
Abstract
A picornavirus (Ljungan virus) has been associated with diabetes in its wild rodent reservoir and in diabetes-prone biobreeding (DP-BB) rats. We attempted to alter the development of diabetes in DP-BB rats using two anti-picornavirus compounds (pleconaril and APO-N039), singly or in combination. Antiviral therapy was initiated 2 weeks before expected onset of diabetes. Pleconaril or APO-N039 alone did not affect the debut of diabetes. However, animals receiving a combination of both compounds were protected for at least the entire period of treatment (4 weeks after expected time of diabetes onset). Immunohistochemistry demonstrated that the presence and distribution of virus antigen in the pancreatic islets coincided with the clinical status of the animal. Data indicate that a treatable picornavirus can be involved in the cellular assault resulting in diabetes and in these cases the disease mechanism appears to involve a virus present in the pancreatic beta cell mass itself.
Introduction
T
We have earlier reported the finding of a picornavirus named Ljungan virus (LV) present in Scandinavian vole and lemming populations and the association of this virus with diabetes in these animals (3,6,16,18). The role of LV as a diabetogenic picornavirus gained further merit when it was found that laboratory-infected mice developed glucose intolerance when they were subjected to stress (19). The severity of diabetes in LV-infected mice could be reduced using specific LV antisera in combination with the picornavirus-specific antiviral compound pleconaril (8). The finding of LV antigen was subsequently reported in beta cells of diabetic BB rats and it was later shown that the onset of diabetes could be delayed in BB rats receiving a combination of two antiviral compounds (pleconaril and ribavirin) before onset (8,17).
It should be noted, however, that we still lack conclusive evidence of the role of LV as the etiologic agent in diabetes disease in its wild rodent reservoir, in animal models such as the BB rat as well as in humans. Evidence such as the fulfillment of Koch's postulate in animal would require a virus-free colony, which has not yet been established. Nor has any virus isolate, molecular confirmation or characterization confirmed the LV diabetes hypothesis.
Still, we believe that the hypothesis has key components favoring the idea that an LV-related virus can cause diabetes in wild rodents, BB rats, and humans. Using antiviral compounds alone, or in combination is one line of evidence that may help us determine the role of virus and also providing evidence of where in the virus world this etiologic agent is to be found.
As part of the effort to gain indirect evidence for a viral involvement in diabetes pathogenesis in general, and in BB rats in particular, we have continued to screen antiviral compounds. The previous report using a combination of pleconaril and ribavirin in BB rats, found a delay of diabetes onset for 1 week still significant. In the present study two antiviral compounds, one of them pleconaril previously used and an anti-picornavirus kinase inhibitor, were used in the BB rat model. In this study, we report the finding that this new combination of antiviral drugs could delay the onset of diabetes for the entire period of treatment and in some cases this effect was maintained for an indefinite period following cessation of treatment.
Materials and Methods
Antivirals compounds
Two antiviral compounds were used in the experiments: First, pleconaril, 3-{3, 5-dimethyl-4-[3-(3-methyl-isoxazole-5-yl)-propoxy]-phenyl}(11)-5-trifluoromethyl-(1,2,4) oxazole, is a direct-acting broad-spectrum anti-picornaviral agent originally developed for the treatment of the common cold (5,20). Pleconaril binds to the hydrophobic pocket of the virus and creates conformational changes, which increases the rigidity of the virion and decreases its ability to interact with the cell receptor.
Second, APO-N039, (3,4-dimethoxyphenyl)-N-[(4-fluorophenyl)methyl]-2,5-dimethyl-pyrazolo[1,5-a]pyrimidin-7-amine is a host-targeting antiviral acting on PI4KIIIβ. An appropriate combination of viral and cellular proteins is necessary for the efficient production of viral RNA, proteins, and particles. Particular viral proteins promoting the specific lipid microenvironment are essential for enterovirus replication. PI4KIIIβ inhibition can interfere with this process and, therefore, have an antiviral effect. APO-N039 demonstrates antiviral influence in tissue culture to a broad spectrum of various picornaviruses. Inhibition of PI4 kinases and PI3 kinases were performed by Reaction Biology Corp. (Reaction Biology Corp., Malvern, PA) and studied by using the ADP-Glo™ Kinase Assay and the homogeneous time-resolved fluorescence assay, respectively.
The antiviral effects of pleconaril and APO-N039 alone or in combination to virus replication were determined in a 96-well cell culture assay that measured the protection by the drugs on infected cells from cytopathic effect (CPE) (20). For each drug combination, two substances were diluted separately by serial dilution before being mixed in a well containing green monkey kidney cells to create a two-dimensional matrix of single and combined drugs. LV 145SL was added to wells containing substances and plates were incubated at 37°C, 5% CO2 until CPE. Viable cells were visualized by staining with Crystal Violet solution. The neutralization was measured using an enzyme-linked immunosorbent assay microplate photometer (540 nm) and calculated by comparing the absorbance for a sample well with uninfected control cells.
Pleconaril and APO-N039 were administered to the animals in a dose of 100 mg/kg body weight once daily using a 10 mg/mL formulation containing the vehicle 0.4% (v/v) Tween 80 + 2% (v/v) glycerol +15% (w/v) Hydroxypropyl-beta-cyclodextrin.
Experimental animals
The DP-BB rats were obtained from the breeding colony maintained at the Karolinska Institute (14). The studies performed in BB rats were approved by the Animal Experiment Ethics Committees at Karolinska Institutet (N445/12). The animals were housed under specific pathogen-free conditions in a temperature- and humidity-controlled room with 12-h light–12-h dark cycles. They were fed R36 diet and water ad libitum. Animals of both sexes were used. The overall incidence of diabetes among the DP rats used in the colony study is 100% with 60 days as the mean age of onset.
Two untreated DP-BB rats were sacrificed on day 33 and 74, respectively. Pancreases were collected and analyzed by pathology and immunohistochemistry (IHC) as a reference to animals in treatment groups
Four experimental treatment groups consisted of six rats given pleconaril only, six given APO-N039 only, seven given a combination of both compounds, and nine given the vehicle solution serving as a control. In a secondary experiment, five of the nine control animals received pleconaril–APO-N039 combination treatment after onset of diabetes to investigate the effect at this time.
The two groups receiving only single drugs and control group animals began treatment at 48 days of age. For the group receiving both compounds, four animals started medication at 48 and three at 53 days of age, respectively.
Therapy in all animals was terminated when an animal reached the age of 90 days in the three treated groups.
All animals were followed until the onset of type-1diabetes (blood glucose 15 mmol/L or above and/or ketones). Following onset of diabetes, animals received subcutaneous insulin injections (Prozink, Boehringer Ingelheim 15 U/kg body weight daily). Antiviral compounds and the control vehicle were administrated using an oral gavage once daily.
Statistics
The basic data (time to onset of diabetes under various experimental treatments) superficially fall within the purview of time-to-event survival analysis, but because the combination drug antiviral treatment resulted in no instances of diabetes onset, the conventional survival analysis assumptions are not met (the consistently censored observations permit no useful observations of that treatment group). Analysis of variance was used to compare the days until diabetes onset across treatments while handling the combination drug group as existing without diabetes up to the time of treatment ending. For the small numbers involved in comparing the effective treatment group with differing drug initiation days, the nonparametric Kruskal–Wallis test was used to compare the outcomes, following significant analysis of variance.
Immunohistochemistry
Formalin-fixed paraffin-embedded pancreatic tissues were analyzed by IHC. The tissues were fixed in 4% formalin and embedded in paraffin as described previously with minor modifications (22,23). The presence of LV-specific antigen was visualized using a polyclonal recombinant LV VP1 raised in rabbits (25). As control we used serum from a rabbit immunized using the same protocol, but with the carrier GST protein only. Tissues from LV-infected and noninfected animals were included as controls. The specificity of the rabbit antibodies was verified by analyzing control specimens generated by mixing infected tissue culture cells with noninfected cells followed by formalin fixation and paraffin embedding. The specificity of the reaction was also confirmed by blocking the signal with LV antigen in parallel with control antigen.
Results
Effects of antiviral compounds in vitro
Pleconaril has previously been characterized as a capsid binder in that it binds to a hydrophobic pocket in the VP1 protein of the picornavirus (12). The compound APO-N039 showed activity on the cellular target PI4KIIIβ and inhibited selectively the kinase with an IC50 of 4.7 nM (PI4KIIIα = 0.8 nM, PI3Kβ >10 nM, PI3Kα >10 nM). Pleconaril showed antiviral activity (>50% inhibition of CPE) to LV 145SL at the concentration of 10 μM or higher. For APO-N039, the same antiviral activity was detected at the concentration of 5 μM or higher. APO-N039 remained active (>50% neutralization) at a 10-fold lower concentration when pleconaril was present at the subactive concentration, suggesting an additive effect between the two compounds.
Effects of antiviral compounds in BB rats
BB rats assigned to one of four drug treatment groups were followed to record time-to-onset of diabetes.
All seven BB rats receiving combination treatment (pleconaril–APO-N039) maintained normal blood glucose levels for the entire period of treatment (90 days of age) that was 1 month after expected onset of diabetes (Fig. 1). Details of each rat treated with combination are given below and in Table 1. Rats receiving APO-N039 alone developed diabetes on mean day 62 (55–69). Antiviral treatment continued for 23 days (7–35) post onset of diabetes without any effect on the diabetes during therapy. The group receiving pleconaril alone developed diabetes on day 62 (54–75). Antiviral treatment continued for 21 days (6–33) post onset of diabetes without any effect on the diabetes during therapy. Control animals (n = 9) developed diabetes on mean day 61 (49–69). After serving as controls for time of disease onset, when receiving sham therapy, the same animals received combination therapy (pleconaril–APO-N039) the same day diabetes was diagnosed and insulin treatment initiated. Five of the nine control rats were given combination therapy after onset of diabetes and therapy was maintained for mean 18 days (7–41 days) without any effect on the diabetes in these “postdiabetes-onset”-treated animals.
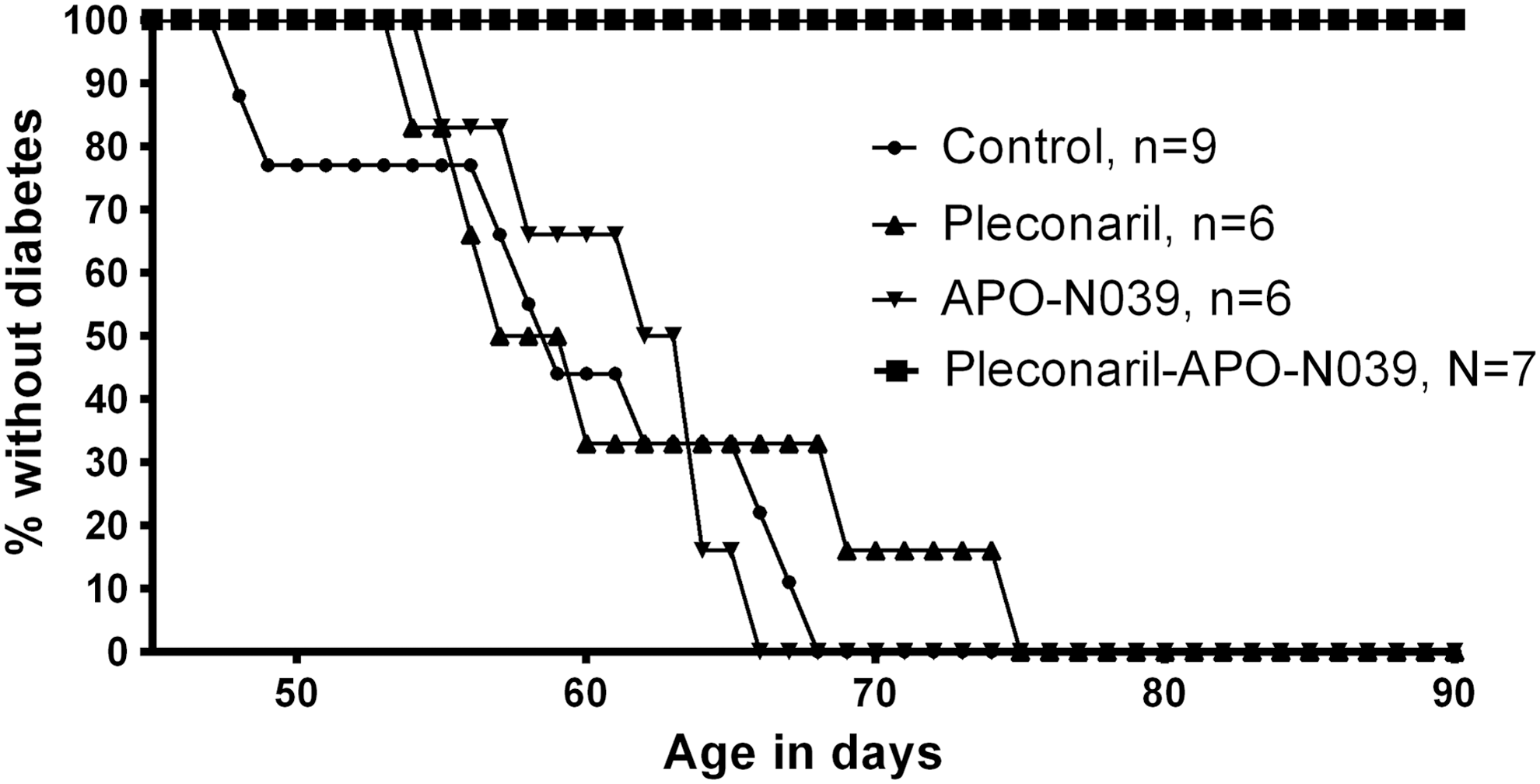
Time to diabetes onset in BB rats under four experimental treatments: Control, Pleconaril, APO-N039, Combination Pleconaril–APO-N039. None of the animals treated with the combination therapy through 90 days became diabetic, while all animals in the other three experimental treatments became diabetic on or before 74 days. BB, biobreeding.
BB, biobreeding; BG, blood glucose; GTT, glucose tolerance test.
The variation in duration of therapy post onset of diabetes was because most animals experienced signs and symptoms of disease and the animal ethical protocols required that these animals be euthanized.
Difference in the time-to-diabetes among the four treatments was highly significant when tested by analysis of variance (F = 25.98, df = 1,4, p < 0.0001), indicating that the time to diabetes of the four groups could not have reasonably come from a single group by chance alone. Post hoc comparisons showed that, the control treatment group and the two single-drug treatment groups do not differ, while each of these are quite distinct from the group receiving the combination therapy, in which no animals became diabetic during treatment (Fig. 1). The statistical result was conservative in that the two-drug treatment group animals were artificially assigned diabetes onset on day 90 for the statistical test, while in fact animals in this group became diabetic only after day 93, if ever.
To examine sensitivity to the timing of treatment during development, the seven animals in the two-drug group were started on treatment at two times: Four animals at 48 days and three animals at 53 days of age. The outcome for each of the seven BB rats receiving the combination of antiviral compounds after terminating therapy at 90 days of age is summarized in Table 1 and described below.
• Rat 1 starting therapy at day 48 and ending antiviral therapy at day 90 never developed diabetes. Normal blood glucose (BG) levels were recorded during the entire follow-up period and a normal glucose tolerance test (GTT) was recorded before the animal was euthanized at 130 days of age.
• Rat 2 developed diabetes 6 days after stopping antiviral treatment on day 90. A second period of treatment was immediately initiated resulting in normal BG after 10 days of treatment. Antiviral treatment continued for 10 days, after which normal BG was restored. This rat maintained normal glucose levels for a total of 24 days during this second period, of which 14 days were after the second period of antiviral treatment was terminated. After this second period of normal blood glucose levels, the animal again became diabetic and was euthanized.
• Rat 3 developed diabetes 18 days after terminating therapy on day 90 and was put on a second period of antiviral treatment 4 days later. This individual did not respond to the second period of treatment, remained diabetic until being euthanized after 13 days.
• Rat 4 developed diabetes 16 days after terminating antiviral treatment and was euthanized at this time.
• Rat 5 developed diabetes 4 days after the ending of the first treatment period on day 90. Second period of therapy started 5 days later and lasted for 2 weeks. Normal BG was measured after 6 days of treatment. This individual maintained normal BG after terminating on treatment day 113 for the entire follow-up period. Normal GTT was recorded at day 200 when euthanized.
• Rats 6 and 7 developed diabetes 3 and 5 days after ending the first period of treatment, respectively. Both animals were euthanized at this time.
Focusing on the seven two-drug-treated rats, we note that one animal (rat 1) never developed diabetes following cessation of treatment, despite 40 days of observation before being euthanized. Another individual (rat 5) developed diabetes during 1 week after the 37-day-long period of treatment. After an additional 2-week-long period of treatment this animal maintained normal blood glucose levels for 3 additional months before being euthanized at 200 days of age.
Of the remaining individuals, three had drug therapy initiated at 48 days of age and three at 53 days. An explicit test of the influence of 5 days difference in therapy initiation on diabetes onset showed a suggestive difference between groups (Kruskal–Wallis p = 0.0495). Thus, despite the small sample size, a period influencing viral pathogenesis may be reflected in later onset of diabetes following earlier antiviral treatment.
Pathology and IHC
One DP BB rat was sacrificed on day 33, ∼1 month before expected disease onset as reference shows normal histology. The cells within the islets are normal with prominent central nuclei of same shape and size and with clear cytoplasmic areas. The islets have no fibrosis, no infiltration with inflammatory cells, and have normal surrounding pancreatic acinar tissue. IHC staining of the same sections shows that all or almost all cells are positive in the periphery of the islet, whereas the central part of the islet is free of detectable viral antigen (Fig. 2A). Rodent islets (including in rats) are composed of beta cells located in the core of an islet, which is surrounded by a mantle of alpha and delta cells, indicating that the insulin-producing beta cells in this animal are at this time free of detectable viral antigen (2). The animal sacrificed 2 weeks postdiabetes onset (74 days of age) shows a significant loss of islet mass. Approximately 10–20% of the number of islets remains and the size of the remaining islets have decreased by ∼80% as compared with the BB rat scarified 1 month before diabetes onset. The remaining islets have irregular boundaries and the rounded/spherical normal architecture does not exist anymore. The cells within these irregular islets are shrunken with wider surrounding extracellular space and signs of both vascular congestion and fibrotic changes. Nuclei are variable in shape and size, and some of the cells have pyknotic nuclei. All cells within the islets are now staining for viral antigen detected by IHC.

Illustrative examples of islets of Langerhans of BB rats stained with H&E (left series) and immunohistochemistry using antisera against LV protein 1 (right series).
BB rats receiving single treatment with pleconaril or APO-N039 islet mass had decreased with significantly fewer islets and with those that remain being of smaller size. The diminished mass of islets associates with decrease in the number of cells and disorganized surrounding pancreatic acinar tissues. The cells have reduced cytoplasmic area (shrunken cells) and the embracing extracellular matrix is expanded with sign of fibrosis. All islets, including the few found with normal size and shape, stained positive for virus antigen in both the central part and the periphery exemplified in Figure 2B and C.
Figure 2D is an animal (rat 6) receiving the two-drug combination. This individual showed a reduced number of islets in total as compared with prediabetic BB rats. Many islets were of normal size and some smaller than normal. All of the islets are surrounded by well-defined acini and at the level of cells; they have normal cytoplasm and rounded nuclei. No inflammatory cell infiltration, no fibrotic changes, and no viral antigen staining were detected in any islet visualized in the pancreas of this animal. Figure 2E (rat 7) is another individual revealing a picture similar to rat 6, with interesting contrasts suggesting a range of response among identically treated animals. It also received the combination treatment, but in this case islets stained positive in the surrounding mantle with the alpha and delta cells.
Discussion
In the present study, we have found that when DP BB rats are treated with pleconaril–APO-N039 in combination, this can affect the onset of diabetes during the period of treatment, demonstrating a means to alter disease outcome. When treatment was withheld, diabetes appeared in 6/7 treated animals, whereas in one, animal diabetes never occurred during the period of observation following cessation of treatment. Another animal developed diabetes shortly after the first period of treatment and received a 2-week-long additional antiviral combination period resulting in a diabetes-free period, terminated after 3 months when this animal was euthanized still diabetes free. These examples at the very least suggest the possibility of viral elimination and cure.
We show that employment of a two-drug cocktail was essential for the favorable outcome, and apparently acting through distinct aspects of the viral life cycle. We also present data that viral pathogenesis and its control through antiviral agents may be dependent on the timing of these treatments, showing that BB rats initiating treatment only a few days later (48 vs. 53 days) experience diabetes earlier.
Prior work on pancreatic islet morphology in the BB rats during diabetic pathogenesis makes it possible to follow cellular changes during diabetogenesis (2,13). Animals treated with only one antiviral compound (Fig. 2B, C) that each became diabetic over the course of the experiment retained visible islets with LV staining of the entire islet, including the insulin-producing beta cells, suggesting that one antiviral may protect the morphology, but not the functionality of the beta cell. Animals given the two-drug cocktail remained glucose tolerant during treatment and sometimes beyond.
The pancreatic micrographs from animals that became diabetic after treatment had ended included examples of both IHC-negative and -positive islets. Similar to observations on untreated BB rats (Fig. 2A) examined 1 month before onset of diabetes, the positive staining was only seen in the periphery of the islets (Fig. 2D, E).
When we initially reported the finding of LV antigen in the islet of BB rats, we used both LV-specific monoclonal antibody and polyclonal antisera raised against recombinant LV VP1 protein (11,17,25). More information is presently available regarding sensitivity and specificity using the polyclonal antisera in IHC and it was therefore used in the present study.
The BB rat is perceived as an animal model in which autoimmunity is the main driving force in the pathogenesis (13). However, the observation of LV antigen staining pattern in the beta cells and the diabetes signs and symptoms both responding to antiviral treatment suggest that a picornavirus infection can be involved in the development of diabetes. In addition, the short time frame between start of antivirals and clinical effect in individuals receiving new bouts of therapy favor this interpretation. Assuming the scenario described above is true, the immune system action of removing the insulin-producing beta cells would be an appropriate response to an infected cell.
Currently, a limitation in our investigation is that the virus infection is diagnosed by IHC only. We previously reported positive results testing tissue from BB rats using polymerase chain reaction (PCR) (17). Unfortunately, despite major efforts, we have not been able to confirm these results. No confirmatory diagnostic assay such as virus isolation or PCR followed by sequence analysis confirming the presence and characteristics of the virus is presently available. This situation mimics in many ways the current debate on type 1 diabetes in humans, in which reports suggest the involvement of picornavirus without standard confirmatory evidence (1,10,21).
The enterovirus was first associated with diabetes in what turned out to be rare, but genuine instances of viral causation several decades ago (7). Observations since then of infections in a range of tissues and organs, including that of the pancreas, have made clear the catholic range of attractions of many picornaviruses (24). Many studies have now examined enteroviruses in cases of type 1 diabetes individuals. While a meta-analysis of this work demonstrated a significant excess of enterovirus infection associated with autoimmunity and type 1 diabetes compared with controls (27), the proportion of type 1 diabetes cases explained was modest, suggesting that a single picornavirus species may not be the typical culprit. Transcriptome signatures of interferon in patients before type 1 diabetes onset identify metabolic signals suggesting a viral infection of the pancreas (4,11). In this study, we offer evidence through treatment of type 1 diabetes-like-disease in a rodent model with picornavirus-specific antivirals pointing to ongoing pathogenesis due to one or more members of the Picornaviridae family.
The historically accepted means of linking an infectious agent to a particular disease has been well defined. Isolation and characterization of the etiologic agent followed fulfilling Koch's postulates. Epidemiological and serological studies may offer valuable supporting evidence associating a suspected etiological agent with a disease. Establishing a suitable animal model and developing prophylactic treatment or specific therapy usually follows.
This study serves as an example as to how, with the identification of safe and potentially effective antivirals, clues to viral etiology in humans may also be gained. Therapy in humans could be initiated when diabetes is first diagnosed. Extrapolating from the present murine experiments, antiviral protective effects appear rapidly enough for pancreatic beta cell rescue. In addition, tools are available to identify patients with imminent diabetic onset (e.g., predisposing human leukocyte antigen haplotypes, glutamic acid decarboxylase-, insulin- or islet antibodies) (26).
In conclusion, this present report advances evidence that a picornavirus related to LV may be involved in the onset of diabetes in the BB rat and that the disease mechanism includes a viral attack on the pancreatic beta cell mass itself that may be curtailed through viral treatment.
Footnotes
Author Disclosure Statement
No competing financial interests exist.