BVDV-1a-SINGER showed a cytopathic effect in MDBK cells.
We then analyzed the apoptosis/necrosis rate of infected moDC with Annexin V/7AAD. Type 1 strains were used in this experiment, together with inactivated virus. Results were similar for the ncp-type 2 virus (data not shown). Mock-treated moDC showed a low spontaneous apoptosis rate (∼14%, Annexin V+; 7AAD− cells) 24 hpi, day 8 after the differentiation treatment (Fig. 2C). Similar levels were found in those cells treated with inactivated virus at 24 hpi (% of Annexin V+ cells: 12.5 ± 3.5). These values increased to ∼23% along the experiment. In all three experiments, while mock infected or iBVDV-treated moDC had similar percentages of apoptotic cells at the different points tested, this number was reduced over time in infected cells. The reduction in the number of apoptotic cells became evident at 48 and 96 hpi for both virus strains. Significant lower numbers of apoptotic moDC were found at these time points for the BVDV-1a-SINGER strain (p < 0.05), while the tendency was similar for BVDV-1b-98/204, differences were not statistically significant.
E2 glycoprotein mediates BVDV infection of moDC
Viruses can enter DCs taken by the phagocytic pathway or by scavenger receptors, by an FcR-mediated mechanism (FMDV) (36), through lectin binding, or by interacting with a specific virus receptor (40). In a first screening, we incubated moDC before infection with Mannan, a bacterial polysaccharide that binds with high affinity to the mannose receptor, and EDTA, to assess calcium dependency of endocytosis. We used high concentrations, reported before to inhibit the entry of other viruses (40). Considering that BVDV E2-envelope glycoprotein is a high-mannose protein, we hypothesized that this high-density envelope protein might be a candidate to bind lectins on moDC. A recombinant truncated E2 protein (rBtE2) (11) was also tested as a putative mediator of specific viral entry. This protein used at 10 μg/mL, a concentration previously found to completely inhibit BVDV infection in susceptible cell lines (18). Infection proceeded for 18–20 h, when supernatants were collected for titration on MDBK cells and pellets were processed for NS3-ELISA.
Pretreatment of mo-DC with Mannan or EDTA did not reduce virus entry to DC (Fig. 3A, B). Conversely, preincubation with rBtE2 completely reduced viral yield. The specificity of E2 recognition was confirmed by diluting purified rBtE2. Infectivity increased as rBtE2 concentrations decreased. Blocking of BVDV infection followed a dose/response curve, as NS3 translation and titer of newly produced virus diminished with the concentration of rBtE2 (Fig. 3C). Fifty percent inhibitory concentration (IC50) was 0.5 μg/mL for both virus strains; a similar value was reported working with MDBK cells (18). These data indicated that infection of moDC was associated to the capacity of viral E2 glycoprotein to be recognized by moDC.
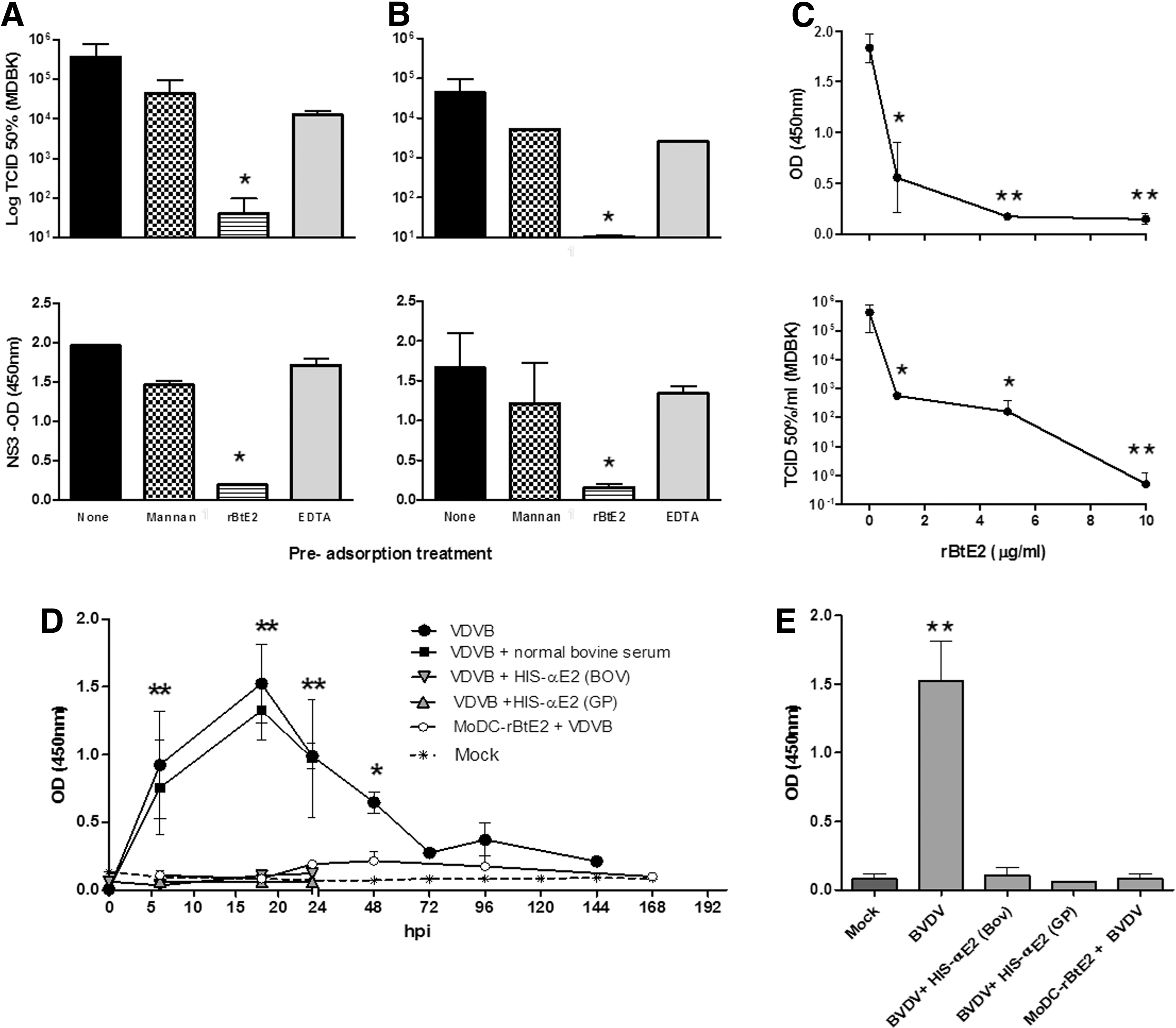
moDC cultures were treated with mannan (10 mg/mL), rBtE2 (10 μg/mL), EDTA (5 mM), or RMPI (“None”) for 1 h, washed, and infected with BVDV-1a-SINGER
To verify this finding, we infected moDC with the viral inoculum preincubated with an anti-E2 bovine serum. A normal bovine (preimmune) serum was included as control. Infection was measured by NS3 detection (Fig. 3D, E). Pre incubation of moDC or the virus with normal bovine serum did not prevent virus infection. Anti-E2 sera completely blocked BVDV from infecting moDC. To test the absence of any Fc-mediated viral entry, we preincubated BVDV inoculum with guinea pig anti-E2 antibodies. Both sera prevented BVDV infection both in MDBK and moDC. In both cases, NS3 could not be detected, showing that viral RNA was not translated. Again, normal bovine, or guinea pig sera did not interfere with BVDV infection. Preincubation of moDC with rBtE2 completely blocked BVDV infection, showing that the virus entered the cells, although an E2-specific mechanism.
BVDV infection prevents the maturation of moDC
The effect of BVDV infection on moDC maturation was studied by measuring the expression of costimulatory molecules CD86, CD40, as well as MHC-I (CD1w2) and II by flow cytometry. Determinations were done 6 h after the different treatments, as assessments performed at 24 hpi were quite variable between the different moDC preparations (data not shown). Infection with BVDV-1a-SINGER and incubation with iBVDV did not change the expression of any of these markers compared to noninfected moDC (Fig. 4 and Table 1). In some experiments, MHC-II expression was reduced when compared to mock-treated cells, however, differences were not significant. No changes were observed for CD205 expression in all treatments (Table 1). Only the infective virus induced the expression of IL-10 at 6 hpi, measured both at mRNA level (by qRT-PCR, showing a sevenfold increase with respect to noninfected cells, data not shown) and as secreted protein quantified by ELISA (Table 1).
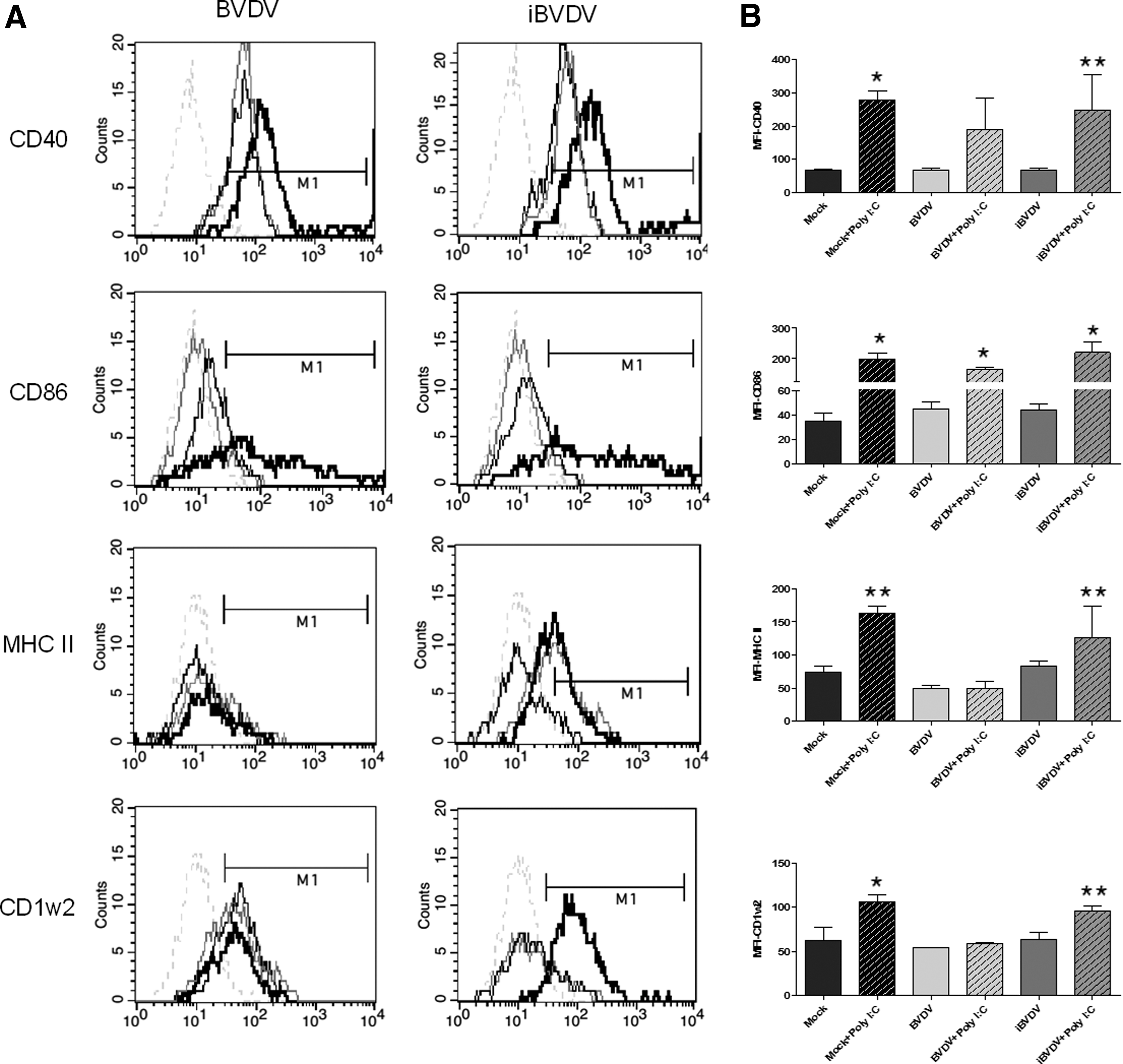
moDC seemed to be arrested in an immature state by the presence of either infectious or inactivated virus shortly after treatment. Previous studies have demonstrated that human moDC could be matured by proinflammatory cytokines, poly I:C, or LPS (19,39). For instance, infection of porcine moDC by classical swine fever virus (CSFV) did not influence the expression of maturation markers and they could be activated by different activation “cocktails” (i.e., TNF-α/IFN-γ) (7). We applied a similar approach to evaluate whether we could induce maturation of BVDV-infected or iBVDV-treated moDC. Cells that had undergone both treatments for 6 h were subsequently incubated with poly I:C or LPS for another 18 h when costimulatory molecules were measured by flow cytometry.
Upregulation of range of all the surface molecules involved in antigen presentation, typical of moDC maturation, was observed after incubation with poly I:C and LPS in those cells treated with the inactivated virus (iBVDV), with comparable levels to those of noninfected cells (Fig. 4 and Table 1). Expression of MHC-I (CD1w2) and II remained downregulated when moDC had been pretreated with infectious BVDV. The same results were obtained for the type 1b strain (Table 1). Only CD86 expression was recovered after poly I:C treatment in BVDV-infected moDC (Fig. 4 and Table 1). Infected moDC seems to recover CD40 expression after poly I:C incubation, although results were quite variable. Although in some experiments we found evidence of upregulation, overall results showed that differences to mock-treated cells were not statistically significant (p > 0.05). Similar results were obtained when LPS was used to induce moDC maturation (Table 1).
Functionality of the infected moDC
We then evaluated the influence of virus infection on an MHC-restricted antigen-specific T-cell response against a third-party antigen. We used a well-characterized model of FMDV presentation to T cells. APC treated with inactivated FMDV are able to present viral antigens to FMD vaccine-primed T cells inducing the production of IFN-γ mainly by CD4+ T cells (6,30). We used this system to study the capacity of BVDV-infected moDC to present antigens to primed T cells. For this purpose, moDCs were pretreated with iBVDV or infectious BVDV (1a-SINGER and 1b-98/204), or mock-infected for 6 h, and then incubated with inactivated FMDV 140S-purified particles. After these treatments, moDC were fixed and cocultured with autologous T-cell-enriched PBMC. We then tested the capacity of treated moDC to stimulate IFN-γ production by CD4+ T cells, which was identified by flow cytometry (Fig. 5A). As a positive control, T cells were treated with PWM, and all of them produced high levels of IFN-γ independent of the moDC they were in contact with, indicating that these cells were viable and metabolically active.

Noninfected moDC as well as those pretreated with iBVDV and pulsed with FMDV-140S particles were able to stimulate the production of IFN-γ by CD4+ T cells; differences to mock-treated moDC were significant (p < 0.05). However, BVDV-infected moDCs were unable to present FMDV antigens to primed T cells, as no differences were found with mock-treated cells. Results were similar for both strains (Fig. 5A).
This first experiment used moDC at 6 hpi, during active BVDV replication. We then investigated if these cells could recover their capacity to uptake and present FMDV antigens when moDC production of new BVDV particles has diminished. Therefore, we measured the production of IFN-γ in the supernatants from T lymphocyte-enriched PBMC, which have been in cocultured with FMDV-loaded moDC. These moDC had previously been in contact with BVDV or iBVDV for 6, 24, or 48 h. Mock-infected cells were used as controls; results are shown in Figure 5B. BVDV-infected moDC could not recall FMDV-specific T-cell responses at 6 and 24 hpi; however, they were able to efficiently present FMDV antigens at 48 hpi. These data indicate that BVDV-infected moDC were functionally intact with respect to their capacity to trigger anamnestic T-cell responses, but this activity was restricted during active virus replication.
Discussion
DCs are specialized APC that are fundamental to initiate both immunity and tolerance (34). The flexibility of DCs or macrophages to adapt to the environment and respond accordingly can be hijacked by pathogens for their own interest to transform a potentially immunogenic APC into a tolerogenic cell, with clear consequences in pathogen clearance. DC can also be involved in the propagation of viral infection; consequently, they may contribute to the pathogenesis of viral diseases (21). During the last 25 years, many studies have addressed the relationships between several viruses and DC, however, only a few ones evaluated the interaction between BVDV and bovine DC in terms of infectivity and effect on their maturation (5,13,33).
In this study, we demonstrated that different BVDV strains can infect CD11b+ moDC. One cycle of infection produced infectious viral particles as soon as 6–24 h after adsorption. This is an accelerated kinetics compared to MDBK infection, reported to peak at 48 hpi for the 1a-SINGER strain (29). Kinetics of BVDV infection in moDC was monitored by titrating virus from supernatants (assessed in MDBK cells) and intracellular production of NS3. This is the first description of the kinetics of BVDV-SINGER type 1a strain growth in mature CD11b+ moDC.
Our results are in agreement with those published by Glew et al., who showed that CD11b+ moDC support BVDV replication and produce infectious virus (13). However, Rajput et al. recently demonstrated that moDC precursor monocytes produced infectious BVDV, while differentiated moDC failed to do so (33). This disagreement can be due to the differences in the moDC used in both studies. We differentiated moDC from nonadherent CD14+ monocytes, while Rajput et al. used adherent cells as precursors. We have performed our infection experiment using moDC derived from adherent monocytes and obtained similar results as Rajput et al. (Fig. 1C). This evidences that both cell lineages are different. Recently, two phenotypically distinct immature myeloid DC circulating in the blood of cattle have been defined by the expression level of CD205, which can also have a different function in shaping the adaptive response (14). Within bovine tissues, cells stained with anti-CD205 antibody were scarce, and their location was mainly in nonlymphoid tissues (37). It was proposed that CD205Hi moDCs may behave as suppressive moDC subpopulation. Thus, it is important to identify DC lineages in further studies. It is also interesting to note that, while NY93 infected both types of mo, 1a-SINGER strain could only infect adherent CD14+ PBMC. The biological relevance of this difference should be further explored.
Infection with BVDV, even with cpBVDV, did not cause cytopathic effect in moDC. This has also been shown previously for CSFV (7) and BVDV (13,33). Interestingly, the cpBVDV used as inoculum to infect moDC and also the virus progeny produced by the infected moDC were both cytopathic in MDBK cells. The mechanisms that DC use to survive infection with cpBVDV remain to be elucidated (3). Minor variants selection in DC has been described for human hepatitis C virus (HCV); analysis of genome sequences has shown that different cell types can harbor different variants (28). It is also important to note that only infectious, but not inactivated BVDV, could decrease moDC apoptosis at 48 hpi, suggesting that the antiapoptotic effect is probably dependent on intracellular viral replication and perhaps related to the effect of a nonstructural viral protein. Experiments designed for purpose are required to test this idea.
To our knowledge, there is no information on how pestiviruses infect moDC. In many flaviviruses, lectins are responsible of virus entry to DC. Kaimori et al. demonstrated that a pseudotyped vesicular stomatitis virus possessing HCV-E1 and E2 enters immature moDC through the interaction with lectins in a Ca(2+)-independent manner (20). DC-SIGN mediates dengue virus infection of human DC (44). Preincubation of moDC with mannan did not inhibit BVDV infection, neither did EDTA, meaning that BVDV infection is not mediated by the mannose receptor or depends on Ca(2+). In this study, we demonstrated the role of E2 glycoprotein in mediating BVDV infection on moDC. Infection was blocked in a dose-dependent manner by competing the virus with a truncated recombinant E2 protein. Our results show that the 50% inhibitory concentration (IC50) was 0.5 μg/mL for both virus strains, which is in agreement with the IC50 values obtained with a similar experiment on MDBK cells (18). Infection was also prevented when using anti-E2 antibodies, regardless the species. Thus, BVDV infection is enabled by E2, not following an Fc-mediated mechanism. In this study, we provided only preliminary evidence on the possibility of a receptor-mediated entrance of BVDV to moDC. More studies are needed to identify the receptor and assign a role of this specific virus entry in the moDC activity and virus pathogenesis.
Infection of CD11b+ moDC with BVDV or treatment with inactivated virus did not modify the expression of surface molecules such as MHC-I, MHC-II, CD40, and CD86 at short times postinfection (6 hpi). These data are in agreement with those reported by Glew et al. (13), who did not find evidence for a downregulatory effect of BVDV infection on the expression of this range of surface molecules at 48 and 72 hpi. BVDV-infected moDC did not upregulate the expression of MHC-I and II after treatment with Toll-like receptors (TLRs) ligands like LPS and poly I:C, indicating that at least at early time points postinfection, moDC were arrested in an immature state. On the contrary, moDC incubated with inactivated BVDV and then stimulated with poly I:C or LPS recovered the expression of activation markers, meaning that these cells remain capable of being matured by an appropriate stimulation signal. This is the first data on the effect of inactivated BVDV on moDC maturation.
BVDV-infected moDC produced IL-10 at 6 hpi. This cytokine can be responsible for inhibiting DC maturation and functions by reducing the expression of costimulatory molecules (25). Expression of IL-10 by PBMC has been reported for CSFV (43) and HCV infection (38). However, Glew et al. could not confirm expression of this cytokine in infected moDC at 48 hpi (13). It is possible that this suppressive interleukin is upregulated only at short times after infection, favoring the DC impairment observed at 6 hpi.
Interference with DC maturation has been reported for many viruses as a strategy to evade the immune system; that is, Vaccinia, Varicella-zoster virus, human immunodeficiency virus, and HCV (1,2,22,26). Porcine DC infected with CSFV (7) or bovine moDC-infected BVDV (13) did not upregulate costimulatory molecules, but retained their capacity to function as APCs after 24–48 hpi. Previous studies have shown that infection with BVDV transiently suppresses a T-cell memory response to a third-party antigen in vivo (8). In this study, we show that the inactive state of the BVDV-infected moDC was evident during the first day after infection, corresponding with the peak of viral replication. However, at 48 hpi, infected moDC could undertake their APC activity. Our data suggest that the capacity of BVDV to infect and replicate in moDC might lead to their arrest in an immature state during the first hpi, while their activity can be recovered once the virus reduces its replication rate.
Taken together, our results show that BVDV (different strains and biotypes) can rapidly infect moDC with high efficiency and replicate in these powerful APC. The virus has probably developed a mechanism for preventing antiviral responses by inducing suboptimal maturation rather than strong virus-specific impairment, which was antiapoptotic, together with a specific entry E2-mediated mechanism. The impairment of moDC function is coincident with the peak viral replication and is reverted once the replication rate diminishes, suggesting that probably one or several nonstructural proteins produced only during viral replication may be responsible of this effect. Experiments designed for purpose are needed to explain the mechanisms underlying these observations.
Overall, this study agrees with a model where DCs can act as an amplification system for BVDV, similar to the one described recently for HCV (35) in which infected APC are not killed and continuously produce new viral particles, allowing the spread of the virus to other cell types. The impairment of the DC function as APC along the first day after infection may facilitate a rapid dissemination of high amount of infectious BVDV particles, contributing to the virus-induced immunosuppression and delaying the onset of the acquired immunity against the virus.
Footnotes
Acknowledgments
Financial support was received from the National Agency for the Promotion of Science and Technology of Argentina (ANPCyT) through the PID 011-2008 awarded to A.V.C. N.C. and A.V.C. are researchers of the National Research Council of Argentina (CONICET). O.L.F., M.E.Q., and F.C.M. were supported by doctoral fellowships from CONICET. W.C. was a postdoctoral fellow of ANPCyT. We would like to thank Mrs. Mariela Gammella for her assistance with the flow cytometry analysis. We are also grateful to Dr. Danilo Bucafusco and Dr. Mariano Pérez-Filgueira for their invaluable help with this article.
Author Disclosure Statement
No competing financial interests exist.