
Abstract
Porcine circovirus type 2 (PCV2) adenovirus vaccine has been reported, but strong immune responses induced by adenovirus vector can decrease vaccine efficacy. To reduce the immunogenicity of adenovirus proteins, in previous study, we constructed the PCV2 adenovirus vaccine either modified with human cytomegalovirus first intron (Intron A) and woodchuck hepatitis virus post-transcriptional regulatory element (WPRE) to increase the expression of Cap, or coexpressed porcine tumor necrosis factor-related activate protein (CD40L) and granulocyte macrophage colony-stimulating factor (GMCSF) to improve the immunogenicity of PCV2 Cap adenovirus vaccine. All these vaccines were evaluated in mice. In the present study, the protective immune responses of Intron A/WPRE-modified recombinant adenovirus Ad-A-C-W and CD40L/GMCSF-modified recombinant adenovirus Ad-CD40L-Cap-GMCSF were evaluated in pigs. Enzyme-linked immunosorbent assay and virus neutralization assay showed that both Ad-A-C-W and Ad-CD40L-Cap-GMCSF could induce a higher specific antibody and neutralizing antibody than Ad-Cap (p < 0.05). Lymphocyte proliferation assay and cytokine release assay showed that Ad-A-C-W and Ad-CD40L-Cap-GMCSF induced a stronger cellular immune response than Ad-Cap. The PCV2 challenge experiment showed that viral loads of Ad-A-C-W-vaccinated group and Ad-CD40L-Cap-GMCSF-vaccinated group were lower than Ad-Cap vaccinated group (p < 0.05) after pigs were oronasally challenged with 5 × 105 TCID50 PCV2. Autopsy and histopathological examination showed that no obvious clinical and microscopic lesions were observed in groups Ad-Cap, Ad-A-C-W, and Ad-CD40L-Cap-GMCSF. Taken together, the results demonstrated that two modified recombinant adenovirus vaccines (Ad-A-C-W and Ad-CD40L-Cap-GMCSF) induced stronger humoral and cellular immune responses and provided better protection than unmodified adenovirus Ad-Cap. Therefore, Ad-A-C-W and Ad-CD40L-Cap-GMCSF would be used as potential vaccines for prevention and control of PCV2 infection.
Introduction
P
The adenovirus expression system, one of the most widely used viral expression systems, shows more advantages than other systems, such as easily constructing, higher efficiency of gene transfer, higher titers, and inducing humoral, mucosal, and cellular immune responses together (3,8,10,14,17,24). However, strong immune responses induced by adenovirus proteins (mainly penton and hexon proteins) result in poor efficacy (32). Although the adenovirus vector system expressing PCV2 Cap has been studied, the vaccine efficacy is limited (27,28). So, it is necessary to reduce the immunogenicity of adenovirus itself. In the previous study, we have verified that either inserting largest intron of the human cytomegalovirus (Intron A) and woodchuck hepatitis virus post-transcriptional regulatory element (WPRE) into adenovirus vector, or coexpression of CD40 ligand (CD40L) and granulocyte macrophage colony-stimulating factor (GMCSF) can increase adenovirus vector vaccine efficacy in mice (15,16).
In this study, the immune responses of recombinant adenoviruses Ad-A-C-W and Ad-CD40L-Cap-GMCSF were further evaluated in pigs. The results demonstrated that two kinds of recombinant adenovirus vaccines could induce stronger humoral and cellular immune responses and provide better protective efficacy than unmodified adenovirus Ad-Cap.
Materials and Methods
Viruses and cell lines
The AdEasy™ adenovirus vector system derived from human adenovirus serotype 5 was bought from Agilent Technologies (La Jolla). Recombinant adenoviruses Ad-Cap, Ad-A-C-W, Ad-CD40L-Cap-GMCSF and empty vector adenovirus Ad-blank (human adenovirus serotype 5) were cultured and propagated in human embryonic kidney 293 cells (HEK293) (ATCC, CRL-1573) as previously described (15,16), and viral titers of these adenoviruses were all about 1010 TCID50/mL. Porcine kidney 15 cells (PK15), which were PCV1-free, were used to propagate PCV2 (GenBank No. EU366323).
Animal experiments
Thirty-five commercial 28-day-old Yorkshire pigs without the PCV2 antibody were randomly divided into seven groups each with five pigs and housed in seven separate rooms. All pigs used in the experiment were confirmed to be free of porcine parvovirus, porcine reproductive and respiratory syndrome virus, and classical swine fever virus by routine serological testing. Pig experiments were approved by the Animal Ethics Committee of North West Agriculture and Forestry University. Groups 1–3 were immunized intramuscularly with 1010 TCID50 of Ad-Cap, Ad-A-C-W, and Ad-CD40L-Cap-GMCSF, respectively; group 4 was vaccinated intramuscularly with commercial inactivated vaccine (inactivated virus of PCV2b [SH Strain]) according to the manufacturer's instruction (Qianyuanhao, Nanjing, China); group 5 was immunized intramuscularly with 1010 TCID50 of empty vector adenovirus Ad-blank (human adenovirus serotype 5); group 6 was vaccinated with phosphate-buffered saline (PBS) (nonvaccinated but challenged group); group 7 was used as untreated control (nonvaccinated and nonchallenged group). Booster immunization was conducted on 14 days postimmunization (dpi) under the same condition. On 28 dpi, groups 1–6 were challenged oronasally with 5 × 105 TCID50 PCV2 per pig. All animals were monitored for 63 days after challenge. Rectal temperatures and clinical signs were detected weekly. Serum samples of each group were collected on 0, 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi for enzyme-linked immunosorbent assay (ELISA) and virus neutralization assay (VN). On 28 dpi, swine peripheral blood lymphocytes were used for further lymphocyte proliferation assay and cytokine (IFN-γ and IL-13) release assay. On 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi, serum samples were collected for detecting viremia by quantitative real-time PCR (Q-PCR). On 91 dpi, all animals were euthanized, and superficial inguinal lymph nodes and lungs were collected for histopathological examination.
Enzyme-linked immunosorbent assay
Sera of five pigs from each group were collected from the precava on 0, 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi for detecting specific antibody by using commercial PCV2 antibody ELISA detection kits (Yuanye Biotechnology Co., Shanghai, China) according to the manufacturer's instruction.
Virus neutralization assay
The PCV2 neutralizing antibody was tested by VN. Briefly, serum samples were diluted in twofold, starting with 1:2. Then, 100 TCID50 PCV2 was added and incubated for 1 h at 37°C. Then, the serum–virus mixture was added into PK15 as previously described (5). After 72 h, PK15 cells were fixed with cold absolute ethanol at 4°C for 15 min, incubated with the rabbit anti-PCV2 antibody, and stained with FITC-conjugated goat anti-rabbit IgG (Boshide, Wuhan, China). Titers were conducted as the reciprocal of the last serum dilution at 80% or greater than 80% fluorescent focus reduction by the fluorescent microscope.
Lymphocyte proliferation assay
On 28 dpi, lymphocytes were aseptically separated from the swine peripheral blood with pig lymphocyte separation medium (Dakewe, Shenzhen, China) following the manufacturer's instruction. Lymphocytes were seeded into a 96-well plate and stimulated for 68 h with concanavalin A (ConA, 5 μg/mL; Sigma, Los Angeles) as positive control, Cap protein (20 μg/mL) as the stimulant, and RPMI-1640 as negative control. Then, 20 μL of MTT (3-[4, 5-dimethylthiazol-2-y]-2, 5-diphenyltetrasodium bromide tetrazolium, Sigma, 5 mg/mL in PBS) was added per well with further incubation for 4 h. Dimethyl sulfoxide was added 100 μL per well to stop the reactions. The OD570 nm was detected and the stimulation index (SI) was determined as follows: SI = mean OD of Cap-stimulated cells/mean OD of unstimulated cells.
Cytokine release assay
Lymphocytes were separated aseptically from the swine peripheral blood of five pigs from each group as above on 28 dpi. Then, the lymphocytes were seeded into a 96-well plate with 2 × 105 cells per well. RPMI-1640 alone and 2 μg Cap protein were added into each well and the supernatants were collected 72 h later. Titers of IFN-γ and IL-13 were detected by pig IFN-γ and pig IL-13 ELISA Kits (ExCell Bio, Shanghai, China) following standard procedures. The concentrations of IFN-γ and IL-13 in the supernatants were calculated by the standard curves.
Detection of viremia in blood
On 28 dpi, groups 1–6 were challenged with 5 × 105 TCID50 PCV2. Following challenge, all pigs were monitored daily for clinical signs, including sneezing, lethargy, and coughing. Serum samples were collected on 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi. Virus DNA was extracted from serum samples and detected by Q-PCR established previously (29). Briefly, PCV2 genomic DNA was extracted by DNA extraction kits (Axygen, Hangzhou, China). The concentrations of plasmid containing PCV2 genomic DNA were determined by the NanoDrop 2000 Spectrophotometer (Thermo Scientific). Copy number was calculated using the following formula: amount (copies/μL) = 6.02 × 1023 (copies/mol) × concentration (g/μL)/MW (g/mol). The viral copy number in samples per μL was tested by real-time PCR with serially diluted plasmids from 102 to 108 copies/μL. Real-time PCR was performed with primers (forward 5′-AAGGGCTGGGTTATGGTATGT-3′ and reverse 5′-CGCTGGAGAAGGAAAAATGG-3′). Fluorescence data were recorded in real time with an IQ5 multicolor real-time PCR detection system (Bio-Rad). The viral loads were determined as the mean of the logarithmic DNA copy number divided by mL (Lg copies/mL). The sensitivity of Q-PCR used in the study was 1,000 copies/mL.
Histopathological examination
The gross lesions of lungs and lymph nodes were detected after necropsy. Superficial inguinal lymph nodes and lungs were removed and fixed in 10% buffered formalin. The fixed samples were dehydrated and embedded in paraffin wax. Consecutive 5-μm-thick sections were cut and processed for hematoxylin and eosin staining (HE). Microscopic pathological lesions were conducted as − (normal), + (mild), ++ (moderate), and +++ (severe) as previously described (28).
Statistical analyses
All data were treated as the mean ± standard error of mean (SEM). The ELISA, VN, lymphocyte proliferation assay, cytokine release assay, and Q-PCR results were assessed by one-way analysis of variance (ANOVA) and the Student–Newman–Keuls (SNK) test; temperature was assessed by two-way repeated measures ANOVA and the SNK test. All statistical analyses were conducted with SPSS 19.0 software (SPSS, Inc., Chicago, IL). A value of p < 0.05 was considered as a significant statistical difference.
Results
Temperature detection
Rectal temperatures of all pigs were detected weekly since the first immunization. As shown in Table 1, temperatures of all groups were normal before 28 dpi. However, temperatures of Ad-blank group and PBS group were significantly higher than the Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group from 35 to 56 dpi (p < 0.05), and the mean temperatures of both the Ad-blank group and PBS group exceeded 39.5°C and even reached 40°C at a certain time point. The temperatures of Ad-blank group and PBS group gradually decreased since 63 dpi and got back within 39.5°C on 77 dpi. Pigs of Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, SH-strain group, and untreated group kept normal temperature (38°C–39.5°C) after challenge.
Rectal temperatures of different groups were detected weekly after immunization. Data are presented as the mean ± SEM. Treatments with different lowercase letters (a–c) indicate statistically significant differences between the indicated groups (p < 0.05).
GMCSF, granulocyte macrophage colony-stimulating factor; PBS, phosphate-buffered saline; SEM, standard error of mean.
Humoral immune response evaluation in pigs
Specific antibody to PCV2 detection in pigs
Anti-PCV2 -specific antibody titers of different groups were evaluated on 0, 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi by commercial ELISA kits as per the manufacturer's instruction. As shown in Table 2, specific antibody titers of Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF- vaccinated group, and SH-strain group were initially detected on 14 dpi and maintained till 63 days. On 28 dpi, the specific antibody of Ad-CD40L-Cap-GMCSF-vaccinated group was higher than others (p < 0.05), Ad-A-C-W-vaccinated group was higher than Ad-Cap-vaccinated group (p < 0.05), but the Ad-A-C-W-vaccinated group was significantly lower than SH-strain group (p < 0.05). On 35, 49, 56, 63, and 70 dpi, the specific antibody of SH-strain group was significantly higher than other groups (p < 0.05), specific antibody of Ad-CD40L-Cap-GMCSF-vaccinated group was higher than Ad-Cap-vaccinated group (p < 0.05) but was lower than Ad-A-C-W-vaccinated group (p < 0.05). On 42 and 77 dpi, specific antibody of SH-strain group was higher than other groups (p < 0.05), specific antibodies of Ad-CD40L-Cap-GMCSF-vaccinated group and Ad-A-C-W-vaccinated group were not different (p > 0.05), but both were significantly higher than Ad-Cap-vaccinated group (p < 0.05). Ad-blank and PBS groups developed certain specific antibodies on 42, 49, 56, 63, and 70 dpi.
PCV2-specific antibodies of different groups were detected weekly after immunization. Data are presented as the mean ± SEM. Treatments with different lowercase letters (a–f) indicate statistically significant differences between the indicated groups (p < 0.05).
ELISA, enzyme-linked immunosorbent assay; PCV2, porcine circovirus type 2.
Neutralizing antibody level detection in pigs
Neutralizing antibody titers were detected on 0, 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi by VN. As shown in Table 3, neutralizing antibodies of Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group were initially detected on 14 dpi and maintained 63 days. On 28 dpi, neutralizing antibodies of Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group were significantly higher than Ad-Cap-vaccinated group (p < 0.05), but there were no differences among them (p > 0.05). On 35 dpi, neutralizing antibody of SH-strain group was higher than other groups (p < 0.05), Ad-CD40L-Cap-GMCSF-vaccinated group was higher than Ad-Cap-vaccinated group (p < 0.05), but was not different from Ad-A-C-W-vaccinated group (p > 0.05). On 42, 49, and 63 dpi, neutralizing antibody of SH-strain group was higher than Ad-Cap-vaccinated group (p < 0.05), but was not different from Ad-CD40L-Cap-GMCSF-vaccinated group and Ad-A-C-W-vaccinated group (p > 0.05). There was no difference among Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, and Ad-CD40L-Cap-GMCSF-vaccinated group (p > 0.05). On 56 dpi, neutralizing antibody of SH-strain group was higher than other groups (p < 0.05), there was no difference among Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, and Ad-CD40L-Cap-GMCSF-vaccinated group (p > 0.05). On 70 dpi, neutralizing antibody of SH-strain group was higher than other groups (p < 0.05), Ad-A-C-W-vaccinated group was higher than Ad-Cap-vaccinated group and Ad-CD40L-Cap-GMCSF-vaccinated group (p < 0.05). On 77 dpi, neutralizing antibody of SH-strain group was not different from Ad-A-C-W-vaccinated group and Ad-CD40L-Cap-GMCSF-vaccinated group (p > 0.05). And there was no difference between Ad-CD40L-Cap-GMCSF-vaccinated group and Ad-Cap-vaccinated group (p > 0.05). Besides, Ad-blank and PBS groups developed certain neutralizing antibodies on 42, 49, 56, 63, 70, and 77 dpi.
Neutralizing titers are reciprocals of the highest dilution of serum that yielded 80% reduction in virus infectivity. Data are presented as the mean ± SEM. Treatments with different lowercase letters (a–d) indicate statistically significant differences in neutralizing antibody levels between the indicated groups (p < 0.05).
IFA, immunofluorescence assay.
Cellular immune response detection in pigs
Lymphocyte proliferative responses
Swine peripheral blood lymphocyte proliferative levels of different groups were detected on 28 dpi. As shown in Figure 1, SI of Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group was significantly higher than that of Ad-blank and PBS-treated pigs (p < 0.05). Ad-CD40L-Cap-GMCSF-vaccinated group and SH-strain group showed higher SI than other groups (p < 0.05), and Ad-A-C-W-vaccinated group was higher than Ad-Cap-vaccinated group (p < 0.05).
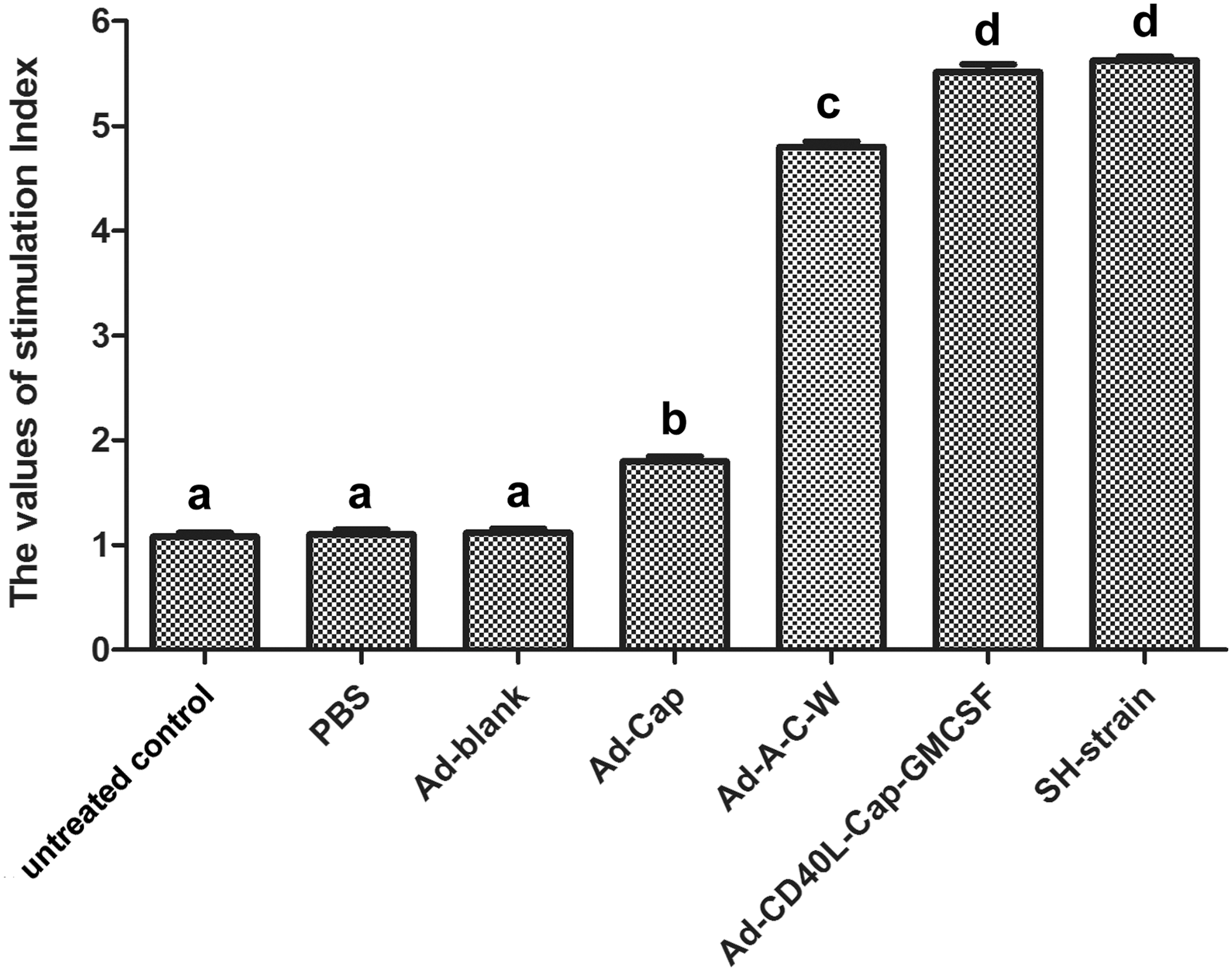
Lymphocyte proliferation levels of swine peripheral blood lymphocytes after specific stimulation. After immunization with different recombinant adenoviruses, swine peripheral blood was collected and lymphocytes were pooled for lymphocyte proliferation assay. Data are presented as the mean ± SEM. Treatments with different lowercase letters (a–d) indicate statistically significant differences in SI between the indicated groups (p < 0.05). GMCSF, granulocyte macrophage colony-stimulating factor; SEM, standard error of mean; SI, stimulation index.
Cytokine detection by ELISA
On 28 dpi, IFN-γ and IL-13 secretion levels from swine peripheral blood lymphocytes were measured to evaluate cellular immune responses. As shown in Figure 2, IFN-γ and IL-13 were higher in Ad-Cap-vaccinated group, Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group than that in Ad-blank group and PBS group (p < 0.05). The Ad-CD40L-Cap-GMCSF-vaccinated group and SH-strain group showed higher IFN-γ and IL-13 levels than other groups (p < 0.05), and Ad-A-C-W-vaccinated group was higher than Ad-Cap-vaccinated group (p < 0.05).
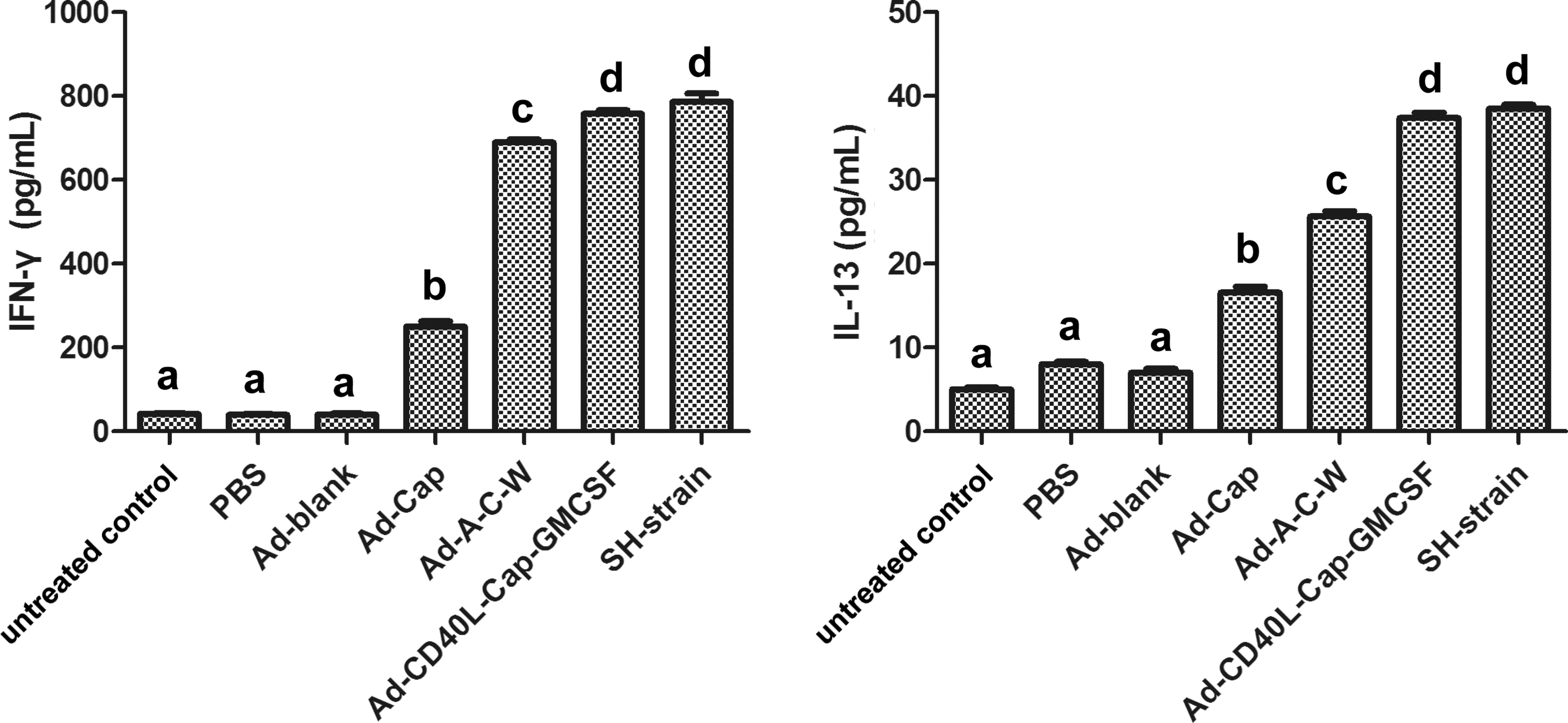
Levels of IFN-γ and IL-13 produced from lymphocytes in immunized pigs. On 28 dpi, swine peripheral blood lymphocytes from each group were harvested, cultured in 96-well plates, and stimulated with purified Cap. IFN-γ and IL-13 production in the supernatant was analyzed by ELISA. Data represent the mean ± SEM. Treatments with different lowercase letters (a–d) indicate statistically significant differences in cytokine levels between the indicated groups (p < 0.05). ELISA, enzyme-linked immunosorbent assay.
Viral load detection in serum samples
Virus DNA extracted from swine peripheral blood of seven groups was measured on 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi. As shown in Table 4, Viremia was undetectable on 28 dpi. On 35, 42, 49, 70, 84, and 91 dpi, the viral loads of Ad-blank group and PBS group were detectable and higher than other vaccinated groups (p < 0.05). Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group showed lower viral loads than Ad-Cap-vaccinated group (p < 0.05). Ad-A-C-W-vaccinated group did not differ from SH-strain group (p > 0.05), but both were lower than Ad-CD40L-Cap-GMCSF-vaccinated group (p < 0.05). On 56, 63, and 77 dpi, the viral loads of Ad-blank group and PBS group were higher than other vaccinated groups (p < 0.05). Ad-A-C-W-vaccinated group, Ad-CD40L-Cap-GMCSF-vaccinated group, and SH-strain group showed lower viral loads than Ad-Cap-vaccinated group (p < 0.05). Ad-A-C-W-vaccinated group was higher than SH-strain group (p < 0.05), but both were lower than Ad-CD40L-Cap-GMCSF-vaccinated group (p < 0.05).
Pigs were challenged with 5 × 105 TCID50 PCV2 on 28 dpi, and PCV2 viral loads were detected by Q-PCR in serum samples collected from pigs on 28, 35, 42, 49, 56, 63, 70, 77, 84, and 91 dpi. Data are presented as the mean ± SEM. The viral loads were determined as the mean of the logarithmic DNA copy number divided by mL (Lg copies/mL). The sensitivity of Q-PCR in our study was 1,000 copies/mL. Treatments with different lowercase letters (a–f) indicate statistically significant differences in viral loads between the indicated groups (p < 0.05). ND refers to not detected.
Histopathological examination in tissues
Pigs of groups 1–6 were challenged with PCV2 on 28 dpi and all pigs were executed on 91 dpi. Superficial inguinal lymph nodes and lungs were collected to conduct histopathological examination. The microscopic lesions were observed by an optical microscope. All pigs of groups 1–6 showed PCVAD pathologic changes in different degrees. The main gross lesions of lungs showed diffuse interstitial pneumonia, swelling, volume augmented, interstitial broadening, scleroid, and varying sizes of brown area scattered on the surface. The main gross lesions of lymph nodes were swelling to varying degrees, and cut surface color of lymph nodes was incanus or kermesinus, cut surface was moist, soft, and sappiness, sometimes even bleeding. The microscopic lesions of lungs were mainly interstitial pneumonia and inflammatory cell infiltration. The alveolar septa of groups 5 and 6 were significantly incrassate and infiltrated by many inflammatory cells, but groups 1–4 were mildly incrassate. The microscopic lesions of lymph nodes showed lymphoid depletion and inflammatory cell infiltration. Lymphoid follicles of groups 5 and 6 were significantly lymphocytes-lacking and many inflammatory cells infiltration, but lymphoid follicles of groups 1–4 were structural integrity, few lymphocytes decreasing and inflammatory cells infiltration (Fig. 3). The histological lesions of lungs and lymph nodes from different groups are summarized in Table 5.

The histopathological features of lungs and lymph nodes from the challenged pigs.
−, normal; +, mild lesions; ++, moderate lesions; +++, severe lesions.
Discussion
Many measures were taken to prevent PCVAD, and vaccination was considered to be the most effective one. Adenovirus vector system is one of the most effective virus vectors and has been used to express various proteins (4,25). Adenovirus vector system expressing PCV2 Cap protein has been studied, but has limited immune efficacy (27,28). One possible reason is that many proteins expressed by adenovirus itself could induce strong immune responses, resulting in poor efficacy (32). So it is necessary to reduce the immunogenicity of adenovirus proteins.
In previous study, we have constructed Intron A and/or WPRE-modified PCV2 adenovirus vaccines and evaluated the vaccine efficacy in mice and showed that Intron A and WPRE synergistically increase Cap expression and enhance the immune responses to Cap protein (15). Besides, we constructed CD40L and/or GMCSF-modified PCV2 adenovirus vaccines and evaluated their efficacy in mice. The results show that CD40L and GMCSF improve the immunogenicity of adenovirus vector vaccines in mice (16). Although the results of mice experiments of Ad-A-C-W and Ad-CD40L-Cap-GMCSF are impressing, the difference in body structures between mice and pigs makes it difficult to determine the efficacy of these vaccines in PCV2-infected pigs. So, in the present study, we conducted the pig experiment to evaluate the protective immune responses of Ad-A-C-W and Ad-CD40L-Cap-GMCSF.
Both humoral and cellular immune responses were evaluated in this study. Anti-PCV2 antibodies measured by ELISA and VN indicated that the humoral immunity was induced. The specific antibody and neutralizing antibody levels of Ad-A-C-W- and Ad-CD40L-Cap-GMCSF-vaccinated group were significantly higher than Ad-Cap-vaccinated group (p < 0.05), while those of Ad-CD40L-Cap-GMCSF-vaccinated group were lower than Ad-A-C-W-vaccinated group (p < 0.05), except for a certain time point. The Ad-A-C-W-vaccinated group and Ad-CD40L-Cap-GMCSF-vaccinated group elicited higher humoral immunity than Ad-Cap-vaccinated group. The possible reason for different humoral immune responses induced by Ad-A-C-W and Ad-CD40L-Cap-GMCSF was that Intron A and WPRE mainly increased the expression of Cap (30,31), but CD40L and GMCSF played an important role in the process of immune response (16).
Lymphocyte proliferation and cytokine levels are two important indexes of cellular immunity, both were detected on 28 dpi. Both Ad-A-C-W and Ad-CD40L-Cap-GMCSF could induce a stronger cellular immune response than Ad-Cap, but Ad-CD40L-Cap-GMCSF could induce a stronger cellular immune response than Ad-A-C-W in pigs. It is well known that CD40L is mainly expressed by activated CD4+ T cells and interacts with its receptor CD40, which plays important roles in both humoral and cellular immune responses, such as Th1 type response development (1,7), Ig type switching, antibody production (6,21), and interactions of between T cell and dendritic cell or T cell and macrophage (33). In addition to CD40L, GMCSF can stimulate neutrophil, monocyte/macrophage, and eosinophil colony formation, and enhance antibody-dependent cell-mediated cytotoxicity (18,19).
PCV2 challenge experiment showed that from 35 to 91 dpi, the viral loads of Ad-A-C-W-vaccinated group and Ad-CD40L-Cap-GMCSF-vaccinated group were lower than Ad-Cap-vaccinated group (p < 0.05). Gross lesions of lungs of group Ad-Cap, Ad-A-C-W, and Ad-CD40L-Cap-GMCSF exhibited slightly interstitial pneumonia, and lymph nodes were also slightly swelling. Microscopic examination showed no obvious lesions. The results demonstrated that recombinant adenoviruses could protect pigs from PCV2 challenge. Besides, Ad-A-C-W and Ad-CD40L-Cap-GMCSF induced better protective immune responses than Ad-Cap.
The results of pig experiment showed that the vaccine efficacy of adenoviruses Ad-A-C-W and Ad-CD40L-Cap-GMCSF was no better than the commercially inactivated vaccine. The possible reasons were as follows. The preparative techniques of commercially inactivated vaccines are mature; inactivated vaccines contain high content of antigen; there are different kinds of immunologic adjuvants for inactivated vaccines; and the whole virus particle could induce stronger immune responses than single protein, Cap. Although many researches focus on adenovirus vector vaccines, there are still some key techniques unsolved, for example, immunologic adjuvants of adenovirus vector vaccines are immature. So, to improve PCV2 adenovirus vaccine efficacy, further study must be focused on parameter optimization, such as the dose and route of delivery, vaccination procedures, and adenovirus adjuvants.
In conclusion, although the immune responses induced by Ad-A-C-W and Ad-CD40L-Cap-GMCSF were no better than inactivated vaccine, the adenovirus vector still has many advantages, for example, high transduction efficiency, wide tropism, high safety, and can infect stationary and division phase cells, making Ad-A-C-W and Ad-CD40L-Cap-GMCSF to be promising vaccine candidates to prevent and control PCV2 infection.
Footnotes
Acknowledgments
This research was supported by the Key Project of Shaanxi province Science and Technology Innovation Team (2013KCT-28), the National Natural Science Foundation of China (Nos.: 31372401, 31372411), and the Science and Technology Innovation Project in Shaanxi province (2016KTCL02-13).
Author Disclosure Statement
No competing financial interests exist.