
Abstract
HIV infection is characterized by a high degree of immune activation. It has an impact on CD4 cell count and populations' distribution and function. T regulatory cells (Tregs) were found to play a controversial role in the course of infection because of their beneficial effect on the degree of immune activation and unfavorable influence on the antigen-specific responses. The goal of the present work was to study the relationship among interferon-γ (IFN-γ), spontaneous lymphocyte proliferation, and regulatory T cells in HIV patients receiving therapy. Three lymphocyte populations, isolated after a stepwise magnetic separation from 17 individuals, were investigated—peripheral blood lymphocytes, CD4+CD45RA− (CD4+TM), and CD4+CD45RA−CD25− (TMCD25depl.) cells. The spontaneous, phytohemagglutinin (PHA) and HIV-1p24Ag-stimulated IFN-γ production and the spontaneous lymphocyte proliferation were evaluated. The potential of Tregs to establish a productive infection was determined by measurement of free HIV-1p24 antigen. Two types of constellations among subsets were found. In the first one (in 11 subjects), the spontaneous INF-γ inversely correlated with the spontaneous proliferation in all fractions (r = −0.9, p < 0.001). Conversely, in the second group (six subjects), no associations between the selected parameters were observed. The overall increase in p24-stimulated IFN-γ from TMCD25depl. cells was weak. Four samples: one in Tregs and three in TMCD25depl. cells were positive for the free p24 antigen. No association with the CD4+ T cell count, percentage of Tregs, and stage of infection was determined. In conclusion, our results demonstrate that IFN-γ could impact the proliferative capacity of non-Treg cells by fuelling Tregs. Furthermore, Tregs may control the spontaneous lymphocyte proliferation, but are less powerful in the suppression of Ag-specific IFN-γ production from non-Treg lymphocytes. The direct viral influence on Treg functions should be also considered.
Introduction
S
The classical and actual knowledge on the role of Tregs in HIV infection suggests that it is complex and ambiguous. The situation becomes more intriguing when considering that Tregs are strongly dependent on IL-2 production and are a subject of invasion for HIV. As a result, it could be expected that (i) the cytokine switch, observed early in the course of infection, could have an impact on the function of Tregs and (ii) the virus per se might affect the Treg machinery in a similar manner to its action in conventional T cells, mainly perturbing the IL-2 signaling pathway (23).
Interferon-γ (IFN-γ) has attracted scientific attention for many years, because of its crucial role in immune response and cytokine profile determination. It exerts its pleiotropic biological effects through a specific receptor at the target cell surface (15,16). IFN-γ has been considered to play a major role in the actual development of immune response. Its effects on Tregs have been studied in autoimmune disorders, tumors, and transplants (34,45). The latest research insights in autoimmunity have demonstrated that besides its pleiotropic activity, IFN-γ exerts additional functions and, thus, could significantly contribute to the immune homeostasis. Moreover, IFN-γ is linked to the exacerbation of clinical symptoms and pathologic events in several autoimmune and proliferative diseases and it is also important for successful transplantation. Interestingly, it was found that the bridge between IFN-γ and autoimmune deteriorations is CD4+CD25+ regulatory T cells (7,10,12).
In the present study, we analyzed the relationship between regulatory T cells and IFN-γ production by non-Treg cells in HIV-1-infected patients receiving ART. To find out whether Treg cells have an impact on the capacity of lymphocytes to proliferate and their potential to produce IFN-γ, three consecutively separated lymphocyte fractions from each sample were investigated. The differences, found between the proliferative activity and IFN-γ production among fractions, indicated that IFN-γ might alter the suppressive capacity of Treg cells.
Materials and Methods
Study subjects: 17 HIV-1-infected individuals (median age 37 years, range between 25 and 55), with median duration of HIV-1 infection of 7.5 years (range between 2 and 13 years), were included in the study. All patients had been receiving highly active ART for more than 1 year (median—6 years), and all of them were compliant to therapy. At the time of investigation, the CD4+ T cell count was 0.144–1.375 × 109/L; median −0.700 × 109/L and viral load were at an undetectable level. For flow cytometry and gene expression assessments, an age- and sex-matched group of 15 healthy subjects was used as a control.
Patients and controls were selected to be free of signs or symptoms of autoimmune disorders and as concerning women, also free from Rh signs of sensitization. No selection criteria based on race, sex, or age have been applied. The study was approved by the local ethics committee (protocol #07-07-2005).
Antibodies and reagents
Magnetic separation of regulatory T cells was performed with a Dynal® CD4+CD25+Tregs Kit and a magnetic particle concentrator (MPC-L) from Dynal Biotech. The following fluorochrome-labeled monoclonal antibodies to cellular antigens were used for flow cytometry: CD45 (clone 2D1-FITC) and CD14 (clone MφP9-PE), purchased from Becton Dickinson and CD4 (clone OKT4-FITC), CD25 (clone BC96-PE-Cy7), and anti FoxP3 (clone PCH101-PE) from eBioscience. Cell culture medium, supplements, and stimulating factors were purchased from BioWhittaker, Sigma Aldrich and eBioscience; 3H-thymidine from Amersham. IFN-γ was determined with the commercially available kit from Cellestis. Free HIV-1 p24 antigen (p24 Ag) was measured using XpressBio ELISA Kit. Real-time PCR was performed with reagents from AJ Roboscreen GmbH and Applied Biosystems.
Sample preparation and isolation of CD4+CD25+ Treg cells were performed as previously described (42). Briefly, peripheral blood mononuclear cells (PBMCs), isolated by Histopaque density gradient centrifugation, underwent separation according to manufacturer's instructions, and four lymphocyte fractions were obtained and used in further experiments. Cell fraction purity was determined by flow cytometry and exceeded 95%.
Lymphoproliferative assay (LPA) for measurement of the spontaneous lymphocyte proliferation (SLP-1h) was performed using freshly isolated PBMCs and separated CD4+CD45RA− T cells and TMCD25depl. T cells as described elsewhere (5,42). Briefly, 1 × 105 cells/well were labelled immediately after isolation with 37 Bq 3H-thymidine and cultured in triplicate in a lymphocyte medium (RPMI 1640), supplemented with 10% FCS, 2 mM
Flow cytometry
The following gating strategy was applied for three-color flow cytometry: first total lymphocyte population was identified using markers anti-CD45-FITC/CD14-PE and then CD4+ activated lymphocytes were identified using the markers CD4 FITC/CD25 PerCP (CD4+CD25+). Appropriate isotype controls were included to discriminate between nonspecific and specific staining. Cells were acquired on a FACScan flow cytometer (Becton Dickinson), and the analysis was performed using CellQuest software. Isolated PBMCs were stained for surface antigens (CD4, CD25) using the following staining protocol: cells were washed with cold CellWASH solution (Becton Dickinson) then fixed and permeabilized. Fc receptors were blocked with normal rat serum before labelling with FoxP3 PE (eBioscience) or isotype control. Tregs were determined as CD4+CD25highFoxP3+ cells. They were presented as a percentage of all lymphocytes, as%CD4 × Tregs/100. The absolute count of Tregs was calculated as % Tregs × absolute lymphocyte count/100.
Cytokine measurements
IFN-γ was assessed using ELISA, following the manufacturer's instructions, and results are presented in pg/mL. Cytokine levels were assayed in cell culture media of mononuclear cells (1 × 105 cells/well), nonstimulated and stimulated with phytohemagglutinin (PHA; 5 μg/mL) and HIV-1 p24 antigen (0.25 μg/mL) for 20 h.
Measurement of HIV-1 p24 antigen
Free p24-antigen was determined in cell culture media from the above cell fractions, being stimulated with rhIL-2 at 25 μg/mL for 96 h, using ELISA (XpressBio), according to manufacturer's instructions. Results were calculated from a standard curve and presented as concentration in pg/mL, according to manufacturer's instructions.
RNA extraction and reverse transcription
Total RNA was extracted from 1 × 106 cell population for each cell fraction, using Invisorb Spin Blood RNA Mini Kit (STRATEC), according to manufacturer's instructions. Ten microliters total RNA was transcribed into cDNA, using High Capacity cDNA Archive Kit, in a final volume of 20 μL with random primers, following manufacturer's instructions.
Relative gene expression analysis by real-time PCR
FoxP3 and HPRT (endogenous control) gene expression was performed in duplicates in 50 μL reaction on ABI 7500 Real-time PCR instrument (Applied Biosystems, Inc., Foster City, CA), as previously described (42). Relative gene expression data were acquired by SDS software and analyzed with RQ module.
Statistical analyses
The mean values of triplicates for LPA and of duplicate FoxP3 gene expressions were further included in statistical analyses. First, to determine the appropriate test, data were assessed for normality using the Shapiro–Wilk test. Independent sample or related sample Mann–Whitney tests were performed when p-values were <0.05 from the Shapiro–Wilk test. While p > 0.05, independent sample two-tailed t-test was applied in comparison upon checking the equality of variances, according to the test of Levene. Wilcoxon signed rank test or paired sample t-test was used to compare variables within a single group. The significance of differences was considered when p ≤ 0.05. Upon bivariate correlations, significant associations were defined as p < 0.05 and Pearson's correlation coefficient r (when correlated groups were expected to have a Gaussian type of distribution) or Spearman's p > 0.5. Statistical analyses were performed using SPSS software.
Results
Analysis of regulatory T cells
All individuals were tested to evaluate the percentage of regulatory population in the peripheral blood. Regulatory T cells were determined as CD4+CD25high+FoxP3+ T lymphocytes thus resembling the total Treg pool in the periphery (Supplementary Figs. S1 and S2; Supplementary Data are available online at
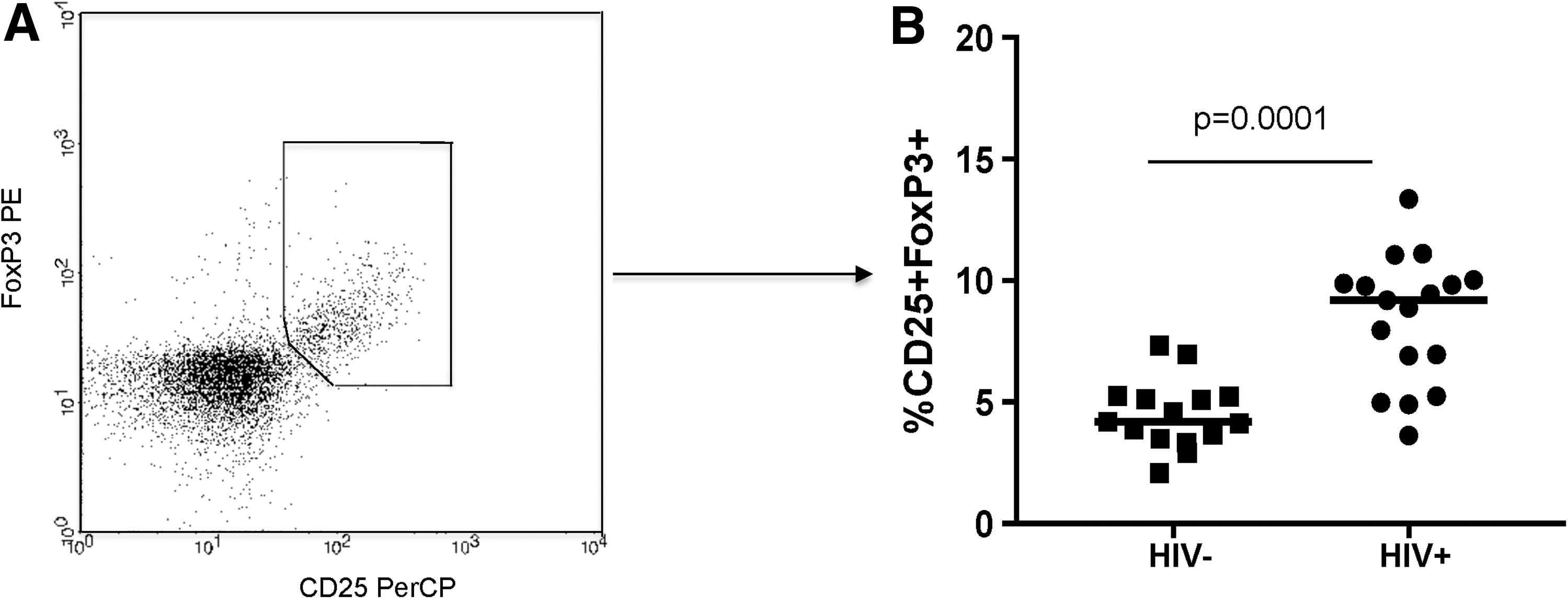
T regulatory cells.
Following magnetic separation steps, three lymphocyte subsets were obtained: CD4+CD45RA− (conditionally termed T-memory—TM) cells, containing both Tregs and non-Tregs; CD4+CD45RA−CD25− (TMCD25depl.) cells; and CD4+CD45RA−CD25+ (Tregs). The analysis of FoxP3 gene expression showed only traces of FoxP3 in the CD25-depleted fraction in the patient group. In all cases, the highest levels of the FoxP3 expression were found in the fraction of CD25+ cells (data not shown).
Tregs influence IFN-γ production from conventional T cells
To investigate the capacity of Tregs to modify IFN-γ production, for every subject sample separately, we studied three lymphocyte fractions: (i) PBMCs, (ii) TM, and (iii) TMCD25depl. cells. Cells were cultured as either nonstimulated, to measure the spontaneous IFN-γ (spIFN-γ) production, or stimulated with HIV-1 p24 antigen (IFN-γ(p24)) and PHA (IFN-γ(PHA)). In two patients with low cell numbers, IFN-γ evaluation was not performed. Tregs per se have not been examined in IFN-γ assays, since they exhibit only an encouraged and transient IFN-γ production (38).
All fractions were found to spontaneously produce IFN-γ, but to a different extent. Not surprisingly, very low levels of free spIFN-γ were found in nonstimulated PBMCs and TM cells in all patient samples: median (range): 1.60 (0.00–29.40) and 2.60 (0.00–28.75) pg/mL, respectively. On the contrary, in supernatants from TMCD25depl. cells, the highest concentrations were determined as: median (range): 5.8 (0.2–46.8) pg/mL, p < 0.05 (for both PBMCs and TM) (Fig. 2A). The overall analysis of the PHA stimulated IFN-γ production in patients demonstrated quite a heterogeneous response in PBMCs and TMCD25depl. cells, while in all
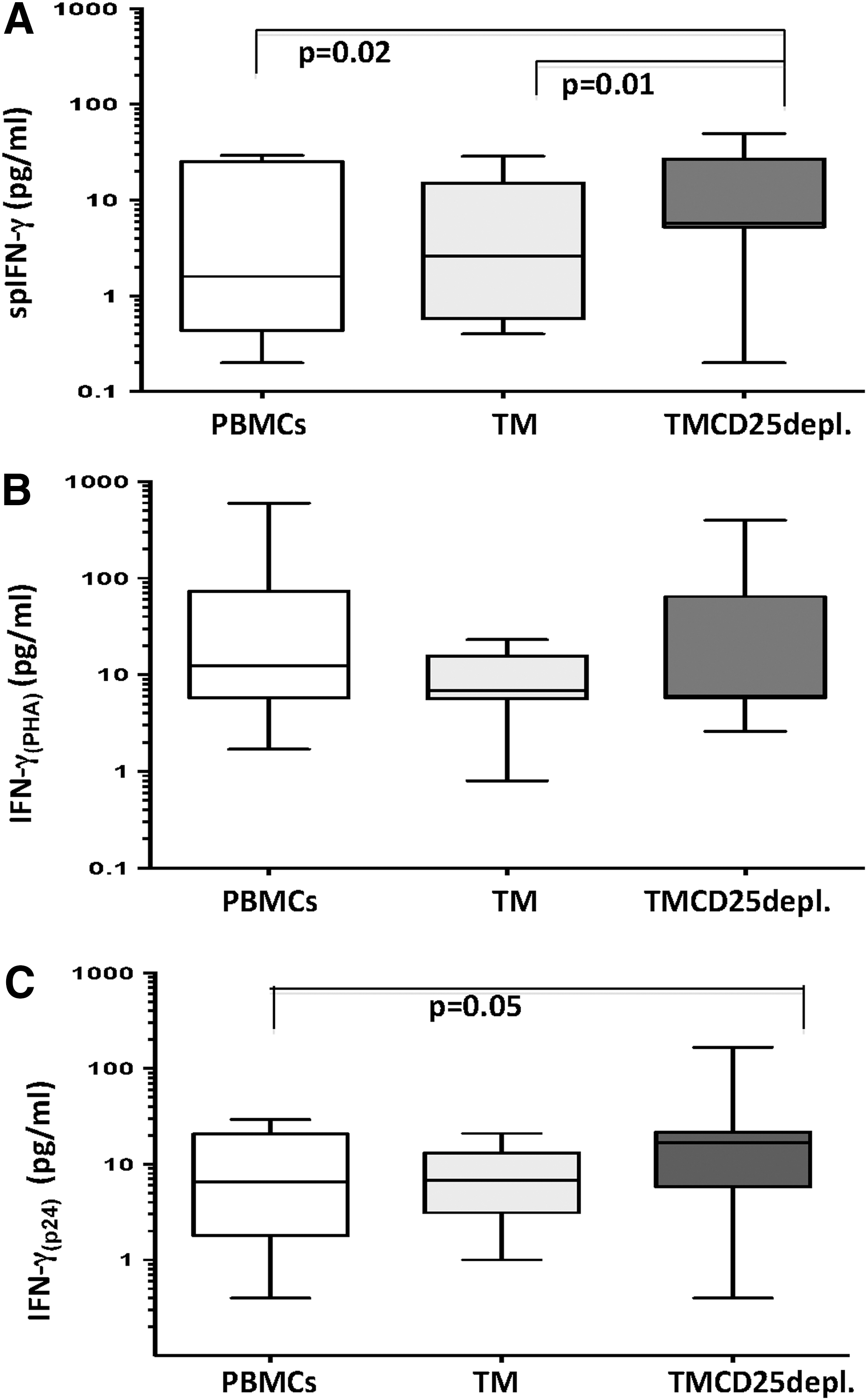
Spontaneous and stimulated IFN-γ production. In vertical order are shown differences of IFN-γ production between nonstimulated
The inside fraction analysis showed an increase of IFN-γ(p24), in comparison to a nonstimulated one, indicating the cells' capacity to respond to the specific antigenic stimulus. Collectively, the amount of free IFN-γ(p24) was found to be lower than the one in the positive control IFN-γ(PHA). The increased IFN-γ(p24) from TMCD25depl. cells versus PBMCs, but not TM, suggested none or slight influence of Tregs on the Ag-specific IFN-γ response by TM (Fig. 3).
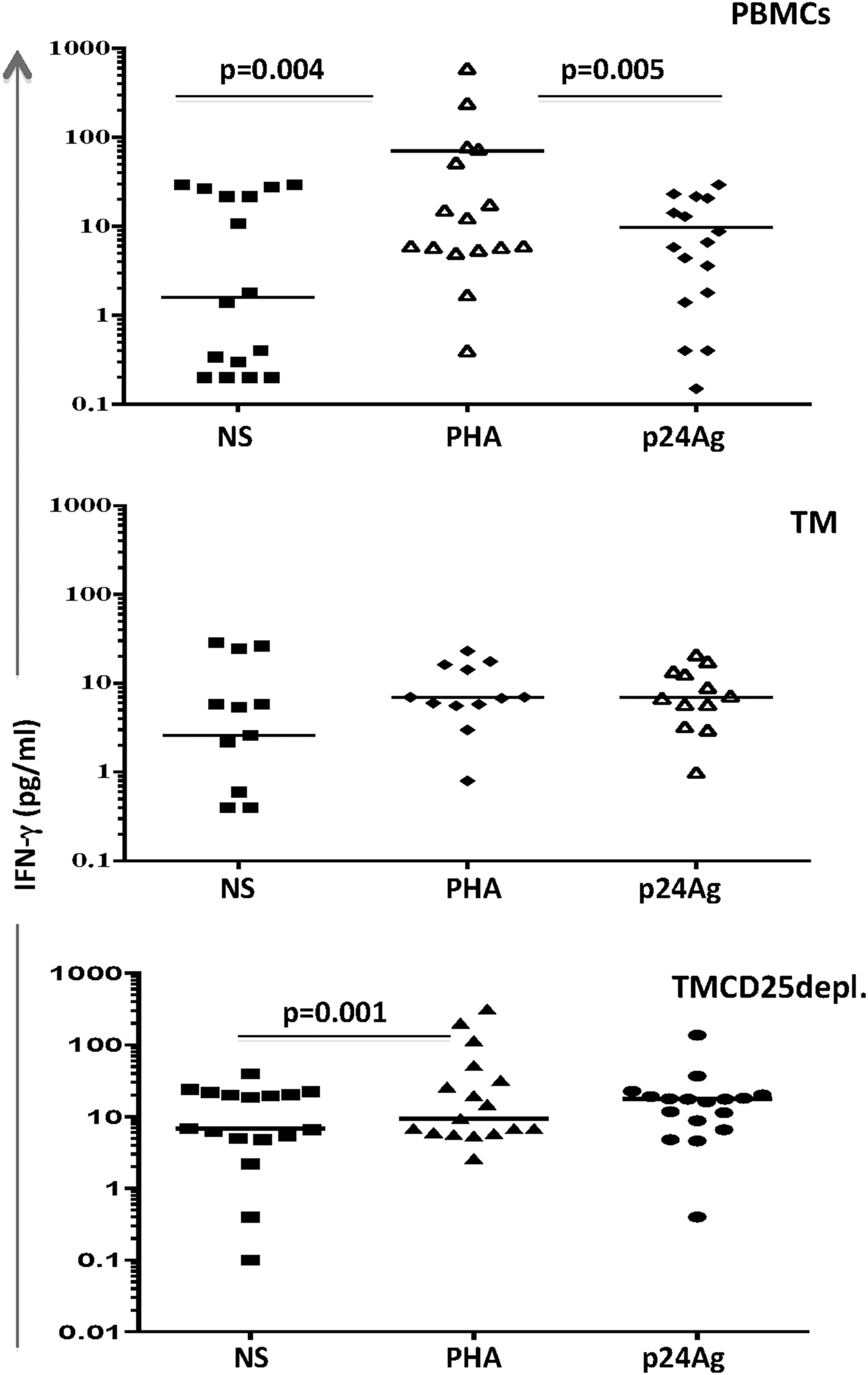
Inside group variations of the spontaneous and stimulated IFN-γ production. Dot plots represent individual data obtained from the evaluation of IFN-γ production in 17 HIV-1 seropositives. One below the other are shown PBMCs, TM (CD4+CD45RA−), and TMCD25 depleted cells. Every subset was cultured alone in the presence of 5 μg/mL PHA and 0.25 μg/mL HIV-1-p24Ag. Median values with range are presented, and the comparisons were performed by the nonparametric t-test, where p < 0.05 was considered significant.
Fine-tuning of the spontaneous lymphocyte proliferation by regulatory T cells
The short-term spontaneous lymphocyte proliferation (SLP-1h) was determined in nonstimulated cultures from the following fractions: (i) PBMCs, (ii)
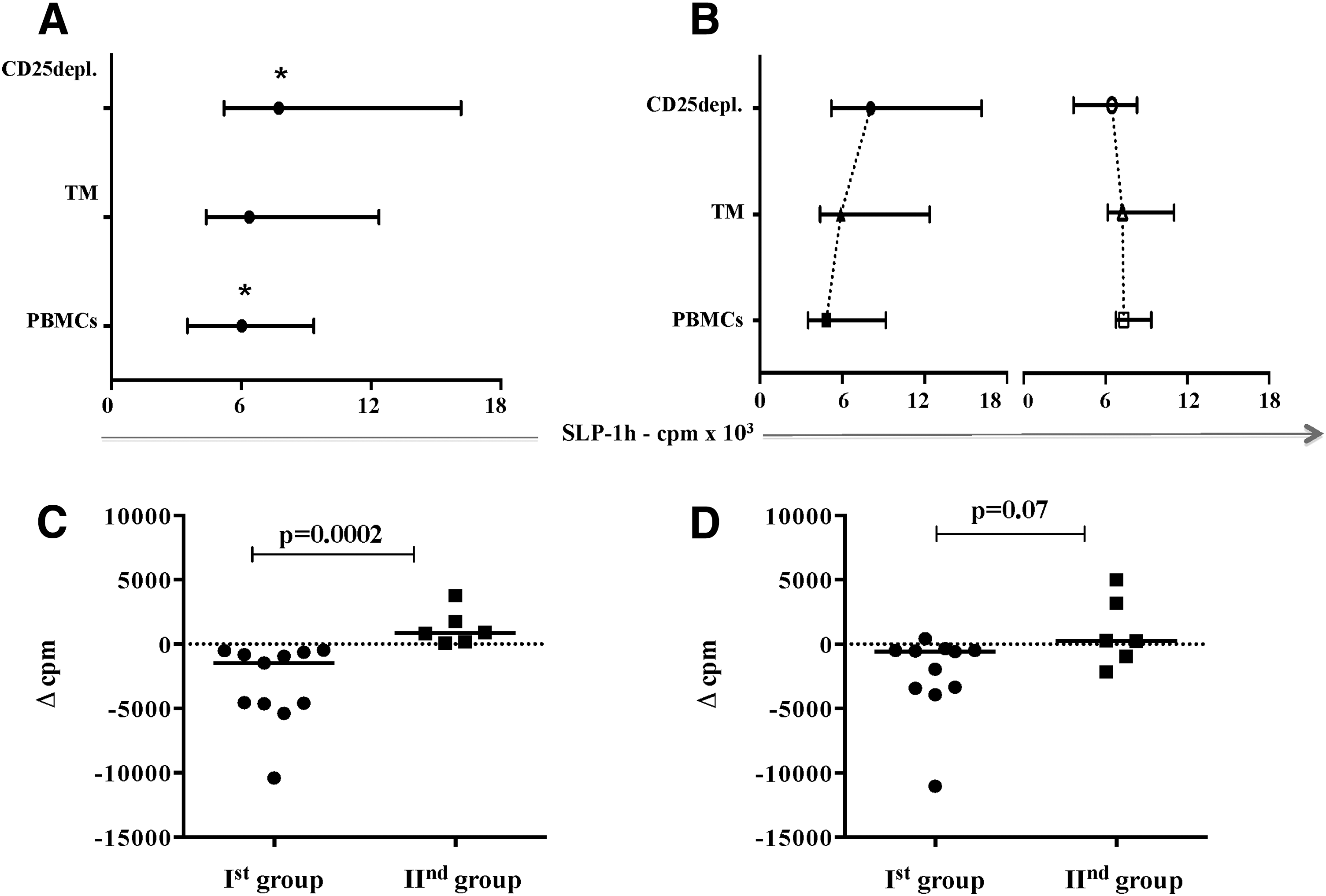
Variations in the SLP-1h among study populations. In all patients (n = 17), the degree of the short-term spontaneous lymphocyte proliferation (SLP-1h), expressed in cpm (counts per minute) of 3H-thymidine incorporation, showed a stepwise increase in each separate fraction, reaching highest values when Tregs were depleted
To determine both the direction and the extent to which SPL-1h has been changed between the fractions, we calculated a Δ value of SLP-1h between the three fractions for each patient as follows: (i) Δ1 = cpm(PBMCs)-cpm(
A gradual increase of the SLP-1h (Δ1 < Δ2 < Δ3) was found in controls. The highest values were found in the CD25depleted fraction (p = 0.008), while CD4+TM and PBMCs were lower with a slight predominance of SLP-1h(TM) (p = 0.02). In patients, we observed higher SLP-1h levels than in controls for all subsets (p < 0.05), but the trend of Δ values was not similar in all subjects. In 11 patients, it was comparable to those in controls, with negative Δ3 (Δ1 vs. Δ2, p = 0.01 and Δ2 vs. Δ3, p = 0.001). In the remaining six patients, no significant changes between fractions were observed, and Δ3 was positive. Accordingly, patients were divided into two groups as follows: the first one (Ist gr.) consisted of 11 and the second (IInd gr.) of six subjects (Fig. 4B). The comparison of the SLP-1h(PBMCs) showed higher values in the second group: median (range): Ist gr. 4,847 (3,519–9,185) cpm and IInd gr. 7,340 (6,292–9,358) cpm, p = 0.03.
The analysis of Δ3 values between groups showed significantly lower values in the Ist group: median (range): Δ3 (Ist gr.): −1,475 (−10,398 to −508.0) cpm versus Δ3 (IInd gr.): 721.8 (67.5 − 3,780) cpm, p = 0.0002. Thus it has been suggested that the contribution of CD25+ cells in the control of spontaneous proliferation is more sensitive in the first group, where not only Δ3 but also Δ2 values were found to be negative (Fig. 4C, D).
We correlated data obtained with clinical parameters like CD4 count, viral load, percentage of CD4+CD25+FoxP3+ cells, CD4+ nadir, and opportunistic/concomitant infections. The general overview of all 17 patients showed that although SLP-1h was not associated with the above parameters, the Δ3 was inversely dependent on the CD4 count at the timepoint of blood taking (Supplementary Fig. S3). The same analysis within groups did not show similar relation, but subjects in the first groups showed higher CD4+ count than those from the second group [median (range): Ist gr 995 (388–1,375) and IInd gr. 506 (144–962), p = 0.04], and CD4 nadir in the second group was lower compared to the first group [median (range): Ist gr. 229 (109–383) and IInd gr. 158 (69–330), p > 0.05]. Patients differed also by the viral load: 9 of 11 from the first group against 3 of 6 from the second one were with undetectable VL (<50 copies) at the time of sample taking. In the first group, one patient had HCV coinfection and in the second, one with Candida albicans. The results obtained were found to be independent from the treatment duration and hints that some patients, although having a good response to therapy (VL and CD4 count), could not efficiently restore T cell function.
Differential impact of the spIFN-γ production on the spontaneous lymphocyte proliferation
The overall analysis of samples showed that spIFN-γ has an unfavorable impact on the capacity of lymphocytes to proliferate. Particularly, the more detailed examinations within each patient group revealed a strong inverse correlation between the nonstimulated production of IFN-γ from PBMCs, CD4+TM, and TMCD25depl. and the respective proliferative activity in the first group of patients (r = −0.9, p = 0.003, r = −0.9, p < 0.0001, r = −0.8, p = 0.01, respectively) (Fig. 5A). On the contrary, in the second group the one and only correlation was found within the fraction of CD4+TM cells (r = −0.8, p = 0.03) (Fig. 5B).

Inside group analysis of SLP-1h dependence from spIFN-γ. In both groups of patients, the SLP-1h(PBMCs) was separately analyzed as function of the spIFN-γ, produced by PBMCs, TM, and TMCD25depl. On the
It was expected that this finding might be a result of qualitative differences between groups. The analysis showed no significant difference in spIFN-γ concentrations between groups in all fractions (Fig. 5C).
Tregs are weak suppressors of p24-antigen stimulated IFN-γ response
Afterward, we examined whether and to what extent the IFN-γ production in response to p24 Ag might be associated with the spontaneous lymphocyte proliferation. The overall analysis did not show a relationship between both parameters, which was not the case within both groups of patients. The more detailed examination showed alterations, specific for each of the above-mentioned groups. In the first group, within the population of PBMCs, the gradual increase of the free spINF-γ in supernatant was in accordance with the elevation of SLP-1h values in the three fractions, more specifically in PBMCs and TMCD25depl. cells. The Ag-specific IFN-γ showed a similar trend as the spIFN-γ. In contrast, the increase, observed in TMCD25depl. fraction, was weak and not significant: median (range) for PBMCs: 8.06 (0.4–17.28), TM: 13.10 (0.01–16.80), and TMCD25depl.: 17.80 (0.4–136.9) pg/mL (Fig. 6A). Collectively, the fact that in TMCD25depl. cells from the first group the SLP-1h and spIFN-γ significantly increased, while IFN-γ(p24) did not, indicated that the suppressive effect of Tregs might be effective mostly on the proliferation control than on the IFN-γ production. In patients from the second group, no relationship between both parameters was found (Fig. 6B). However, the case of PBMCs is particular for both examined groups, due to diverse cellular pattern undergoing activation and/or having the potential to produce IFN-γ, while the remaining two fractions are more prone to represent a pure model of the relations of interest. That was better evidenced in the first group of patients by the results of IFN-γ(PHA). The highest amount of free IFN-γ was detected in PBMCs and TMCD25depl. cultures from the first group. Of note, TM cells showed almost no free IFN-γ in the cell culture media: median (range) for PBMCs: 18.46 (5.0–478.3) pg/mL; TM: 5.8 (0.8–18.52); and TMCD25depl.: 14.80 (2.60–316.4) pg/mL, p < 0.05. In the second group, no variations were found (Fig. 6A, B).
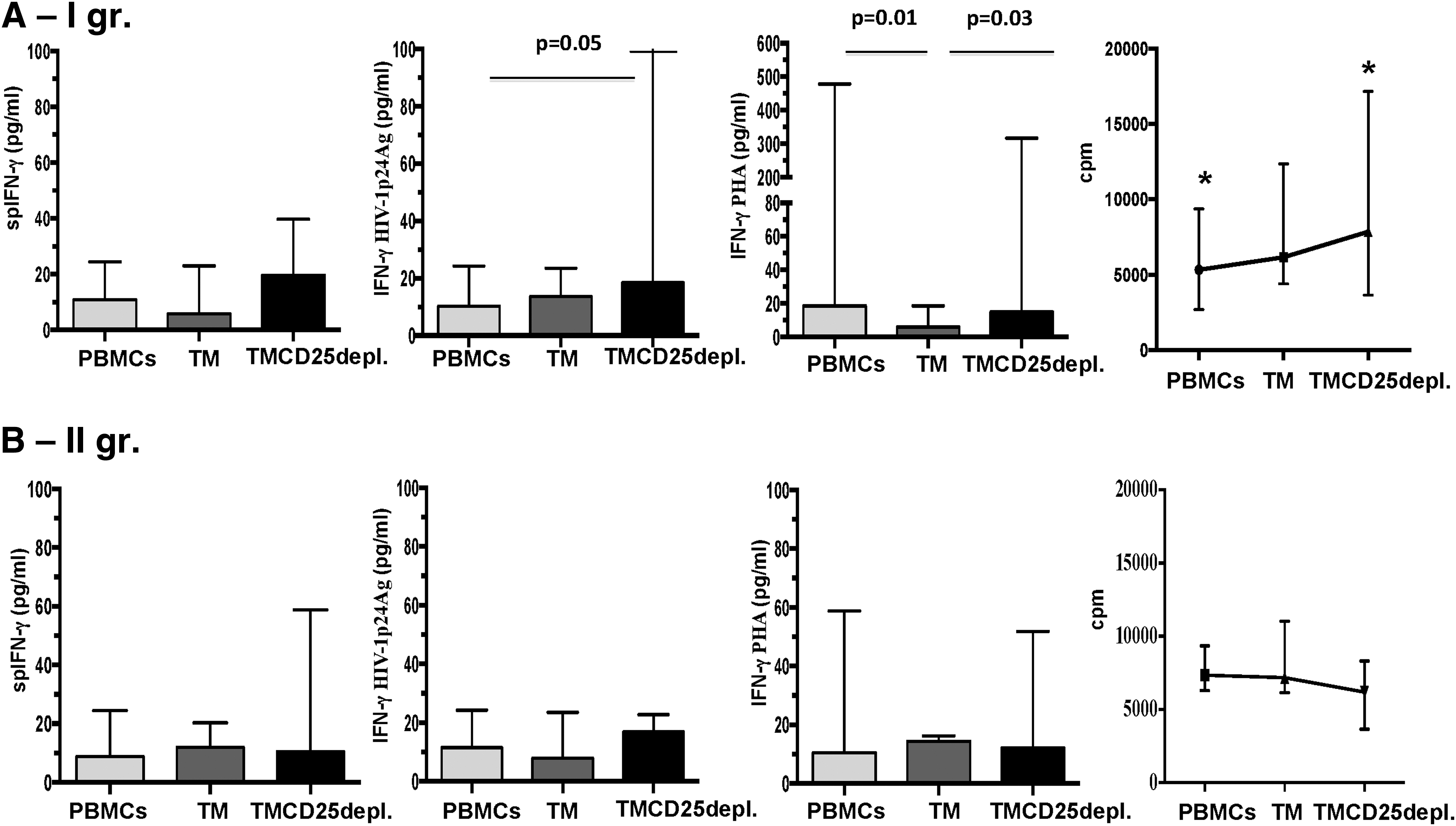
Concentrations of IFN-γ production by groups in the light of fraction driven changes in SLP-1h. The in-depth group analysis of IFN-γ concentrations in cell culture media from PBMCs, TM, and TMCD25depl. showed that in the Ist group
Evaluation of the free p24 antigen
Tregs are prone to be infected by HIV due to the surface expression of both HIV ligands. We hypothesized that the infection of Tregs might have an effect on their function. PBMCs, TM, TMCD25depl. fraction, and Tregs were cultured for 96 h with and without IL-2. From 17 HIV-1+ patients, only in one patient from the first group, free p24Ag was detected in Treg supernatant. In three other subjects, two from the Ist gr. and one from the IInd gr., p24 Ag was found in the cell culture media from TMCD25depl. cells. As it is shown in Table 1, in all of them a gradual elevation of SLP-1 from PBMCs to
IFN-γ, interferon-γ; PBMC, peripheral blood mononuclear cell.
Discussion
The role of Treg cells in the course of HIV-1 infection is extensively studied, but results obtained open more questions than providing answers. In the era of ART, the behavior and the impact of Tregs have become much more challenging. Till now, there has been no consensus about the quantitative parameters of Tregs in ART-treated patients. Our results demonstrate increased Treg percentages in comparison to controls. This finding is in agreement with other studies showing that ART-induced Treg increase is associated with a decrease in the immune activation and CD8 T cell apoptosis in complete responders, but not in nonresponder subjects (18,44).
We have specifically evaluated a particular aspect of Treg role in the immune activation network, the relationship between IFN-γ and Tregs. We found that Treg cells positively control the spontaneous lymphocyte proliferation and their presence significantly decreases it. At the same time, Tregs may not be considered as sufficiently strong inhibitors of both spontaneous and antigen-induced IFN-γ. Thus, although being in accordance with other studies, showing decreased proliferative capacity of the remaining T cells, our results do not fully support the negative impact of Tregs on the cytokine production from the conventional T cells (22,48). Furthermore, not all patients had one and the same result profile. In some of them (Ist group), the high proliferation values were accompanied with low spIFN-γ in PBMCs. These data corresponded well to the classical statement for the antiproliferative activity of IFN-γ. In the rest of the subjects (IInd group), both parameters were independent of each other.
To better assess the impact of Tregs on proliferation capacity, Tregs were removed and the proliferation of the remaining CD25-depleted non-FoxP3 cells was estimated by Δ3 = cpm(PBMCs)-cpm(TMCD25depl.) value. Results obtained showed that both groups were clearly defined; in the first, but not in the second group, the increase in SLP-1h between fractions was demonstrated by the negative Δ3, thus confirming the antiproliferative potential of Tregs. When comparing these data with clinical parameters for all subjects, the lowest Δ3 values were found in subjects with the highest CD4 count. However, the inside group analysis did not show similar results, the majority of subjects from the first group were with an undetectable VL at the time point of investigation (9 of 11 vs. 3 of 6) and a higher CD4 count. Logically, we asked the question: “why are these cells not able to fully express their function in some patients?” The most reasonable suggestion was the viral influence on Treg function. In very interesting studies, it has been shown that CD45RO+ Tregs might be infected with R5.HIV and CXCR4 strains and, consequently, undergo virus-mediated modulation in their function (2,9,20,30).
In the present study, the level of the free p24 antigen in the cell culture media from nonstimulated and stimulated Tregs was assessed. Our results confirmed that p24 Ag might be detected even in nonstimulated Tregs. It was increased under the appropriate stimulation with IL-2 when cells were not specifically transfected by the virus. Moreover, this fact was independent from the stage of infection, CD4+ T cell count, and the percentage of Tregs in the peripheral blood. This might probably be valid for induced Tregs, which belong to conventional, supposedly already infected CD4+T cells. The question seems much more intriguing considering the fact that the thymus becomes infected very early in the course of infection and the contamination of natural Tregs could not be excluded (35). Most probably, being infected with HIV and placed in an over activated milieu, Tregs are exposed to a broad spectrum of cytokines and also experience impairment of their suppressive potential. We also tried to associate the appearance of the free p24Ag in Treg cell culture media with the duration of the infection. The fact that positive results were obtained from patients with 6–14 years of infection is indicative that the attention might be drawn toward already established virus–host interactions. A confirmation of our suggestion comes from several studies, focused on viral induced transcriptome modifications in Treg genes—LEF-1, STAT5, and ETS-1, leading to downregulation of FoxP3 expression (1,17,25,31,33).
The second result stemming from our work is the increased spINF-γ production upon depletion of Treg cells, while p24 Ag and PHA-stimulated ones remained unchanged. Our preliminary hypothesis that Tregs are powerful controllers of the spontaneous and induced IFN-γ was reevaluated in light of the HIV-modulated immune dysfunction. Very early studies on HIV infection have shown a decreased capacity of CD4+ T cells to respond to recall and specific antigens and in a lesser extent to mitogens. Later on, it has been shown that this is a consequence of two major events, the direct viral impact and the effect of bystander activation, which leads to cell exhaustion (4,19). Our previous study showed that subjects, able to maintain IL-2 production, have a low level of immune activation and well-established IL-10 production (42). Accordingly, it might be proposed that decreased IL-2 would impair Treg function, as well. Another hypothesis is based on the impairment of SOCS-1/Treg cross talk by HIV. SOCS-1 (suppressor of cytokine signaling-1) has been shown to play an important role in the regulation of inflammation and autoimmunity (11). The key moment is the parallel increase of FoxP3 and microRNA-155, associated with downregulation of SOCS-1 (27). Despite the fact that this mechanism is more applicable to the thymic Tregs, the absence of SOCS-1 was associated with increased production of the pro-inflammatory IL-17 and IFN-γ by FoxP3+ Tregs in the periphery (40). In HIV infection, SOCS impacts both the immune system and the virus. The precise experiments of Miller et al. have shown increased SOCS-1 and SOCS-3 mRNA levels in HIV-positive individuals, but reduced SOCS-1 and SOCS-3 protein levels together with increased IFN-γ mRNA and immune activation compared to T cells from HIV-negative volunteers (29). Besides, SOCS-1 directly binds to the nucleocapsid regions on the HIV Gag polyprotein, enhancing Gag stability promoting intracellular trafficking and new viral particle production. That is the reason why low SOCS-1 levels are associated with the inability to hamper the excessive activation (32,36,37).
Treg cells are extensively studied in autoimmune pathology and transplantation, where they are of critical importance. Data from autoimmune deviations show an accelerated onset and more severe form of collagen-induced arthritis, associated with a functional defect in CD4+CD25+ Tregs in mice lacking IFN-γR (21), together with deficiency in STAT1 in the experimental autoimmune encephalomyelitis, due to reduced number and impaired function of Tregs in mice (47). Studies on alloantigen-reactive Treg cells clearly demonstrate that IFN-γ is critical for conversion of CD4+CD25−T cells to CD4+ Tregs (26,39,46). Also, it has been shown that IFN-γ/SOCS signaling negatively regulated the expansion of Tregs (24,25). Data from animal experiments have shown that SOCS1 deletion specifically in Tregs induces the development of spontaneous dermatitis, splenomegaly, and lymphadenopathy, suggesting a defective Treg function (41).
Recent analysis on gene expression profiles in HIV-1-infected individuals showed that blocking of IFN-γ reduces the suppressive effect of allo-specific Tregs, thus suggesting that defective expression of IFN-γ receptor and/or signaling pathway might occur in some pathological conditions, like HIV. HIV significantly changes the cellular machinery and signal transduction. In a recently published large study, the authors identified 4,157 differentially expressed genes in patients with HIV infection and showed that the major components of JAK-STAT signaling were downregulated [i.e., IFNG, JAK2, STAT1, STAT2, and IRF9) (28)].
In conclusion, data from the present study provide another point of view on the role of IFN-γ as a factor that may reconcile the cross talk between Treg and non-Treg cells and are in agreement with the latest achievements in autoimmunity. The infection-associated overactivation and proliferation may be encouraged by the twofold effect of HIV-1 on the T cell regulatory arm, throughout virus–host cell interactions and by an impairment of IFN-γ production and signaling. In addition, our results stimulate the hypothesis that Tregs are mostly effective in the proliferation control to establish immune quietness rather than immune suppression (8). Further investigations are needed to better clarify the precise mechanisms involved during the natural course of HIV infection and the impact of ART. Finally, this work indicates that Treg cells are not only an important player in the immunopathogenesis of HIV-1 infection but also have a significant potential for therapeutic manipulations.
Footnotes
Acknowledgments
The authors thank Maya Kicheva and Yana Todorova for their helpful work and all patients and controls for their collaboration. This work was supported by Grant No. 1407/04 of National Science Fund, Ministry of Education and Science, Republic of Bulgaria.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.