
Abstract
We previously generated an oral hepatitis C virus (HCV) vaccine using Bifidobacterium displaying the HCV nonstructural protein 3 (NS3) polypeptide. NS3-specific cellular immunity is important for viral clearance and recovery from HCV infection. In this study, we enhanced the cellular immune responses induced by our oral HCV vaccine, Bifidobacterium longum 2165 (B. longum 2165), by combining interferon-α (IFN-α) as an adjuvant with the vaccine in a mouse experimental model. IFN-α is a widely used cytokine meeting the standard of care (SOC) for HCV infection and plays various immunoregulatory roles. We treated C57BL/6N mice with B. longum 2165 every other day and/or IFN-α twice a week for a month and then analyzed the immune responses using spleen cells. We determined the induction of NS3-specific cellular immunity by cytokine quantification, intracellular cytokine staining, and a cytotoxic T lymphocyte (CTL) assay targeting EL4 tumor cells expressing NS3/4A protein (EL4-NS3/4A). We also treated mice bearing EL4-NS3/4A tumor with the combination therapy in vivo. The results confirmed that the combination therapy of B. longum 2165 and IFN-α induced significantly higher IFN-γ secretion, higher population of CD4+T and CD8+T cells secreting IFN-γ, and higher CTL activity against EL4-NS3/4A cells compared with the control groups of phosphate-buffered saline, B. longum 2165 alone, and IFN-α alone (p < 0.05). We also confirmed that the combination therapy strongly enhanced tumor growth inhibitory effects in vivo with no serious adverse effects (p < 0.05). These results suggest that the combination of B. longum 2165 and IFN-α could induce a strong cellular immunity specific to NS3 protein as a combination therapy augmenting the current SOC immunotherapy against chronic HCV infection.
Introduction
H
We previously constructed a genetically modified recombinant Bifidobacterium longum displaying HCV nonstructural protein 3 (NS3) peptides and successfully applied it as an oral vaccine against chronic hepatitis C (18). This oral vaccine, named B. longum 2165, was designed to express recombinant NS3 peptides on the bacterial cell surface by galacto-N-biose/lacto-N-biose I-binding protein (GLBP), which is the ATP-binding cassette (ABC) transporter of B. longum. We confirmed that multiple oral administrations induced strong systemic NS3-specific immune responses, such as Th1/Th2 responses and inductions of anti-NS3 IgA and IgG, in mice (18). The HCV-NS3 protein has serine protease activity and RNA helicase activity for viral replication, which is known to carry a variety of CD4+T and CD8+T cell epitopes to induce strong HCV-specific T cell responses (8). Importantly, strong NS3-specific T cell immune responses, especially mediated by cytotoxic T lymphocytes (CTLs), play important roles in viral clearance and recovery from acute hepatitis C infection (14). In addition, any effective therapeutic vaccine against chronic HCV infection must induce sufficient levels of cellular immune responses to clear HCV and infected cells from infected patients (13). Thus, NS3 could be an ideal target antigen for the development of an effective therapeutic vaccine.
In this study, we sought to further develop the B. longum 2165 vaccine and confirm its immunological features in vivo and to examine the functional NS3-specific cellular immunity induced by B. longum 2165 in vivo. To enhance its therapeutic efficacy, we attempted to combine IFN-α therapy as an adjuvant to B. longum 2165 oral vaccination in an animal model. IFN-α has various immunoregulatory functions and is conventionally used in the SOC combined with ribavirin for HCV treatment (2,3). IFN-α reportedly promotes proliferation of T cells and antigen presentation and maturation of antigen-presenting cells to induce robust cellular immunity (3,9).
Materials and Methods
Bacterial strains
B. longum 2165 and B. longum 2012 were generated as described previously (18). B. longum 2165 expressed the GLBP-NS3 fusion protein on the bacterial cell surface. The NS3 protein was derived from genotype 1b, MKC1a strain (GenBank accession no. BAA08120.1). The GLBP gene, which is a membrane protein in the ABC transporter on the B. longum cell wall (21), was derived from a B. longum JCM1217 strain and the C-terminal of GLBP was fused to the recombinant NS3 protein. B. longum 2012 expressed only GLBP and was used as a control. Figure 1A is a schematic drawing of the antigen surface displaying the B. longum 2165 system. The B. longum 2015 and 2012 strains were grown anaerobically in Gifu anaerobic agar broth (Nissui, Tokyo, Japan) overnight at 37°C.
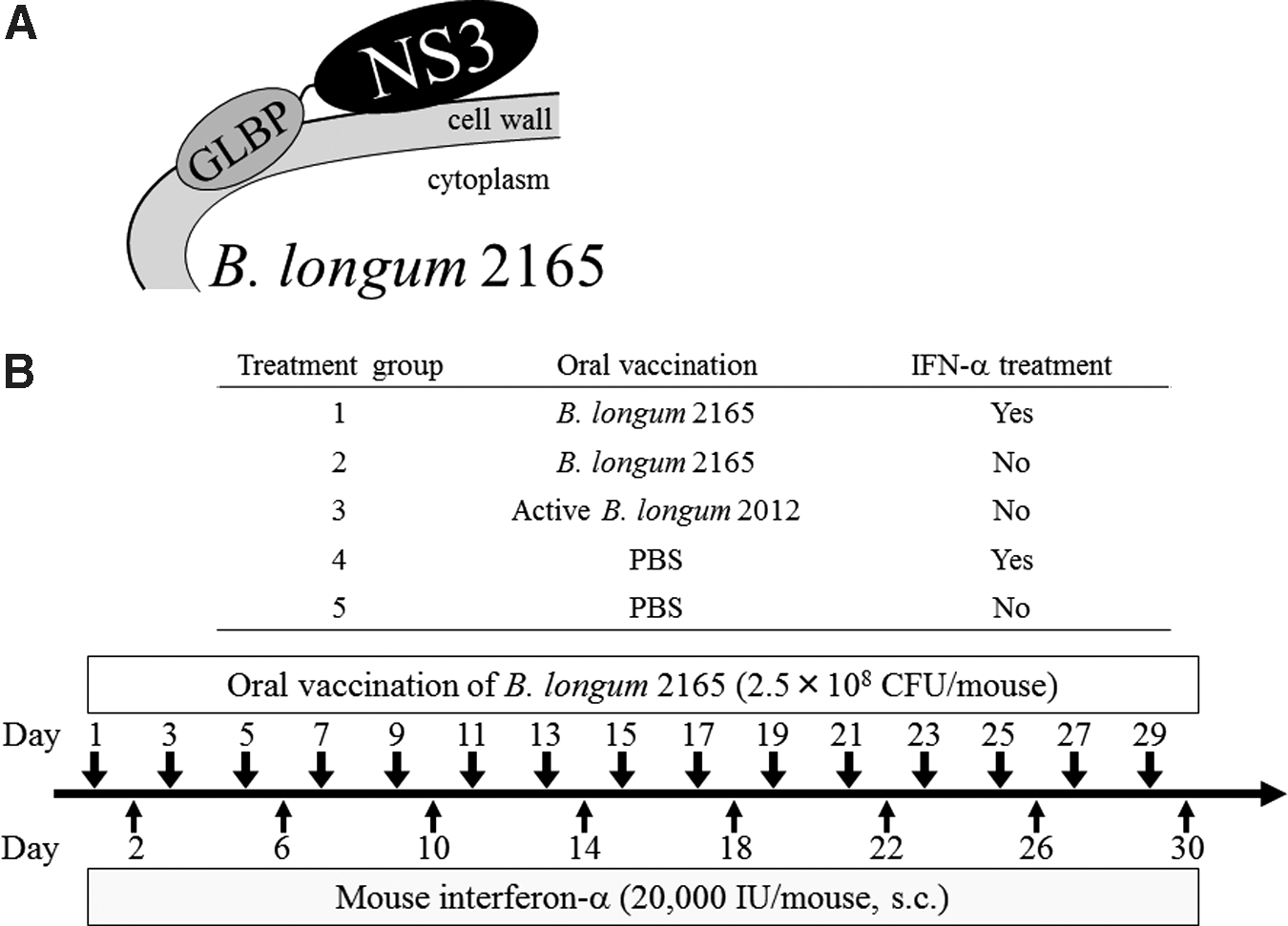
Schematic drawing of the antigen-displaying system of Bifidobacterium longum
Cells
The EL4 murine lymphoma cell line (H-2Db) was obtained from ATCC (Manassas, VA) and maintained in complete RPMI-1640 medium (6). EL4 cells stably expressing HCV-NS3/4A (EL4-NS3/4A) were generated by transduction of EL4 cells with the full-length NS3/4A complex derived from the HCV genotype 1b MKC1a strains inserted into pBApo-CMV Neo mammalian expression vector (Takara Bio, Otsu, Japan) (5,13). NS4A is a cofactor for NS3, and NS3 and NS4A coexpression essentially stabilizes its structure, serine protease activity, and stable expression in mammalian cells (8). EL4-Mock cells were also generated by transduction of EL4 cells with pBApo-CMV Neo vector. EL4-NS3/4A and EL4-Mock cells were maintained in complete RPMI-1640 medium supplemented with 800 μg/mL G-418 (Nacalai Tesque, Kyoto, Japan).
Immunization and sampling for evaluation of immune responses
To investigate the vaccine effect of B. longum 2165 in the IFN-α combination therapy, we performed an animal experiment according to the schedule shown in Figure 1B. Female C57BL/6N mice (H-2b), 6–8 weeks of age, were purchased from CLEA Japan (Tokyo, Japan) and maintained under specific pathogen-free environment. Thirty mice were randomly assigned to five oral vaccination groups, with 6 mice/group, as follows: group 1, B. longum 2165 with IFN-α; group 2, B. longum 2165; group 3, B. longum 2012; group 4, phosphate-buffered saline (PBS) with IFN-α control; and group 5, PBS control (Fig. 1A). The bacteria were attenuated by using heat and the expression of the fusion protein was corrected before administration. B. longum 2165 or B. longum 2012 (2.5 × 108 colony-forming unit/100 μL of PBS) or 100 μL of PBS was orally administered directly into the stomach using a feeding needle every other day (days 1–29, total 15 times), using a dose and period of immunization that were determined during a previous study (18). In addition to oral vaccination, group 1 and group 4 were subcutaneously (s.c.) injected with 20,000 IU mouse IFN-α (Miltenyi Biotec, Auburn, CA), twice weekly (days 2–30, total 8 times), using the dose and period of IFN-α treatment determined during a previous study and with regard to the current clinical regimen (12,20). Body weight was monitored weekly. On day 32 or 33, 3 mice per group on each day were euthanized under sevoflurane anesthesia and each spleen was aseptically resected. Isolated splenocytes were strained, hemolyzed, and suspended in complete RPMI-1640 medium for later in vitro assays. All aspects of the experimental design and procedure were reviewed and approved by the institutional ethics and animal welfare committees of the Kobe University Graduate School of Medicine.
Expression level of IFN-γ in splenocytes by enzyme-linked immunosorbent assay
An in vitro splenocyte stimulation procedure was performed as described previously (18). Briefly, isolated 4 × 105 splenocytes were plated into a 96-well microplate and stimulated with the GST-NS3 peptide (10 μg/mL) or the medium. GST-NS3 peptide was a kind gift from Dr. Hotta (13). Splenocyte cultures were incubated for 72 h at 37°C in 5% CO2; then, the supernatants were collected for enzyme-linked immunosorbent assay (ELISA) and frozen at −80°C until use. The concentration of IFN-γ in the culture supernatants was determined using a Mouse IFN gamma ELISA Kit (Thermo Fisher Scientific, Rockford, IL). The measurement procedure was performed according to the manufacturer's instructions.
Quantification of CD4+T and CD8+T cells producing IFN-γ by intracellular cytokine staining
To investigate the NS3-specific T cell immune response induced by the combination therapy, the frequency of CD4+T and CD8+T cells producing IFN-γ was measured by intracellular cytokine staining (ICCS). The procedure is described in a previous study (18). ICCS was performed using a BD Cytofix/Cytoperm™ Plus Fixation/Permeabilization Kit (BD Biosciences, San Diego, CA), according to the manufacturer's instructions. Briefly, 2 × 106 of splenocytes were restimulated with GST-NS3 peptide for 61 h. Then, GolgiStop was added and incubated for an additional 11 h. The splenocytes were collected and blocked with anti-mouse CD16/32 antibody (BioLegend, San Diego, CA) for 20 min on ice. After washing, the cells were stained with PerCP-conjugated anti-CD3, FITC-conjugated anti-CD4, and Alexa Fluor™ 647-conjugated anti-CD8a antibody (BD Biosciences) for 30 min on ice in the dark. The cells were washed twice and incubated with Fixation/Permeabilization Solution for 20 min on ice in the dark. After washing, the cells were stained with 5 μg/mL PE-conjugated anti-mouse IFN-γ antibody (BD Biosciences) for 30 min on ice in the dark. Then, the cells were washed and resuspended in the staining buffer. Fluorescence staining was assessed using FACSCalibur (BD Biosciences) and analyzed using CellQuest software.
Assay for NS3-specific CTL activity
To investigate functional CTL responses induced by the combination therapy, we performed a CTL assay using restimulated splenocytes. Splenocytes obtained from immunized mice were cocultured for 5 days with mitomycin C-treated EL4-NS3/4A cells and 20 ng/mL mouse IL-2 (Miltenyi Biotec) to generate effector cells (23). The viability of splenocytes was measured before the assay (data not shown). The effector splenocytes and target EL4-NS3/4A cells or EL4-Mock cells (5 × 103 cells) were cocultured in 96-well V-bottom plates for 4 h at 37°C in 5% CO2 at ratios of 20:1, 10:1, and 5:1. Specific CTL activity was measured using a lactate dehydrogenase Cytotoxicity Assay Kit (CytoTox 96 Nonradioactive Cytotoxicity Assay; Promega, Fitchburg, WI). The assay procedure was performed according to the manufacturer's instructions. The percentage of specific killing was calculated by the following formula: % specific killing = (experimental release—effector spontaneous release—target spontaneous release)/(target maximum release—target spontaneous release) × 100, according to the manufacturer's instructions.
Tumor growth inhibitory effect of vaccination with IFN-α on EL4-NS3/4A tumor
To investigate cellular immunity against the NS3-expressing tumor in vivo, we performed a tumor challenge with EL4-NS3/4A tumor cells, according to the methods described previously (6,23). On day 0, EL4-NS3/4A tumor cells (1 × 106 cells) were injected subcutaneously in the right flank of female C57BL/6N mice, 6–8 weeks of age. Following tumor injection, 30 tumor-injected mice were randomly assigned to six treatment groups and as described above (5 mice/group). The treatment procedure was performed as described above for 1–31 days and tumor development was monitored by measuring tumor size every fourth day, from day 7 to 35 after tumor injection. Tumor volume was expressed by the following formula: (longest diameter) × (shortest diameter)2 × 0.5.
Statistical analysis
Comparisons between multiple groups were performed using a one-way analysis of variance followed by the Tukey–Kramer method. Differences among means were considered significant when p < 0.05.
Results
The combination therapy did not affect the body weight change of the mice
To evaluate potential adverse effects of the combination therapy, the body weight of mice was measured. Throughout the immunization period, the mean body weights showed no significant differences among the different groups (Fig. 2). Furthermore, there were no obvious signs of toxicity, such as rough hair, hunched posture, or lethargy. Our findings suggested that the combination therapy of B. longum 2165 and IFN-α caused no serious side effects in these mice.
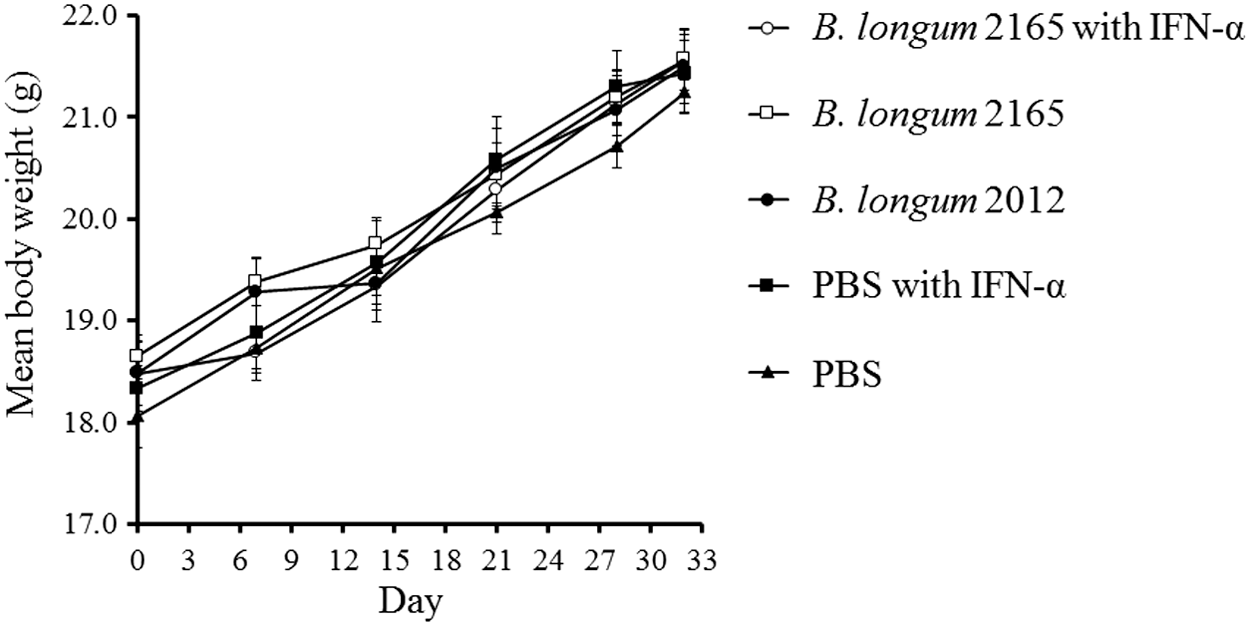
Body weight changes following combination therapy. Body weight of the mice (n = 6 per group) was measured on days 0, 7, 14, 21, 28, and 32. The mice were treated with B. longum 2165 and IFN-α (group 1), B. longum 2165 (group 2), B. longum 2012 (group 3), PBS with IFN-α (group 4), or PBS (group 5). Each data point represents the average of each group; bars ± SE. The mean body weight of each treatment group showed normal growth and there were no significant differences among the groups. SE, standard error.
The highest NS3-specific IFN-γ production in splenocytes was associated with the combination therapy
To determine the systemic cellular immune response after the vaccination, the levels of IFN-γ secretion in the splenocyte cultures were measured by ELISA. The results obtained showed that group 1 (B. longum 2165 with IFN-α) secreted significantly higher IFN-γ following pulsation of the GST-NS3 peptide compared with the other groups (p < 0.05) (Fig. 3). In addition, the IFN-γ secretion level of group 2 (B. longum 2165) was significantly higher than that in groups 3, 4, and 5 (p < 0.05).
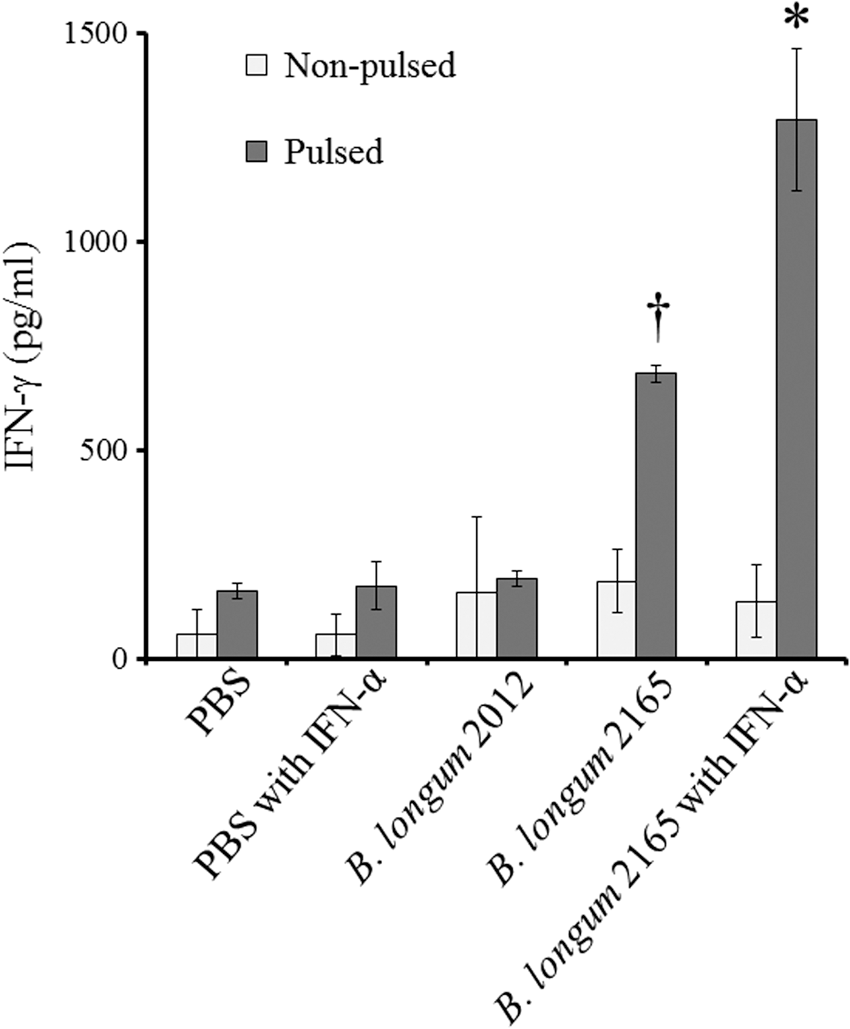
IFN-γ production in mouse splenocytes induced by the combination therapy. On day 32, following the treatment with B. longum 2165 with IFN-α, B. longum 2165, B. longum 2012, PBS with IFN-α, or PBS, splenocytes (n = 3 per group) were isolated and cultured with (pulsed) or without (nonpulsed) GST-NS3 peptide for 72 h in vitro. Each data point represents the average of each group; bars ± SE. Significantly higher IFN-γ secretion was detected from the pulsed splenocytes of the group, which received B. longum 2165 compared with the other groups († p < 0.05), and IFN-α treatment significantly enhanced the secretion (*<0.05).
The highest number of CD4+T and CD8+T cells producing IFN-γ was associated with the combination therapy by ICCS
We performed ICCS to investigate the frequency of CD4+T and CD8+T cells producing IFN-γ in splenocytes obtained from mice. T lymphocytes in splenocytes were gated by forward scatter/side scatter plot and as CD3-positive cells (Fig. 4A). Figure 4B shows the representative flow cytometric dot plots of each treatment group. As shown in Figure 4C, the frequency of CD4+T and CD8+T cells producing IFN-γ in group 1, which received B. longum 2165 and IFN-α, was significantly higher than that in the other groups (p < 0.05). In addition, the frequencies of both IFN-γ-producing CD4+T and CD8+T cells in the B. longum 2165 group were significantly higher than those in the other control groups 3, 4, and 5 (p < 0.05).
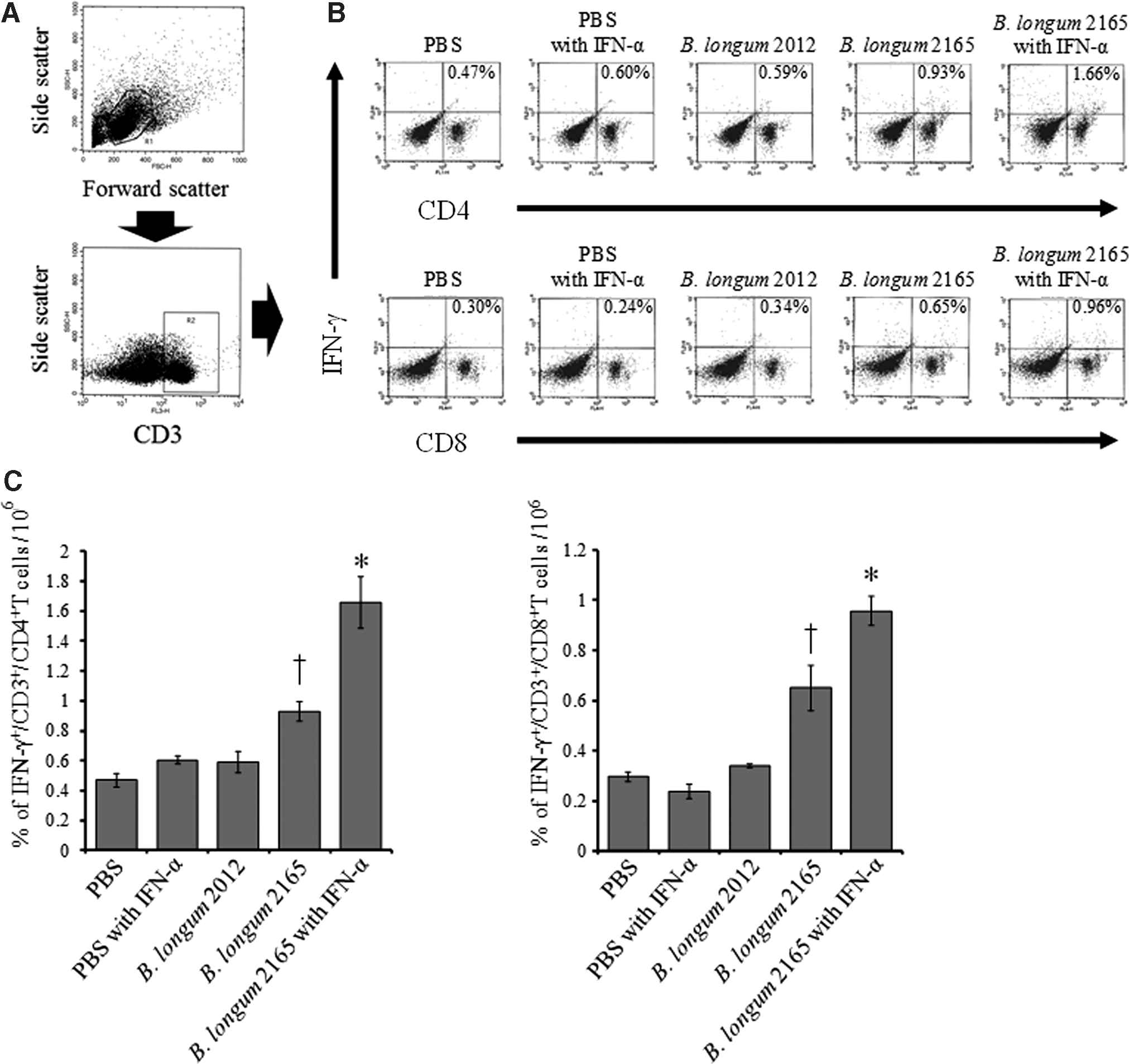
Intracellular cytokine staining for detection of IFN-γ-producing CD4+T and CD8+T cells. On day 32, following treatment with B. longum 2165 with IFN-α, B. longum 2165, B. longum 2012, PBS with IFN-α, or PBS, splenocytes (n = 3 per group) were restimulated with GST-NS3 peptide for 72 h. The frequency of IFN-γ-producing CD4+T and CD8+T cells was detected by flow cytometry. T lymphocytes were gated by forward scatter/side scatter plot and as CD3-positive cells
The highest NS3-specific CTL activity in vitro was observed in mice with combination therapy
NS3-specific CTL activity was measured by a CTL assay against EL4-NS3/4A tumor cells. Figure 5A shows that the NS3-specific CTL activities of each effector/target ratio were significantly stronger in group 1 receiving B. longum 2165 and IFN-α, compared with the other groups (p < 0.05). In addition, B. longum 2165 single therapy induced significantly higher NS3-specific lysis compared with other control groups (p < 0.05). In contrast, these CTL activities were background level against EL4-Mock cells (Fig. 5B). These results indicated that multiple oral administrations of B. longum 2165 induced functional NS3-specific CTL activity and that the IFN-α combination treatment could have enhanced therapeutic efficacy against NS3.
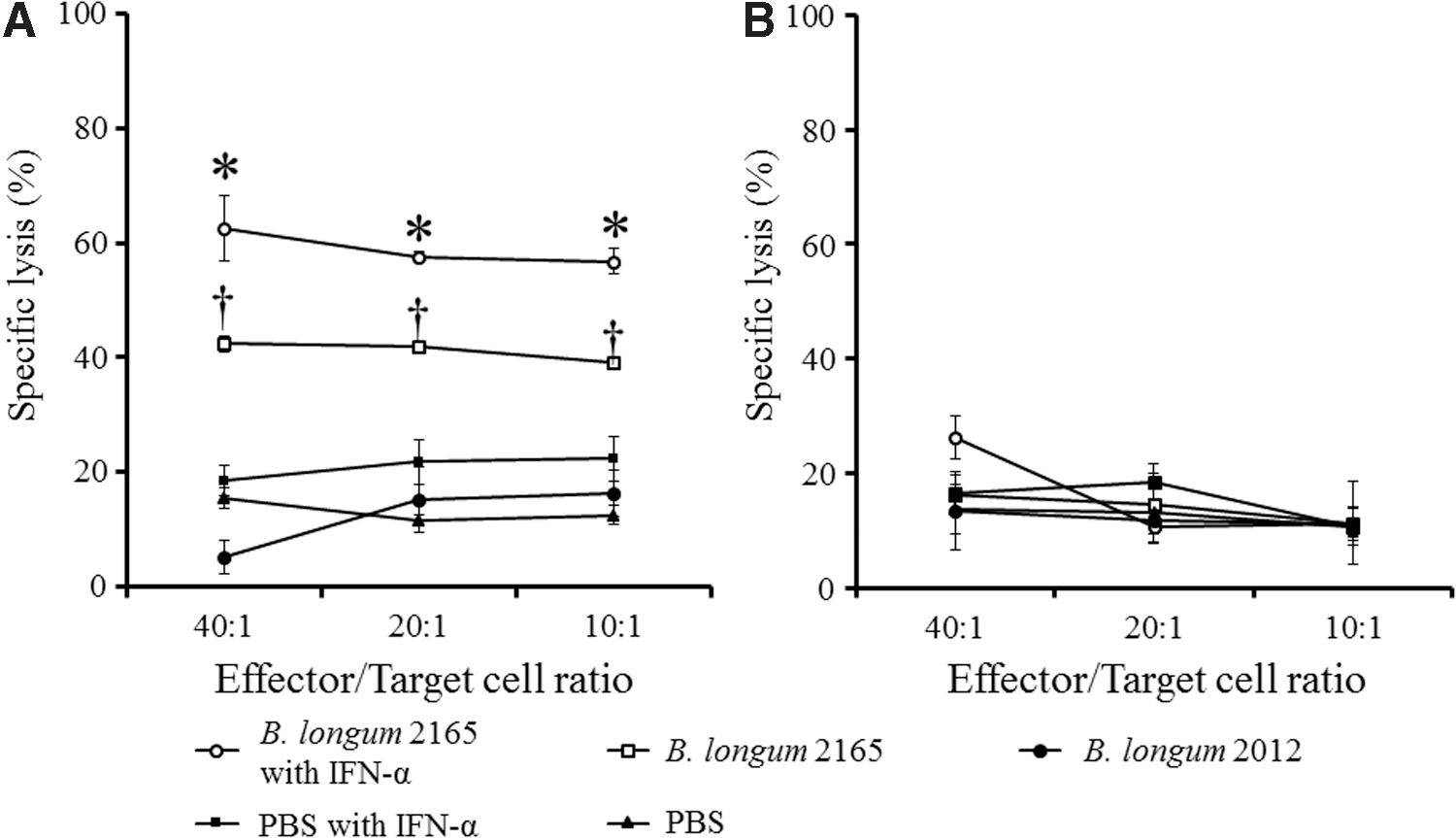
NS3-specific CTL activity induced by the combination therapy. On day 33, following treatment with B. longum 2165 with IFN-α, B. longum 2165, B. longum 2012, PBS with IFN-α, or PBS, splenocytes were isolated (n = 3 per group) and restimulated in vitro for 5 days with EL4-NS3/4A tumor cells and IL-2 to generate effector cells. The effectors and targets (EL4-NS3/4A or EL4-Mock) were cocultured for 4 h at ratios of 40:1, 20:1, and 10:1, respectively. The amount of released lactate dehydrogenase was measured and the percent of specific cell lysis is shown. Each data point represents the average of each group; bars ± SE. A significantly higher NS3-specific CTL activity against EL4-NS3/4A was detected in the group receiving B. longum 2165 compared with the other groups at each ratio (†
p < 0.05), and IFN-α significantly enhanced CTL activity (*p < 0.05)
The highest in vivo antitumor effect on EL4-NS3/4A tumors was associated with the combination therapy
To investigate the functional vaccine efficacy of combination therapy with B. longum 2165 and IFN-α, we measured the tumor growth of EL4-NS3/4A tumors in vivo. Figure 6 shows the tumor growth after inoculation. Tumor growth in group 1 was remarkably inhibited compared with the other groups throughout the therapeutic period. At day 35, after the inoculation, the mean tumor volume in group 1 receiving B. longum 2165 and IFN-α was significantly smaller than that in the other groups (p < 0.05). In group 2, where B. longum 2165 only was administered, the tumor volumes were significantly smaller than those in groups 3 and 5 (p < 0.05). The tumor volume in group 4 was slightly decreased compared with group 5 (not significant). These results indicated that combination therapy with B. longum 2165 and IFN-α could induce highly functional cellular immunity against HCV-NS3 and that B. longum 2165-alone therapy could also induce substantial immunity against HCV-NS3 in vivo.
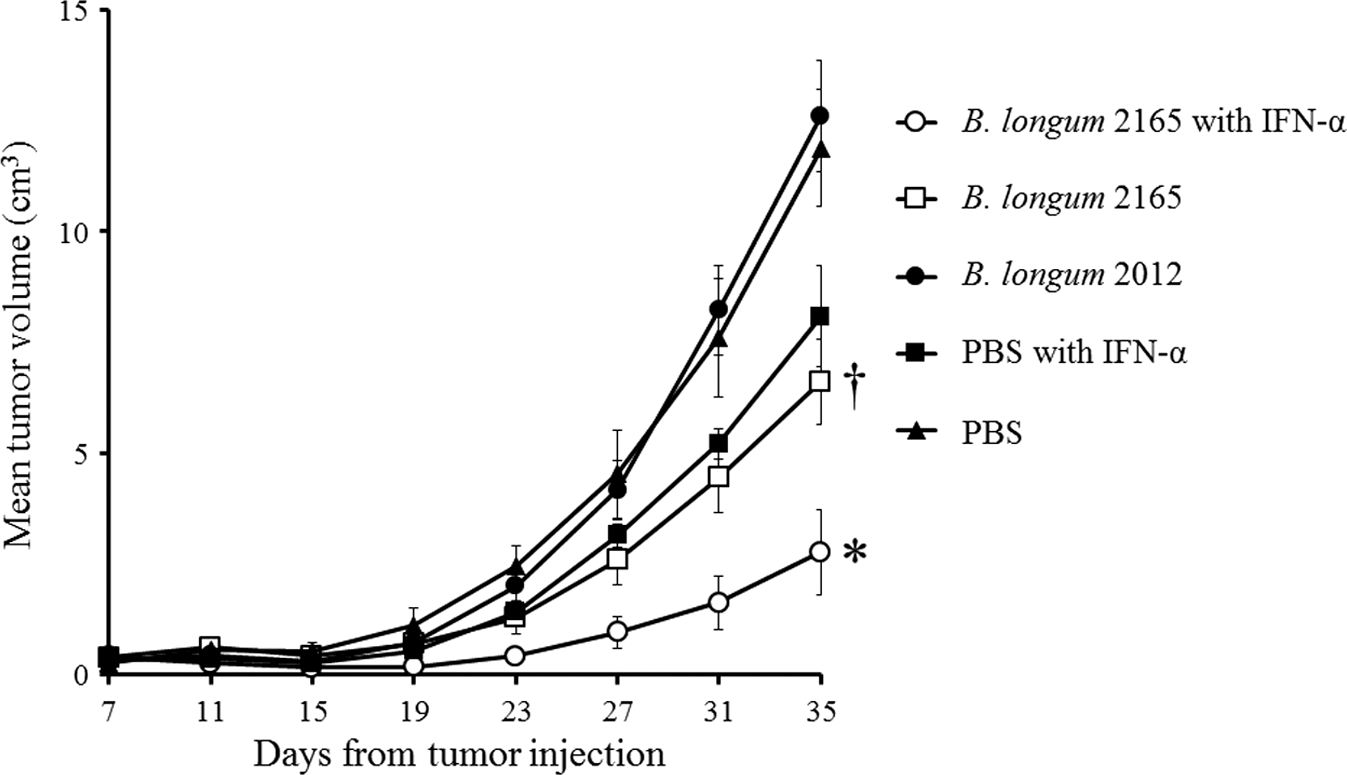
EL4-NS3/4A tumor growth curves. Mice (n = 5 per group) were s.c. injected with 1 × 106 EL4-NS3/4A on day 0, and received treatment with B. longum 2165 with IFN-α, B. longum 2165, B. longum 2012, PBS with IFN-α, or PBS, from day 1 to day 30. After the tumor cell injections, the tumor size was measured every fourth day and the volume was calculated using the following formula: (longest diameter) × (shortest diameter)2 × 0.5. Each data point represents the average tumor volume of each group; bars ± SE. Combination therapy with B. longum 2165 and IFN-α significantly inhibited tumor growth compared with the other treatments (*p < 0.05). In addition, B. longum 2165 single therapy significantly inhibited tumor growth compared with the other controls († p < 0.05).
Discussion
We previously developed a novel oral HCV vaccine using a recombinant Bifidobacterium displaying HCV-NS3 and demonstrated that this vaccine could induce efficient HCV-NS3-specific immunity in a mouse model (18). In this study, we investigated the feasibility of combining our oral HCV vaccine with IFN-α, which is conventionally used as a component of SOCs for the treatment of chronic HCV infection.
IFN-α is a multifunctional cytokine originally discovered as a direct antiviral protein and it has been used for SOC treatment for hepatitis C (2). In addition, IFN-α has various immunoregulatory functions, especially as it promotes the transition from innate to adaptive immunity. IFN-α upregulates IFN-γ production by both CD4+T and CD8+T cells, the proliferation and survival of CD8+T cells, and the proportion of CD4+ Th1 cell differentiation, through various mechanisms, including enhanced antigen presentation, costimulation, and cross-priming (1 –3). IFN-α also affects the maturation of dendritic cells (DCs) by upregulating the expression of MHC class I and important T cell costimulatory molecules (9,10). These findings suggest that IFN-α enhances adaptive T cell-mediated immunity and thus can be considered a good candidate as an adjuvant for vaccines against tumors and infectious diseases (7,16,17). In this study, we demonstrated a synergistic effect of a combination therapy using IFN-α and our B. longum 2165 oral HCV vaccine. In addition, the combination therapy showed no serious adverse effects in mice. These findings suggest that our combination therapy is probably as safe as treatment with IFN-α alone.
In regard to the vaccine's efficacy, we demonstrated the induction of strong NS3-specific cellular immune responses to the combination therapy. We observed the enhanced production of IFN-γ, considered as Th1 cytokines (Fig. 3). ICCS also showed that both CD4+T and CD8+T cells were activated by the combination therapy (Fig. 4). These results indicated that GLBP-NS3 fusion protein could efficiently stimulate T cell-mediated immunity, and additional IFN-α administration enhanced the immunological efficiency of B. longum 2165.
NS3-specific CTL responses reportedly play crucial roles in viral clearance and recovery from HCV infection (14). Thus, induction of functional NS3-specific CTL responses by the vaccine would be crucial for effectiveness against chronic HCV. In this study, we showed that strong NS3-specific CTL activities were induced by the oral administration of B. longum 2165 and were further enhanced by combination therapy with IFN-α in in vitro CTL assays targeting EL4-NS3/4A tumor cells (Fig. 5).
Subsequently, we showed that a single B. longum 2165 vaccination significantly inhibited EL4-NS3/4A growth in vivo and that IFN-α combination therapy could significantly enhance therapeutic efficacy (Fig. 6). PBS with IFN-α therapy slightly inhibited tumor growth; however, this inhibition probably reflected nonspecific innate responses.
B. longum 2165 is designed to induce adaptive NS3-specific systemic immune responses through the intestinal mucosal immune system. In general, orally administered antigens are recognized by microfold cells on the Payer's patches that belong to mucosa-associated lymphoid tissue in the intestine, followed by antigen presenting by DCs to lymphocytes such as B cells and T cells (11). In this study, IFN-α could promote the maturation of DCs and activate T cells, resulting in IFN-γ secretion of CD4+T and CD8+T cells, and the proliferation and survival of CTLs. Then, activated antigen-specific CTLs could elicit antitumor/antiviral effector activities (1,10,17). As a result of these immunological interactions, we consider that IFN-α could enhance the NS3-specific adaptive cellular immunity induced by this oral vaccine and elicit synergistic antitumor/antiviral effects in a combination therapy with B. longum 2165. IFN-α is the most commonly used agent in the SOC for chronic HCV infection. Thus, our findings suggest that a B. longum 2165 oral vaccination could be a good candidate to enhance the SOC as an additive agent for recovery from chronic hepatitis C and to prevent progression to liver cancer.
Recently, several clinical and preclinical studies of HCV therapeutic vaccines using peptides, DNA, viral vectors, yeasts, or DCs (8,24) showed substantial decline of HCV serum RNA level and antigen-specific immune responses especially mediated by HCV-specific CTL activity (8). In this study, B. longum 2165 also induced strong HCV-NS3-specific CTL activity in mice. The findings in the previous studies support the use of our oral vaccine as a candidate therapeutic vaccine for chronic HCV treatment. In addition, we would like to emphasize the advantages of the oral formula of our vaccine as a less invasive approach. As previously reported, other HCV therapeutic vaccines are administered by parenteral routes using needles, while B. longum 2165 can be administered orally. This allows a safer and more cost-effective approach for large numbers of chronic HCV patients.
Although new DAAs, including sofosbuvir, have achieved a greater than 90% SVR rate in chronic HCV infections without IFN-α, the long-term recurrence and cancer incidence outcomes of these DAA-based treatments without IFN-α have yet to be determined (22). It is still commonly considered that induction of cellular immunity against HCV is important for the best long-term outcomes (3,21). In addition, DAAs are currently very expensive and not affordable for many patients, especially in developing countries. When HCV infection is diagnosed, IFN-α treatment is usually the principle SOC protocol (24). Our oral vaccine can be easily added to IFN-α treatment to enhance its therapeutic effects. In this study, we demonstrated the potential of B. longum 2165 as a novel and effective therapeutic oral HCV vaccine. Bifidobacteria, the basis of this oral vaccine, is safe, easy to amplify and refine, and can be carried and stored in cold chain-free conditions. The B. longum 2165 HCV oral vaccine represents a potentially important new paradigm in HCV treatment, especially given its cost-effectiveness and low toxicity.
Footnotes
Acknowledgments
This study was supported, in part, by a grant-in-aid for Scientific Research (No. 23591475) from the Japan Society for the Promotion of Science; a grant-in-aid through the Program of Japan Initiative for Global Research Network on Infectious Diseases from the Ministry of Education, Culture, Sports, Science and Technology; and a grant for Science and Technology Research Partnerships for Sustainable Development from the Japan Science and Technology Agency, the Japan International Cooperation Agency, and MSD K.K., Japan.
Author Disclosure Statement
No competing financial interests exist.