
Abstract
The study was aimed to estimate the role and prevalence rates of genotypes, haplotypes, and alleles, located within the single-nucleotide polymorphisms (SNPs) of interleukin (IL) 1A, IL1B, and IL6 genes, in the occurrence and development of human cytomegalovirus (HCMV) infection among pregnant women. A research was conducted in 129 pregnant women, out of whom, 65 were HCMV infected and 64 were age-matched control uninfected individuals. HCMV DNA was quantitated for UL55 gene by the real-time Q PCR in the body fluids. The genotypic statuses within the SNPs were determined by nested PCR-RFLP assays and confirmed, by sequencing for randomly selected representative PCR products. A relationship between the genotypes and alleles, as well as haplotypes and multiple variants in the studied polymorphisms, and the occurrence of HCMV infection in pregnant women, was determined using a logistic regression model. TT genotype within IL1A polymorphism significantly decreased the risk of HCMV infection (OR 0.32, 95% CI 0.09–1.05; p ≤ 0.050). Considering IL6 SNP, the prevalence rate of GC genotype was significantly decreased among the HCMV infected, compared to the uninfected control individuals (OR 0.45, 95% CI 0.21–0.99; p ≤ 0.050). Moreover, CC homozygotic status in IL6 SNP, found in pregnant women, significantly decreased the risk of congenital infection with HCMV in their offsprings (OR 0.12; p ≤ 0.050). In multiple SNP analysis, TC haplotype within the IL1 polymorphisms significantly decreased the risk of the infection in pregnant women (OR 0.38 95% CI 0.15–0.96; p ≤ 0.050). In addition, TTG complex variants for all the studied polymorphisms and TG variants for IL1B and IL6 SNPs were significantly more prevalent among the infected offsprings with symptomatic congenital cytomegaly than among the asymptomatic cases (p ≤ 0.050). In conclusion, the analyzed IL1A −889 C>T, IL1B +3954 C>T, and IL6 -174 G>C polymorphisms may be associated with the occurrence and development of HCMV infection among studied patients.
Introduction
T
A few studies showed an important role of IL1α, IL1β, and IL6 cytokines in the immunity against HCMV (6,9,49). Significantly increased IL1β levels were found in amniotic fluids from congenitally HCMV-infected patients (41). In allogeneic hematopoietic stem cell transplantation recipients, HCMV infection resulted in decreased blood levels of both IFN-γ and IL1β, while the lowered levels of IL1β and IL10 could be related to decreased survival rates of the patients (12). The immediate early 2 (IE2) protein of HCMV was reported to induce transcription of the IL1B gene, encoding IL1β in monocytes (28). The function of IE2 was dependent on the interactions, localized at two C/EBP sites in the IL1B promoter (28).
An in vitro study, performed for gingival fibroblasts, showed some relationship between HCMV infection and the production of IL1α, IL1β, IL6, and other cytokines, with an increased expression of IL1β mRNA after the infection, as well as with an elevated production of IL1β in HCMV-positive periodontitis specimens (6). In HCMV-infected villous syncytiotrophoblast, the paracrine release of IL1β cytokine and TNFα was found to be involved in the stimulation of intercellular adhesion molecule (ICAM)-1 expression after the infection (9).
In HCMV-infected pregnant women with early-onset preeclampsia with hemolysis, elevated liver enzyme, and the low platelets syndrome, the HCMV IgG antibody index was associated with the production of IL6 and TNFα cytokines (48). Similarly, higher expression levels of IL6, as well as of TNFα, were also observed in Chinese pregnant women infected with HCMV (49). A study performed on trophoblasts from term and 1st trimester placentas, as well as on choriocarcinoma cell lines, infected with HCMV, reported IL6 to be upregulated by the infection, but independent of complete virus replication (21).
Considering genetic alterations located in IL1 or IL6 genes, some articles mentioned their involvement in various infectious diseases, including Helicobacter pylori infections in patients with gastritis, as well as severe malaria in the Vietnamese (7,16,35). However, no previous article has successfully correlated genetic alterations located in IL1 or IL6 genes with HCMV infection among pregnant women, during pregnancy. In the reported study, we aimed to investigate a possible role of the single-nucleotide polymorphisms (SNPs) from IL1A, IL1B, and IL6 genes in the development of HCMV infection in pregnant women during gestation. Moreover, a relationship was estimated between maternal genetic status within the analyzed SNPs and the occurrence and development of congenital HCMV infection in their fetuses and neonates.
Materials and Methods
The study was performed with participation of 129 pregnant women, out of whom, 65 were infected with HCMV during pregnancy and 64 were age-matched control uninfected individuals, between 18 and 41 years (mean 28.52 years). The mean age of infected pregnant women and uninfected individuals was 28.29 ± 5.29 and 28.75 ± 4.79 years, respectively (Table 1).The pregnant women were treated at the Department of Fetal-Maternal Medicine and Gynecology at the Polish Mother's Memorial Hospital - Research Institute in Lodz, between the years 2002 and 2014.
n, number of tested pregnant women.
In case of HCMV-infected pregnant women, the serological status was obtained for 58 individuals.
n, number of offspring was 67.
HCMV, human cytomegalovirus; IUGR, intrauterine growth restriction; SD, standard deviation.
The clinical materials used in genetic studies contained whole blood, plasma or serum, and urine specimens. Intragestational HCMV infection was diagnosed by the serological status for anti-HCMV antibodies and by cytomegaly-related ultrasound markers, observed in the fetuses of affected women. The intrauterine congenital infections of fetuses were determined in 18 out of 65 (27.7%) HCMV-infected pregnant women. Symptomatic cytomegaly was found in 15 of 67 (22.4%) diagnosed offsprings, of 65 HCMV-infected pregnant women (one pregnant women gave birth to triplets), while the asymptomatic infection was found among five of 67 (7.5%) cases.
The symptoms associated with congenital disease included microcephaly (1.49%), ventriculomegaly (1.49%), respiratory failure (7.46%), heart defects (5.97%), hepatitis (1.49%), ascites (2.96%), intrauterine growth restriction (IUGR, 8.96%), as well as fetal/neonatal death (10.45%, Table 1). The active infection with HCMV was confirmed by the presence of viral DNA in body fluids of 15 pregnant women, as well as in amniotic fluids or umbilical cord blood specimens of their 10 offsprings. The research was approved by the Research Ethics Committee at the Polish Mother's Memorial Hospital - Research Institute. All the samples, previously collected for diagnostic purposes, are anonymized in this report. Informed consent forms were signed by all the enrolled pregnant women.
Serological tests
Blood specimens were collected from the pregnant women by venipuncture on their first visit to the Hospital, between the 5th and 38th week of gestation (mean 22.14 weeks). The mean gestational age of HCMV-infected pregnant women and uninfected individuals was 22.50 ± 9.21 weeks and 21.63 ± 9.02 weeks, respectively (Table 1). Serum samples were obtained by centrifugation and then stored at 4°C until analysis, on the day of blood collection. Serological tests were performed at the Hospital's Department of Clinical Microbiology.
Serological status for anti-HCMV IgG and IgM antibodies was tested with VIDAS CMV IgG and IgM tests (bioMérieux)—used between 2002 and 2006, anti-CMV IgG and IgM tests (Diasorin/Biomedica)—used between 2006 and 2011 years, and ELFA assays—from the year 2012, as previously described (47). The occurrence of HCMV infection during pregnancy was determined by IgG seroconversion, the presence of IgG- and IgM-specific antibodies, or a low IgG avidity index. Among HCMV-infected pregnant women, IgG seropositivity was confirmed in 96.55% (56/58) of cases, while the IgM antibodies suggestive of recent infection were determined in 86.21% (50/58) of the patients (Table 1). In turn, the control group included uninfected pregnant women seronegative for both IgG and IgM antibodies against HCMV.
DNA extraction
Genomic and viral DNA were extracted from 200 μL whole blood, and plasma or serum specimens, as well as from 5 mL urine samples, using a QIAamp DNA Mini Kit (QIAGEN). The isolated DNA was diluted in 100 μL of elution buffer and stored at −20°C until further molecular analyses.
Quantitative analysis of HCMV DNA loads
Among HCMV-infected patients, the viral DNA loads in the study body fluids were determined by the real-time Q PCR assay for UL55 gene fragment of HCMV genome, as described previously (22,38,47). The reactions were performed in triplicate and the PCR conditions were as follows: initial activation for 10 min at 95°C, 50 cycles of repeated denaturation at 95°C for 15 sec, and annealing at 60°C for 1 min. The standard curves were obtained from serial 10-fold dilutions from 105 to 1 plasmid DNA, containing the entire HCMV UL55 open reading frame (43). The amplification was performed by a 7900HT Fast Real-Time PCR System (Applied Biosystems).
SNPs genotyping of IL1A, IL1B, and IL6 genes
Genotypes in IL1A −889 C>T, IL1B +3954 C>T, and IL6 -174 G>C polymorphic sites were determined, using self-designed nested PCR assays, as previously described (45). The sequences of external and internal primers, amplicon lengths, and annealing temperatures were used according to previous data (45). External primer sequences were developed, using the Vector NTI Suite 5.5 software, while the internal primers were based on published data (10,14,19, 26,27,45). The amplifications were performed with a HotStarTaq® Master Mix Kit (Qiagen).
In brief, the PCR conditions were as follows: an initial activation for 15 min at 95°C and for 40 cycles of repeated denaturation at 94°C for 30 sec, annealing at temperatures designed for the used primer pairs, for 1 min, extension at 72°C for 2 min, and final extension at 72°C for 10 min. The resulting amplicons were resolved on 1% agarose gels, stained with ethidium bromide, followed by overnight restriction digestion with NcoI, TaqI, or Hsp92II endonucleases, used for determination of genotypes in the range of IL1A −889 C>T, IL1B +3954 C>T, or IL6 -174 G>C SNPs, respectively. The digested products were resolved on 2% agarose gels. Genotypes in the analyzed polymorphic sites were profiled, based on the length of restriction fragments [(10,14,19, 26,27,45); Fig. 1].
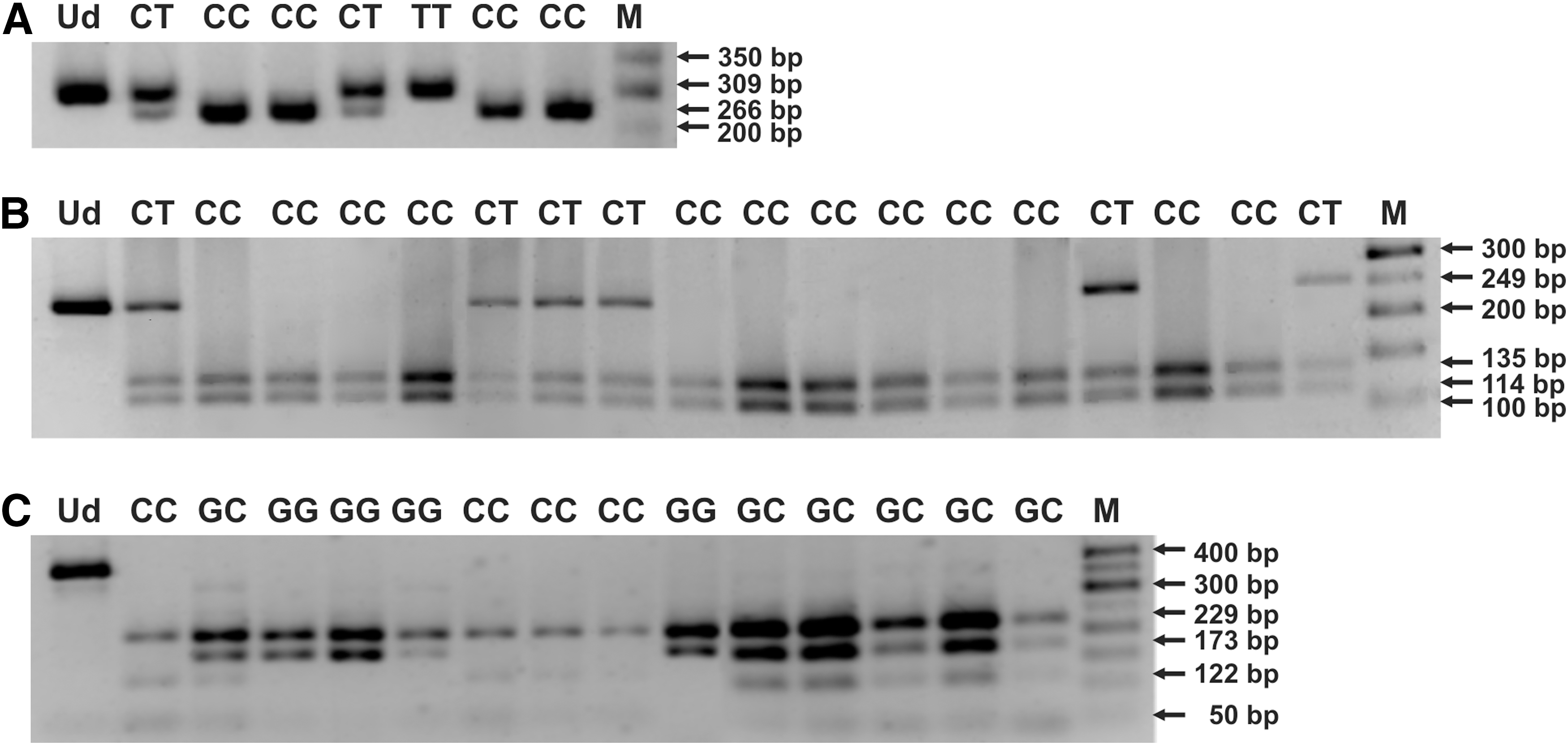
Electropherograms for PCR-RFLP profiles of distinct genotypes in IL1A −889 C>T
The randomly selected PCR products, representing distinct genotypes in IL1 and IL6 SNPs, were also sequenced according to the Sanger method, at the Genomed Joint-Stock Company, to verify the nested PCR-RFLP outcomes. The sequencing was performed for six and five CC homozygotes, six and six CT heterozygotes, as well as three and four TT homozygotes in IL1A and IL1B SNPs, respectively. In case of IL6 SNP, two GG homozygotes, three GC heterozygotes, and two CC homozygotes were sequenced in the analyzed polymorphism. The exemplary chromatograms with fragments of DNA sequences, containing distinct genotypes within the analyzed polymorphisms, are shown in Figures 2 and 3. The sequenced and referenced DNA sequences were compared using the BLASTN program for alignment of two (or more) sequences. The chromatograms were read out using the Chromas Lite 2.1.1 software.

DNA sequences, comprising IL1A −889 C>T
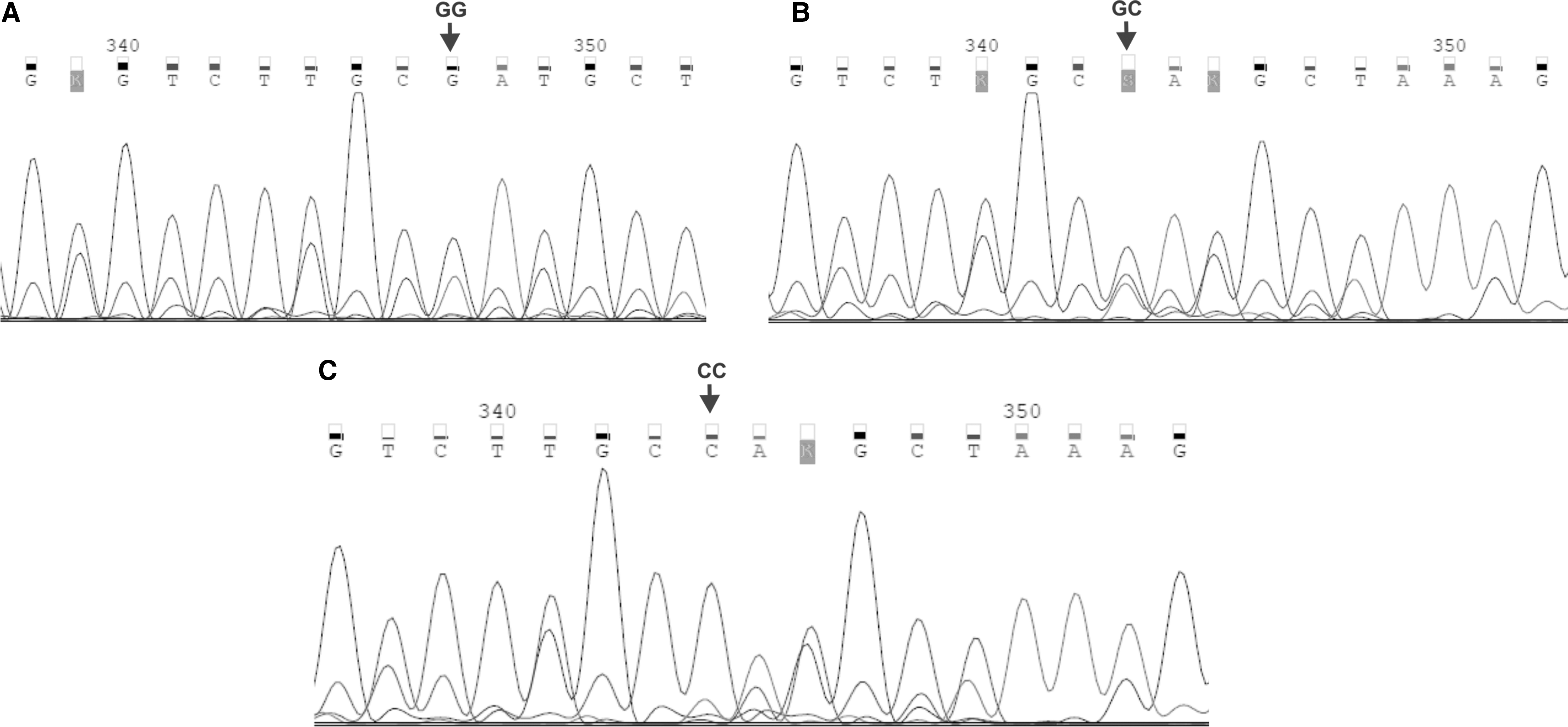
Chromatograms of DNA sequences, encompassing IL6 -174 G>C polymorphism
Statistical analysis
Distribution of genotypes and alleles in IL1A, IL1B, and IL6 SNPs was determined in the analyzed groups of pregnant women by means of descriptive statistics. The patients were studied for the Hardy–Weinberg (H-W) equilibrium, the linkage disequilibrium, haplotypes, and multiple genotypic variants, using the SNPStats software (
The outcomes of the study were estimated as statistically significant when the significance level of p ≤ 0.050 was attained. The statistical analysis was, in part, supported by the NCSS 2004 software.
Results
Hardy–Weinberg equilibrium, linkage disequilibrium
Taking into account IL1A −889 C>T SNP, the genotypes preserved the H-W equilibrium among pregnant women infected with HCMV (p = 0.760), while the equilibrium was not determined for the uninfected patients (p ≤ 0.050). Regarding IL1B +3954 C>T polymorphism, the H-W equilibrium was observed among both HCMV-infected and uninfected pregnant women (p = 1.000 and p = 0.740, respectively). Moreover, the IL1A and IL1B SNPs were seen in linkage disequilibrium among the studied groups of pregnant women (p ≤ 0.050). In case of IL6 -174 G>C polymorphism, the genotypes were not seen in the H-W equilibrium among the patients infected with HCMV (p ≤ 0.050), while the equilibrium was preserved in uninfected pregnant women (p = 0.180).
HCMV DNA loads in clinical samples
In maternal whole blood specimens, the median load of HCMV DNA was 3.8 × 102 copies/mL and ranged from 1.3 × 102 to 1.1 × 103 copies/mL, while the mean viral load was 5.2 × 102 copies/mL (Table 1). In plasma samples, the median viral load was 5.6 × 102 copies/mL and ranged from 1.8 × 102 to 3.6 × 103 copies/mL, whereas the mean viral load was 1.4 × 103 copies/mL. Considering urine samples, the median HCMV DNA load was 4.4 × 102 copies/mL and ranged from 1.5 × 102 to 2.4 × 103 copies/mL, and the mean viral load was 3.2 × 103 copies/mL. In case of fetal amniotic fluids, the median HCMV DNA load was 9.9 × 102 copies/mL ranging from 2.2 × 102 to 1.5 × 103 copies/mL, while the mean viral load was 9.2 × 102 copies/mL. In umbilical cord blood specimens, the median HCMV DNA load was 3.2 × 103 copies/mL, ranging from 1.6 × 102 to 6.6 × 103 copies/mL, and the mean viral load was 3.3 × 103 copies/mL.
Genotypes, haplotypes, and multiple-SNP variants in IL1 and IL6 SNPs
Among HCMV-infected pregnant women, the prevalence rates of CC, CT, and TT genotypes in IL1A −889 C>T SNP were 51.6% (33/64), 42.2% (27/64), and 6.2% (4/64), respectively (Table 2). Considering IL1B +3954 C>T polymorphism, the prevalence rates of CC, CT, and TT genotypes were 60.0% (39/65), 35.4% (23/65), and 4.6% (3/65), respectively. In case of IL6 -174 G>C SNP, the prevalence rates of GG, GC, and CC genotypes were 58.7% (37/63), 22.2% (14/63), and 19.1% (12/63), respectively.
n, number of tested pregnant women.
Logistic regression model; p ≤ 0.050 is considered significant. The statistically significant values are underlined.
OR, odds ratio; 95% CI, confidence interval.
Regarding uninfected control pregnant women, the analyzed genotypes in IL1A polymorphism were found in 54.0% (34/63), 28.6% (18/63), and 17.5% (11/63), of the patients, respectively. In case of IL1B polymorphism, the studied genotypes were confirmed for 55.6% (35/63), 39.7% (25/63), and 4.8% (3/63) of the uninfected controls, respectively. Regarding IL6 SNP, the prevalence rates of GG, GC, and CC genotypes were 41.9% (26/62), 38.7% (24/62), and 19.4% (12/62), respectively.
The occurrence of TT genotype in IL1A −889 C>T SNP significantly decreased the risk of HCMV infection among pregnant women (OR 0.32, 95% CI 0.09–1.05 in the recessive model; p ≤ 0.050; Table 2). In case of IL1B +3954 C>T SNP, distinct genotypes were distributed with similar frequency among the infected pregnant women and uninfected control pregnant women. Taking into account IL6 SNP, the prevalence rate of GC genotype was significantly decreased among HCMV-infected women, compared to uninfected pregnant women (OR 0.45, 95% CI 0.21–0.99 in the overdominant model; p ≤ 0.050; Table 2). Moreover, CC homozygotic status in IL6 polymorphism, found in pregnant women, significantly decreased the risk of congenital infection with HCMV in their offsprings (OR 0.12 95% CI 0.01–1.00 in the codominant model, or 0.01–1.01 in the recessive model; p ≤ 0.050; Table 3). In case of IL1A and IL1B polymorphisms, distinct genotypes were distributed similarly between mothers of congenitally infected and uninfected fetuses and neonates. All the studied genotypes within IL1 and IL6 polymorphisms were similarly distributed among HCMV-infected mothers of both congenitally infected and uninfected offsprings.
n, number of tested pregnant women.
Logistic regression model; p ≤ 0.050 is considered significant. The statistically significant values are underlined.
In multiple SNP analysis, the TC haplotype in the range of the analyzed two polymorphisms within IL1 genes significantly decreased the risk of HCMV infection among pregnant women (OR 0.38 95% CI 0.15–0.96; p ≤ 0.050; Table 4). Both TT and CT haplotypes in the analyzed IL1 polymorphisms were distributed similarly among the infected and uninfected patients. Multiple SNP variants in the range of all the analyzed IL1A, IL1B, and IL6 polymorphisms were also distributed similarly among the studied groups of pregnant women. However, both the TTG complex variants for IL1A, IL1B, and IL6 polymorphisms (OR 5.11 95% CI 1.48–17.62; p ≤ 0.050) and the TG variants for IL1B and IL6 (OR 5.68 95% CI 1.76–18.32; p ≤ 0.050) SNPs were significantly more prevalent among the infected offsprings with symptomatic congenital cytomegaly, compared to the asymptomatic controls.
Logistic regression model; p ≤ 0.050 is considered significant. The statistically significant values are underlined; global haplotype association p-value was estimated to be 0.028.
SNPs, single-nucleotide polymorphisms.
Distribution of alleles in IL1A, IL1B, and IL6 SNPs
Among the infected pregnant women, the prevalence rates of C and T alleles in IL1A −889 C>T SNP were 73.0% (93/128) and 27.0% (35/128), respectively. In case of IL1B +3954 C>T polymorphism, the prevalence rates of alleles C and T were 78.0% (101/130) and 22.0% (29/130), respectively. Taking into account IL6 -174 G>C SNP, the prevalence rates of G and C alleles were 70.0% (88/126) and 30% (38/126), respectively.
Regarding the uninfected control pregnant women, the prevalence rates of C and T alleles in IL1A SNP were 68.0% (86/126) and 32.0% (40/126), respectively. In the range of IL1B variation, the studied alleles were observed with prevalence rates of 75.0% (95/126) and 25.0% (31/126), respectively. In case of IL6 polymorphism, the observed prevalence rates of G and C alleles were 61.0% (76/124) and 39.0% (48/124), respectively. The distribution of distinct alleles in the analyzed IL1 and IL6 SNPs was similar among the infected pregnant women and uninfected control pregnant women.
Discussion
In the reported study, TT genotype in IL1A −889 C>T SNP, defined in the recessive model of inheritance, as well as TC haplotype in the range of both the analyzed IL1A and IL1B polymorphisms, were found to be associated with a decreased risk for the occurrence of HCMV infection in pregnant women.
So far, no previous study revealed the role of IL1A and IL1B SNPs in the development of HCMV infection, although the contribution of the encoded IL1α and IL1β cytokines to the disease was confirmed (28,41). Similar to our study, a research performed on 2-year-old Jamaican children, infected with H. pylori, reported a relationship between the presence of the minor allele in IL1A −889 C>T polymorphic site and a reduced risk for the infection (44). Among the Jamaican children, infected with H. pylori, the prevalence rates of distinct genotypes in the IL1A −889 C>T region were similar to those in our study among the HCMV-infected pregnant women (44).
Considering the functional role of the minor allele in IL1A −889 C>T SNP, a significantly higher transcriptional activity of the gene was previously found for TT carriers, compared to CC genotype in the analyzed polymorphism (15). The reported TT genotype in IL1A SNP was associated with slightly elevated plasma levels of IL1A mRNA and encoded protein (15). The relationship between the presence of TT genotype in IL1A −889 C>T locus and an increased expression of the cytokine was also found in severe periodontal disease, as well as in chronic iridocyclitis in juvenile rheumatoid arthritis (31,42). Another study also showed a correlation of TT genotype in IL1A −889 C>T SNP with the occurrence of osteomyelitis (1).
In turn, IL1B +3954 C>T polymorphism was reported to be associated with tuberculosis and the lipodystrophic syndrome (LD) in human immunodeficiency virus (HIV)-infected Caucasian patients (2,20,32). In household contacts with tuberculous patients, the presence of CC homozygotic genotype in IL1B +3954 C>T SNP and GA heterozygotic variant in IL10 -1082 G>A SNP correlated with an increased risk of disease development (32). Among HIV-infected patients on highly active antiretroviral treatment, the carriers of T allele in IL1B variation were reported to have a decreased risk of developing LD (2). In turn, a similar effect on LD was determined among TT homozygotic and TC heterozygotic patients in the range of the analyzed polymorphism (2).
In our study, the genotypic status in IL1B +3954 C>T polymorphism had no significant influence on HCMV infection in pregnant women. However, the relationship between TC haplotype in both the analyzed IL1A and IL1B SNPs and the occurrence of HCMV infection may indicate a possible association of IL1B SNP with the infection as well. This seems to be caused by linkage disequlibrium between IL1B and IL1A polymorphisms, since both genes are localized very close in the long arm of chromosome 2 (2,34). In turn, IL1A SNP may alter the immune response against HCMV infection through creation of a consensus site for, at least, one additional transcription factor, Skn-1 (15). In addition, the presence of the TT genotype in the polymorphic site was reported to have resulted in the elevated expression of IL1α (15).
It seems that more detailed studies could be beneficial to reveal the mechanism by which both analyzed IL1A and IL1B SNPs associate with the occurrence of HCMV infection among pregnant women.
Regarding IL6 -174 G>C polymorphism, GC heterozygotic status, estimated in the overdominant model, was determined to be correlated with lowered risk of HCMV infection among studied pregnant women. In addition, pregnant women with the CC homozygote significantly less frequently gave birth to offsprings congenitally infected with HCMV, compared to women with GG or GC genotypes within the analyzed polymorphism, observed in the codominant and recessive models. Considering multiple SNP analysis, both TTG complex variants in the range of all the three (IL1A, IL1B, and IL6) polymorphisms, as well as TG variants for IL1B and IL6 SNPs, were significantly associated with an increased risk for the occurrence of congenital symptomatic cytomegaly, compared to the asymptomatic disease.
Previously, GC heterozygotes in IL6 -174 G>C polymorphism were determined to have a protective role against the occurrence and recurrence of condyloma acuminatum (3). In turn, brucellosis related focal complications were associated with GC genotype in IL6 SNP, while the function against disease development was typical for the occurrence of GG genotype in the polymorphism (24). The prevalence rates of distinct genotypes in IL6 SNP were similar between the group of patients with brucellosis and controls (24). The reported prevalence rates of GC heterozygotes among control cases without brucellosis were comparative to the prevalence rates, observed in uninfected pregnant women in our research (24). Several studies also showed CC genotype in IL6 -174 G>C polymorphism to be associated with neuroprotection through decreased IL6 gene expression and lower IL6 plasma levels (13,36).
However, no such relationship was also observed among both, patients with a first myocardial infarction before the age of 60 as well as the Caucasian lung transplant recipients (4,33). In hematological patients with invasive pulmonary aspergillosis, the genotypic status in IL6 SNP was reported to have no influence on susceptibility to the disease (39). Therefore, the role of IL6 -174 G>C SNP in immune response seems to vary between different diseases.
Considering previous data, the analyzed IL6 polymorphism might be associated with HCMV infection, through generation of new binding sites for NF-1 and Smad4 transcription factors, related to the presence of the C allele at the polymorphic site, as well as by the altered expression of the encoded protein (17,40,45). Moreover, a multiple SNP analysis showed a possible common involvement of IL6 polymorphism, together with IL1A and IL1B SNPs analyzed in the reported study, in the development of congenital symptomatic HCMV infections in the fetuses and neonates of infected pregnant women. We suggest that further studies on a detailed mechanism of the relationship between IL6 -174 G>C, as well as IL1A and IL1B polymorphisms, and the occurrence and development of HCMV infection among both pregnant women and their offsprings, would be justified.
Footnotes
Acknowledgments
Sponsor: The Polish Ministry of Science & Higher Education, Polish Mother's Memorial Hospital–Research Institute–Grant supporting statutory research No. 2015/II/16-MN.
Author Disclosure Statement
No competing financial interests exist.