
Abstract
Synthetic peptides are one of the hepatitis C virus (HCV)-specific small molecules that have antiviral activity and represent a target for HCV vaccine. This study aims to determine the lowest concentration of adjuvanted and non-adjuvanted (multiple antigenic peptide [MAP]) form of three conserved HCV envelope peptides that can induce murine immunogenic responses and evaluate the neutralization capacities of the generated antibodies (Abs) against HCV in cultured Huh7.5 cells. In this study, three HCV synthetic peptides, E1 peptide (a.a 315–323) and E2 peptides (a.a 412–419 and a.a 516–531) were synthesized. Female Balb/c mice were immunized with different concentration of either adjuvanted linear peptides or nonadjuvanted MAP peptides to determine the lowest dose that generates Ab responses enough to confer viral neutralization in vitro. The humoral responses targeting these peptides in immunized mice sera were measured by enzyme-linked immunosorbent assay (ELISA). Viral neutralization capacities of the generated mice Abs were assessed using Huh7.5 cells infected with the HCVcc infectious system (J6/JFH-1). The results of this study showed that the MAPs induce higher Ab titers than adjuvanted linear peptides after 4 weeks of immunization (p = 0.003). The viral neutralization experiments showed that the immunized mice sera contain anti E1/E2 Abs that blocked HCVcc (J6/JFH-1) entry into Huh7.5 cells. In conclusion, the three HCV envelope MAP peptides are more immunogenic and produce higher neutralizing Abs than linear peptides; therefore, they can be essential components for HCV vaccine.
Introduction
T
The treatment of hepatitis C infection has been greatly improved during the recent years. There are many HCV-specific small molecules with an antiviral activity that have been reported, such as antisense oligodeoxynucleotides (9,27), synthetic peptides (12,13,25), miRNA interference, and direct-acting antivirals, which targeted specific HCV nonstructural proteins (4,40,41). The recently introduced HCV/NS3-protease inhibitors (boceprevir and telaprevir) increased the rate of sustained viral response, but their use is still limited, and is associated with drug resistance (53). The costs for the treatment of hepatitis C infection and disease complications, as well as the need for liver transplantation are immense (28,32). So the need of new therapies to prevent and/or treat HCV infection is essential. Recently, the multiple antigenic peptides (MAPs), which are the superior designs of synthetic peptides, are used in developing HCV vaccine models (21). However, the research is still ongoing to find an effective and specific vaccine for HCV infection (30).
Several findings showed the importance of humoral immune response, as demonstrated early that the administration of polyclonal HCV Ig antibodies (Abs) prevents sexual transmission (38). Moreover, it has been found that HCV viral clearance was associated with a rapid induction of neutralizing Abs (nAbs) in the acute phase of HCV infection. In contrast, the Abs from patients progressing to chronic infection displayed a delayed induction or a complete absence and thus fail to neutralize the virus in the early infection (5,37). Also, it was demonstrated that humanized anti-E2 Abs have the potential to prevent HCV recurrence in liver transplant patients, while escape from nAbs influences HCV recurrence in the newly transplanted liver graft (14,15). Therefore, these studies proved that the rapid induction, quality, and quantity of the nAbs during the acute phase of infection are very essential to control HCV infection and progression.
The HCV envelope glycoproteins (E1, E2) are the main targets of protective Ab response. Recently, different synthetic peptides were shown to generate specific nAbs against HCV infections (6,12,18,25,35,57). Other studies showed that Abs directed to epitopes present within the E2 glycoprotein hypervariable region 1 (HVR1) and downstream of HVR1 are neutralizing (6,7,8,12,57). It was reported that Abs generated against HVR1 synthetic peptides prevented HCV viral infection in permissive cell cultures (50) and protected chimpanzees from infection with HCV (18), while others were presumed to be genotype specific and could not confer global protection (16,23,42). Also, few HCV epitopes proved to be immunogenic. Therefore, identifying epitopes on the HCV E1 and E2 glycoprotein that can elicit nAb responses is very imperative.
It was believed that nAbs targeting conserved regions of the HCV viral envelope glycoproteins were able to neutralize the different viral genotypes, subtypes, and quasi-species present within an infected individual. Previously, we demonstrated that conserved peptides derived from HCV E1 and E2 envelope proteins produced murine and caprine humoral and cellular responses. Moreover, the generated Abs exhibited viral neutralization in permissive cell cultures (8,9,12). In this study, we define the effective murine immunogenic concentrations of three conserved envelope epitopes and explore their viral neutralization capacities in cultured Huh7.5 cells. Also, we aim to examine the immunogenic potential of adjuvant-free regimen by using 8 arms MAP formula of synthetic peptides.
Materials and Methods
Design and synthesis of HCV E1/E2 peptides
Peptide sequences were selected using CLUSTAL W multiple sequence alignments with different HCV isolates derived from HCV (1a, 1b, 2a, 2b, 3a, 4a, 5a, and 6a) (
Immunization of mice
Two methods of immunization were followed: using adjuvanted linear peptides and nonadjuvanted MAPs to determine the lowest dose that triggers Ab responses enough to confer viral neutralization in vitro. Female Balb/c mice were purchased from Theodor Bilharz Research Institute (Giza, Egypt).
Immunization of mice with linear adjuvanted peptides
Mice were maintained and studied according to local guidelines for animal care. Alum phosphate was used as an adjuvant with the synthetic peptides and was prepared according to Ref. (20). Animals were divided into five groups. Each group consists of 15 mice (6–8 weeks old; 25 gm body weight) and received subcutaneous timely doses of 50, 100, 200, 400, and 800 ng of peptide mix of the three peptides in a ratio of 1:1:1, coupled with Alum phosphate (Al PO4), on days 0, 28, and 56. Sera from mice were collected on days 27, 55, and 62. As a control, the serum was collected from each individual mouse the day before the first injection.
Immunization of mice with nonadjuvanted MAPs
Mice were divided into three groups. Each group consists of 15 mice (6–8 weeks old; 25 gm body weight) and received subcutaneous timely doses of 800, 2,400, and 8,000 ng of MAP mix in a ratio of 1:1:1 on days 0, 28, and 56. Sera from mice were collected on days 28, 56, and 85. As a control, sera from individual mouse were withdrawn 1 day before the first injection.
Levels of Ab responses toward E1-/E2-derived peptides
Specific anti-E1/E2 Ab titers targeting peptide mix were determined by an in-house enzyme-linked immunosorbent assay (ELISA) as described previously by El-Awady et al. (11). The assay was carried out in U-shaped polystyrene microtiter ELISA plates (ALTO) that were initially coated with 100 μL/well of the three linear peptide mix conjugated with Keyhole Limpet Hemocyanin (KLH) at a concentration of 5 ng peptide-KLH/mL of carbonate/bicarbonate buffer (pH 9.6). The plates were incubated overnight at room temperature and were washed thrice by 0.05% (v/v) phosphate-buffered saline (PBS)-T20 (pH 7.2). Free active sites were blocked using 0.5% (w/v) bovine serum albumin in a carbonate/bicarbonate buffer for 2 h at 37°C. After washing thrice, 100 μL/well from each mouse serum that was prediluted 1: 50 in PBS was added as a primary Ab and incubated at 37°C for 2 h. Plates were washed thrice and then 100 μL/well of anti-mouse IgG peroxidase conjugate (KPL; Gaithersburg) diluted 1:1,000 in 0.5% (w/v) bovine serum albumin in the carbonate/bicarbonate buffer was added and the mixture was incubated at 37°C for 1 h. Then, 100 μL/well orthophenylenediamine substrate (Sigma) was added and plates were left for 10 min at room temperature till color development. The enzymatic reaction was stopped using 40 μL/well 2 N HCl and the optical density was read at 490 nm using a multiwell plate reader (TECAN; SUNRISE).
Viral neutralization by mouse anti-HCV E1/E2 peptides polyclonal Abs
Hepatoma Huh7.5 cells were cultured and infected with HCVcc (J6/JFH-1) infectious system as described in Ref. (49). The J6/JFH-1 was a generous gift from Charles Rice (The Rockefeller University, USA, MTA# 642 to Dr. Mostafa K. El-Awady 2007). Viral neutralization experiment was done as described previously by Susser S et al. (52) with some modifications. In brief, Huh7.5 cells were cultured in 24-well plates (32,000 cells per well) in a complete Dulbecco's Modified Eagle's Medium and were incubated in a CO2 incubator at 37°C and 5% CO2 overnight. Then, media obtained from infected Huh7.5 cells with HCVcc (J6/JFH-1) infectious system were used as a source of virus. For the neutralization assay, sera of immunized and control mice (diluted at 1:20 in sterile PBS to a final volume of 250 μL) were heat inactivated at 56°C for 1 h, mixed with equal volume of HCVcc (J6/JFH-1) in 10% fetal bovine serum (FBS)/DMEM, and incubated at 37°C for 1 h. HCVcc (J6/JFH-1) infectious system incubated with Abs containing serum was used to infect Huh7.5 cells seeded in 24-well plates and then incubated at 37°C for 3 h. The virus-containing media were removed, washed with sterile PBS and replaced with 15% FBS DMEM, and incubated at 37°C. After 48 h, media were removed and cells were washed twice with sterile PBS. RNA was extracted from cells of each well using acid guanidinium thiocyanate-phenol-chloroform extraction method described by Chomczynski P et al. (3). The RNA was reverse transcribed and amplified to detect the presence of HCV RNA as described by El-Awady et al. (10) and modified by El-Awady et al. (12).
Results
Humoral response against adjuvanted linear peptide mix
Five groups (15 mice each) received timely doses of 50, 100, 200, 400, and 800 ng of Al PO4-adjuvanted linear peptide mixes on days 0, 28, and 56. Blood samples were collected from the retro-orbital plexus/sinus of immunized mice 1 day before immunization and screened by ELISA for Ab response against the conserved E1/E2 peptides. In general, Abs raised against adjuvanted E1/E2 peptides were detected in mice sera and their titers increased by both time and dose. The Abs level in each group showed a statically significant difference (p < 0.001). The results shown in Figure 1 and Table 1 showed that the generated antibodies were specific. The results indicated a good specificity of the generated antibodies.
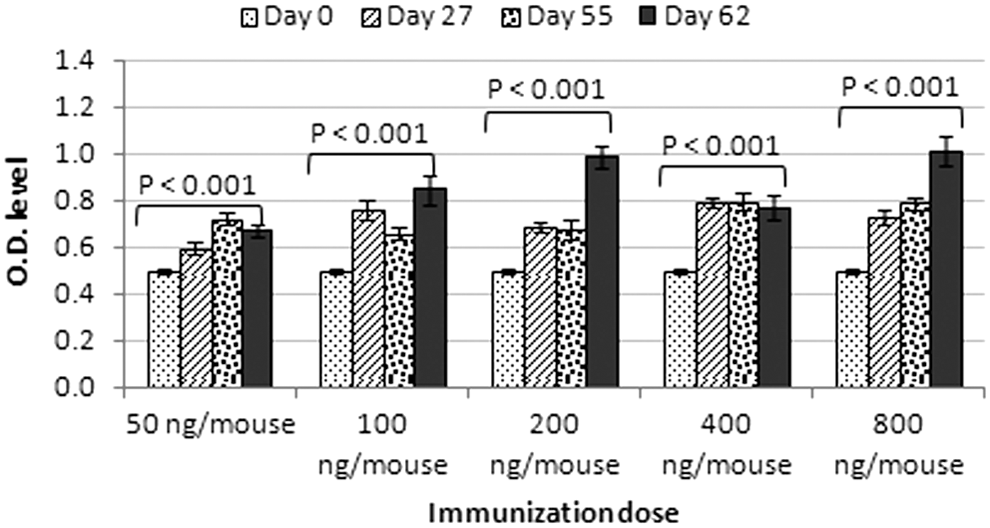
Humoral responses to Al PO4/linear E1/E2 peptides. Five groups (15 mice each) received timely doses (50, 100, 200, 400, and 800 ng) of adjuvanted peptide mix on days 0, 28, and 56. Ab responses were measured in sera of mice on days 0, 27, 55, and 62. Ab titers increased by time and dose. The doses 200 and 800 ng/mouse were the most immunogenic at the end of the injection protocol. Bars represent mean ± SE of Ab titers. The Abs level in each group showed a statistically significant difference (p < 0.001). Abs, antibodies; SE, standard error; O.D., optical density.
Significant difference between days 27, 55, or 63 and day 0 in each immunization dose at significant level (0.01 level).
OD, optical density; SE, standard error.
Humoral immune response against MAPs
Three groups (15 mice each) received timely doses of 800, 2,400, and 8,000 ng of MAP mix on days 0, 28, and 56. Blood samples were collected from the retro-orbital plexus/sinus of immunized mice the day before immunization and screened by ELISA for Ab response against the conserved E1/E2 peptides. Ab titers were elevated by time, but not by doses of MAP peptides, which indicated that humoral response to MAP is specific in mice as shown in Figure 2 and Table 2.
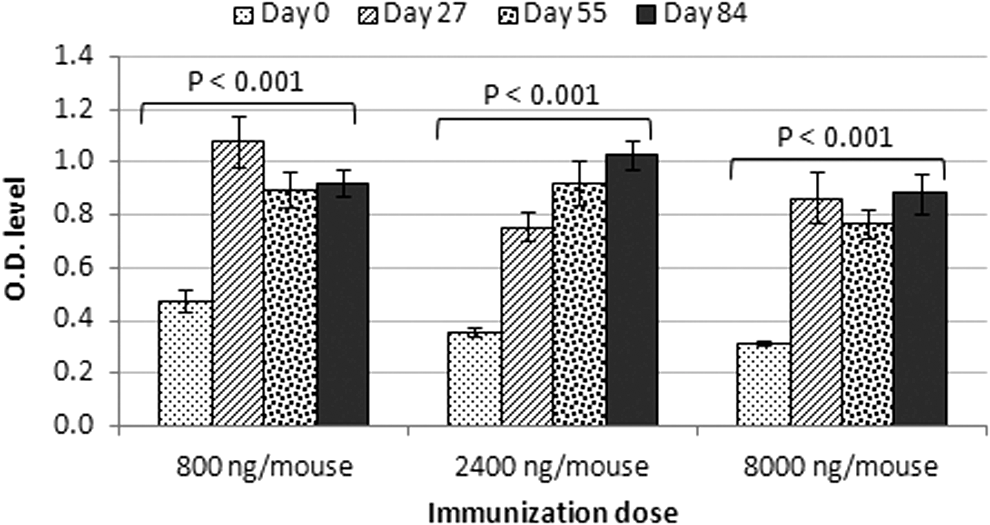
Humoral response of the MAP mix. Three groups (15 mice each) received timely doses of MAP mix on days 0, 28, and 56. Specific Abs against the MAP mix were detected in the sera of mice immunized with doses 800, 2,400, and 8,000 ng on days 27, 55, and 84. Bars represent mean ± SE of Ab titers. The Abs level in 800, 2,400, and 8,000 ng groups showed statistically significant differences (p-values were <0.001). MAP, multiple antigenic peptide.
Significant difference between days 27, 55, or 84 and day 0 in each immunization dose at significant level (0.01 level).
MAP, multiple antigenic peptide.
Comparison between humoral responses to Al PO4-adjuvanted linear and MAP-nonadjuvanted peptides in mice
The humoral response against 800 ng linear adjuvanted and MAP was compared and the results showed that the immune response against MAP was higher than the adjuvanted peptides as shown in Figure 3 and Table 3. These results suggest that MAP induce higher Ab titers than Al PO4/linear peptides at a dose of 800 ng/mouse after 4 weeks of immunization with a statistically significant difference (p < 0.01).
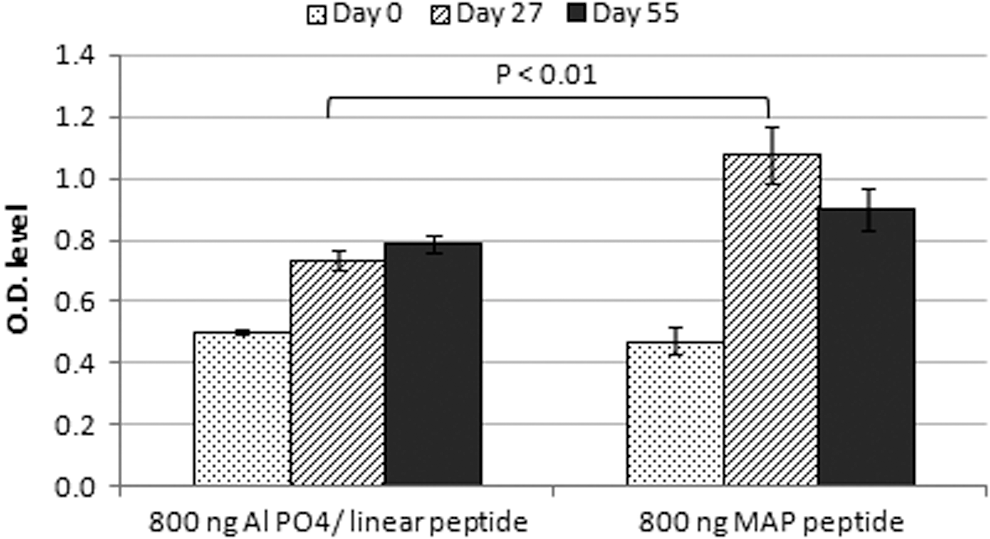
Comparison between humoral response to 800 ng Al PO4/linear and 800 ng MAP: Balb/c mice were injected with either Al PO4/linear peptide mix or MAP mix (15 mice each) and Ab titers were measured on days 27 and 55. Bars represent mean ± SE of Ab titers. The immune response against MAP was higher than the adjuvanted peptides (p < 0.01).
Individualized humoral response of each mouse to 800 ng MAP
Specific anti MAP Abs were detected in the tested mice, except for mouse #4 who failed to elicit humoral response as shown in (Fig. 4). In the remaining eight mice, the Ab response peaked at either 27 or 56 days with slight decline on day 84 (in six out of eight animals).
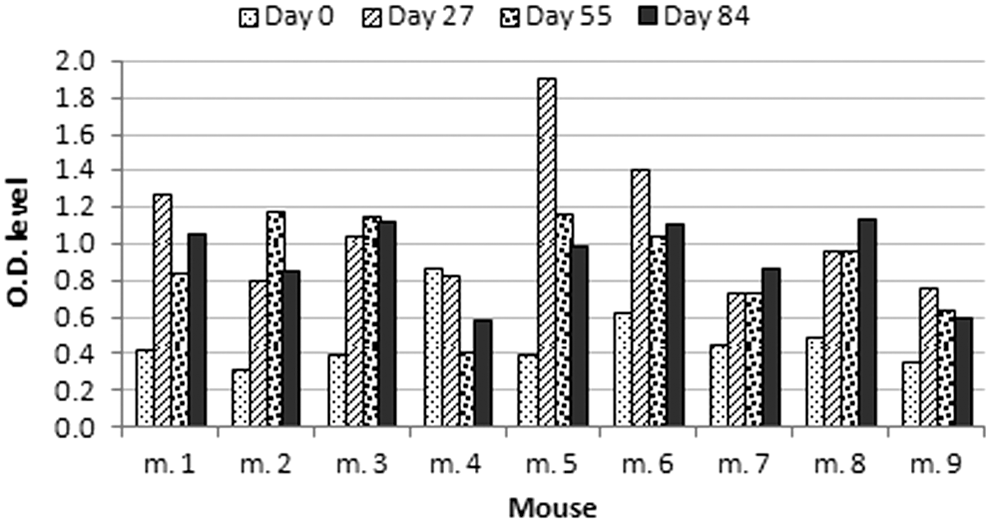
Humoral responses of individual mice to 800 ng MAP mix: Balb/c mice were marked by ear poring, then injected by 800 ng of MAP mix on days 0, 28, and 56. The Ab response was measured on days 0, 27, 55, and 84. Bars represent the Ab levels for each animal at different time points.
Viral neutralization by anti-mouse anti E1/E2 Abs
Sera of immunized and nonimmunized mice were heat inactivated and incubated with HCVcc (J6/JFH-1 replicon) and the mix was used in the infection of Huh7.5 cells as described in Materials and Methods section. RNA was extracted from infected cells and the intracellular load of HCV RNA was detected by nested RT-PCR. Eight serum samples were used in this experiment: three from nonimmunized mice and five from mice immunized with the mix of the three MAP described in this study and derived from E1/E2 proteins. Cells infected with sera from nonimmunized mice+HCVcc (J6/JFH-1) (n = 3) showed successful HCV infection, whereas four out of five cultures that were infected with HCVcc (J6/JFH-1), which was preincubated with immunized mice sera showed complete inhibition of HCV entry as shown in Figure 5. These results indicated that the immunized mice sera contain anti E1/E2 Abs that block HCVcc (J6/JFH-1) entry into Huh7.5 cells, that is, nAbs.
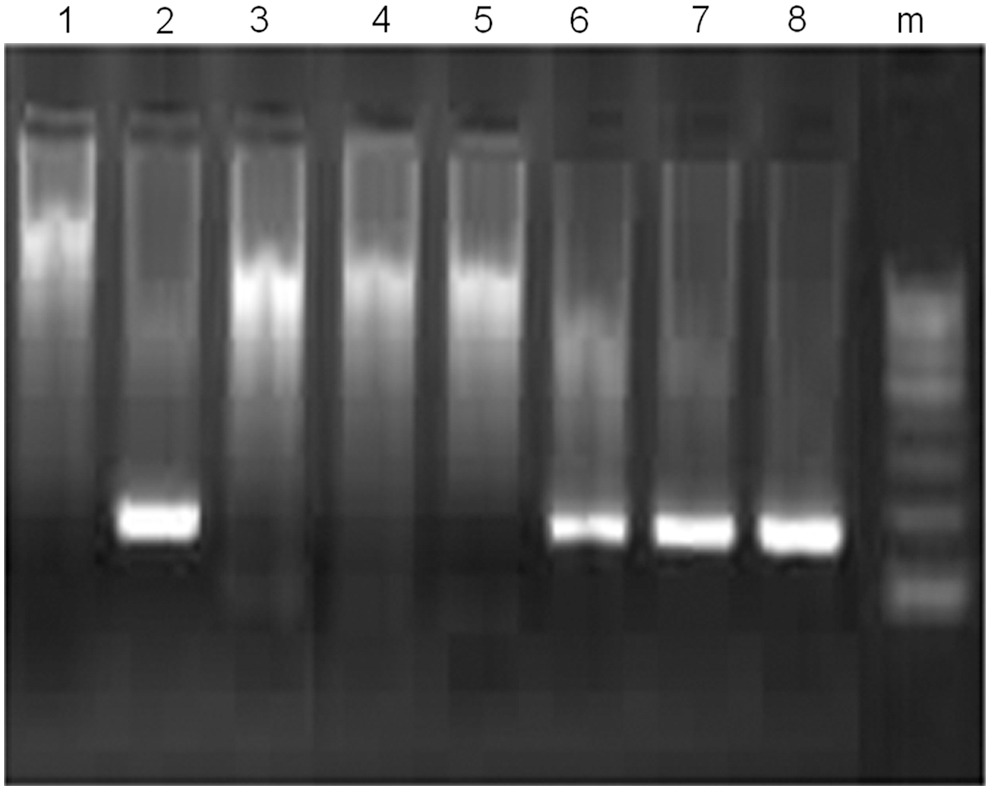
Effect of anti-E1/E2 Abs on HCVcc infection into Huh7.5 cells: HCVcc (J6/JFH-1) infectious system was incubated with sera from either immunized mice or preimmunized mice as immunization control and the mix was used to infect Huh7.5 cells. Intracellular RNA was extracted from infected cells and HCV RNA was RT-PCR amplified. Lanes 1, 2, 3, 4, 5 are viral RNA amplification products in presence of five highly immunized mice sera; lanes 6, 7, 8 are PCR products after incubation with preimmunized mice sera. Lane M is φx 174-DNA Hae III digest molecular weight marker. HCV, hepatitis C virus.
Discussion
The infection with HCV is a major global health problem with serious sequelae, including liver cirrhosis, hepatic failure, and hepatocellular carcinoma, and it is the leading cause for liver transplantation. The annual rate of infection is as high as 150,000 new cases each year in Egypt. Currently, there is no prophylactic or therapeutic HCV vaccine. However, there are several evidences supporting the feasibility of such approaches (29,30,58). In HCV infection, the role of Abs in controlling or preventing the infection remains controversial, unlike hepatitis B where immune protection is associated with strong Ab response against the envelope proteins. However, the fact that 15–45% of HCV-infected individuals are able to spontaneously clear their infection indicates the importance of the adaptive immune response in clearance of the virus. Studies by Edwards VC et al. and Wang Y et al. (7,57) proved the nAbs' role in controlling HCV infection.
The main problem with the development of immunogenic products against HCV infection is the genetic diversity among different viral strains, and only some HCV epitopes confirmed to be immunogenic (26,31,42,51). Therefore, the major challenge is to select conserved epitopes with immunogenic characteristics on the HCV E1 and E2 glycoprotein that can generate Abs able to neutralize different HCV genotypes and provide universal protection. Studies showed that linear peptides have poor immune response and low reactivity, while the synthesis of bulk synthetic peptides in the form of MAP leads to chemical stability and high specificity and reactivity (24,43,44). Also, several studies suggested the successful use of MAP peptides and anti MAP Abs as safe, specific, and efficient for viral antigen detection and vaccine development (1,22,23,44).
In a previous study from our laboratory (12), three MAP on the bases of sequence homology among genotype 4 subtypes were designed. All the three peptides were able to react with the corresponding Abs in almost ∼100% of sera from chronically infected HCV genotype 4 patients.
In this study, mice were immunized with escalating doses of E1/E2 peptide mix using two formulations: linear peptides with aluminum phosphate as an adjuvant or the peptide mix in an 8 arm MAP adjuvant-free form to determine humoral responses at different time points postinjection. Specific anti E1/E2 Ab titers continued to elevate by time and by doses up to 800 ng/mouse concentration. Current ELISA results support previous data from our laboratory that selected MAP mix-induced murine polyclonal Abs that were able to react independently with each individual peptide either in a linear form or as part of circulating viral particles in chronic HCV patients (8,12). Also, these results showed that nonadjuvanted MAP induce higher immune response than adjuvanted peptides (p < 0.01). These results are in agreement with findings by Sadler K et al. (45) who reported that the immunization with MAP form produced pure and specific Abs, which reacted with the cognate protein, while antisera produced from linear peptide conjugated to adjuvants or carrier proteins have interference Abs. Also, Saravanan et al. (46) reported that Abs produced against MAP from different infectious bursal disease virus epitope segments have high efficiency to detect the virus than linear peptides. Moreover, the generated murine Abs showed unprecedented capacity to interfere with HCVcc infectious system into hepatoma cells. Recently, two MAP designs containing HCV core, NS4B and NS5A epitopes, elicited cellular immunogenicity and inducted INFγ and IL2 responses (21).
Although studies on E1-specific nAbs are limited, there is evidence that E1-specific Abs were raised after experimental vaccination and these Abs were protective (33,43). Moreover, El-Awady et al. (11) previously showed that murine Abs to peptide spanning a.a (315–323) of genotype 4a blocked HCV entry in vitro. These results were later confirmed by Meunier JC et al. (33) who reported that E1 linear epitope encompassing a.a 313–327 generated two Abs that neutralize HCVpp bearing E1/E2 derived from different HCV genotypes. Also, a study by Verstrepen BE et al. (56) reported that the protective effect of the E1-based vaccine was better than that induced by an E2-based vaccine in chimpanzees infected with HCV genotype 1b. Mouse monoclonal Abs against the E1 peptide described herein blocked 69% of Huh7 cell cultures infected with HCV genotype 4a-positive sera from different patients (54).
Early data from our laboratory showed that Abs generated against HCV E2 (a.a 412–419) were neutralizing (12). These results supported the early findings of Owsianka A et al. and Tarr AW et al. (34,55) who reported that a linear epitope in the E2 region a.a 412–423 generated the first mouse monoclonal Ab AP33. The current neutralization results of Abs against the three peptides described herein support the previous viral neutralization data by our laboratory and others generated against individual peptides. (2,8,19,31,33,34,36,43,55,56). Caprine anti a.a 412–423 and a.a 517–531 Abs were neutralizing to HCV genotype 4a at 87.5% and 75%, respectively (8). These sequences are thought to be involved in binding to CD81 and SR-B1 receptors (39,48) where monoclonal or polyclonal Abs targeting these epitopes have been shown to mediate virus neutralization (8,57).
In conclusion, the humoral responses, viral neutralization and specific binding assays suggest that the three HCV envelope MAP described in this study are more immunogenic than linear peptides; therefore, they can be essential components for HCV vaccine.
Footnotes
Acknowledgment
The authors would like to thank Dr. Khaled Atef for his kind assistance.
Author Disclosure statement
No competing financial interests exist.