
Abstract
Toll-like receptor 3 (TLR3) is one of the TLRs whose ligand is double-stranded RNA (dsRNA). Infectious bursal disease virus (IBDV) is a dsRNA virus that could be recognized by TLR3. The purpose of this study was to determine the role of the virulence of IBDV on the expression of chicken TLR3 (chTLR3). For this purpose, the levels of chTLR3 expression and its downstream effectors, Interferon β (IFN-β) and Interleukin 8 (IL-8), were detected and analyzed after infection of IBDV field isolates with differential virulence in vitro (chicken embryo fibroblast and/or chicken peripheral blood mononuclear cells) and in vivo (commercial Three-Yellow chicken). The results showed that chTLR3 was activated by IBDV, resulting in the expression of antiviral IFN-β and chemokine IL-8. The expression of chTLR3, IFN-β, and IL-8 correlated well with the virulence of IBDV as the more virulent the IBDV strain that was used, the more pronounced was the expression of chTLR3, IFN-β, and IL-8. These results suggest that chTLR3 is involved in the pathogenesis of IBDV in commercial chickens and its downstream effectors (IFN-β and IL-8) might play an important role in this process.
Introduction
I
Toll-like receptor 3 (TLR3) is a member of the TLR family, which plays a fundamental role in pathogen recognition and activation of innate immunities. Chicken TLR3 (chTLR3) is confirmed to be represented as a true ortholog of mammalian TLR3 (35). ChTLR3 is able to bind double-stranded RNA (dsRNA) derived from virus, leading to activation of proinflammatory cytokines and chemokines, which would play an important role in chicken antiviral responses during viral infection (1,11). Once infection occurs, viral dsRNAs are often recognized by TLR3 found in immune cells. Recognition of microbial components by TLRs in immune cells initiates signal cascades, which trigger expression of genes that produce proinflammatory cytokines and costimulatory molecules (40). Type I IFN and chemokine IL-8 are two important downstream effectors that could be induced followed by TLR3 activation, regulating host immune response to infection, and play an important role in controlling and eliminating invading pathogens (1,14,40).
It has been documented that IBDV can be recognized by chTLR3 (3,12). It has been confirmed that the virus can replicate in immature bursa-derived B-lymphocyte, monocyte/macrophage, and even NK cell of chicken (23,24,25). During an IBDV infection, the immune system response is triggered by the interaction between viral dsRNA and chTLR3, which leads to activation of proinflammatory cytokines and chemokines. The expression of chTLR3 can be demonstrated either in vitro or in vivo. Iqbal's group (20) revealed that chTLR3 shows a broad pattern of tissue expression, including bursa and bone marrow. Rauf et al. (33) found that the expression of chTLR3, IFN-β, and chemokine IL-8 was more pronounced in cIBDV-infected chicken than the avIBDV-infected group. Liu et al. (28) revealed that some of the Th1 and Th2 cytokines were differentially upregulated after infection with vvIBDV. Similarly, Eldaghayes et al. (7) demonstrated that a proinflammatory response was induced to a greater degree by vvIBDV than cIBDV and they suggested that IBDV strains with different virulence modulated the host's immune response by using a different mechanism.
At the present time, little is known about the innate immune responses triggered by the TLR signaling pathway involved in the pathogenesis of IBDV. This study was aimed at determining the correlation between chTLR3 expression and the virulence of IBDV by both in vitro and in vivo experiments (in commercial chickens).
Materials and Methods
Viruses
IBDV field isolates used in this study included NN1172 (vvIBDV isolated in Guangxi, China in 2011), NN040124 (intermediated plus virulent strain isolated in Guangxi, China in 2004), and B87 (in) (intermediated virulent strain, a commercial vaccine strain commonly used in China).
Cell cultures
Chicken embryo fibroblast (CEF) cells were maintained in Dulbecco's modified Eagle's medium containing 10% fetal calf serum (Gibco BRL) and antibiotics (100 U penicillin/mL and 100 μg streptomycin/mL) at 37°C, 5% CO2, and 95% humidity. Chicken peripheral blood mononuclear cells (PBMCs) were separated from peripheral blood collected from healthy Three-Yellow chicken using Ficoll-PaqueTM PREMIUM (Armersham Biosciences, Sweden) and maintained in RPMI 1640 containing 10% fetal calf serum (Gibco BRL), 5% chicken serum (Gibco BRL), and antibiotics (100 U penicillin/mL and 100 μg streptomycin/mL) at 37°C, 5% CO2, and 95% humidity, as described by Kogut et al. (26) and He et al. (13).
Chickens
One-day-old commercial Three-Yellow chickens without vaccination were purchased and transferred to an isolator. All the chickens were tested to be free of maternally derived IBDV antibodies by using a commercial ELISA kit, Flock Check Infectious Bursal Disease Antibody Test Kits (IDEXX Laboratory, Inc.), before virus inoculation. Chickens were reared in isolation units for the duration of the study and were housed in separate isolation units after the challenge. The animal experiments were conducted in accordance with the International Guiding Principles for Biomedical Research Involving Animals as issued by the Council for the International Organizations of Medical Sciences.
Experiment design
Experiment 1 was designed to test the abilities of the IBDV field strain modulating the expression of chTLR3 in vitro (both PBMCs and CEFs). In this experiment, cells were infected by the IBDV field strain NN040124 in a titer of 50TCID50, and the infected cells were harvested at 1.5, 3, 6, 12, 24, and 48 h postinfection (hpi). The viral RNA and the expression of chTLR3, IFN-β, and IL-8 were detected by quantitative real-time polymerase chain reaction (qRT-PCR). Subsequently, PBMCs and CEFs were infected with different IBDV field strains with different virulence, including NN1172 and NN040124, and a vaccine strain B87 (in), respectively, in a titer of 50TCID50. The cells were harvested at the time when the viral load level was the highest. The viral RNA and the expression of chTLR3, IFN-β, and IL-8 were detected by qRT-PCR or Western blot or by both methods.
Experiment 2 was designed to test the differential expression of chTLR3, IFN-β, and IL-8 induced by vvIBDV field strain and vaccine strain B87 (in) in commercial chickens. In this experiment, 90 four-week-old commercial Three-Yellow chickens were randomly divided into three groups with 30 birds per group. Birds in Group 1 and Group 2 were inoculated intraocularly with NN1172 and B87 (in) in a dose of 105TCID50, respectively, and Group 3 was inoculated through the same route with PBS that served as a mock infection control. Five birds per group were bled and sacrificed, respectively, at 1, 2, 3, 5, and 7 days postinfection (dpi). The individual bursa sample was collected from each bird, and the bursa/body weight (B/BW) ratio was determined according to the methods described by Ismail and Saif (21) using the following formula: (organ weight in grams × 1,000)/BW in grams. The viral load and the expression of chTLR3, IFN-β, and IL-8 were detected by qRT-PCR using these bursa samples.
RNA extraction and real-time quantitative PCR
PBMCs or bursal samples collected from birds of infected and uninfected groups were subjected to RNA extraction by using Ultrapure RNA Kit (CWBIO, Beijing) according to the manufacturer's instructions. Briefly, for viral RNA extraction, the cells or bursa samples (after homogenization) were frozen and thawed three times and centrifuged at 3,000 × g for 10 min. Two hundred microliters of the supernatants were suspended in 1 mL of TRIzol reagent and the total RNA was prepared according to the manufacturer's instructions. Finally, total RNA extracted from each sample was suspended in 30 μL of water. For total RNA extraction, either the cells (from one flask) or homogenized bursa sample suspension (200 μL) was suspended in 1 mL of TRIzol reagent. Then, 1 μg of RNA sample was used for cDNA synthesis using HiFiScript 1st Strand cDNA Synthesis Kit (CWBIO, Beijing) according to the manufacturer's instructions.
qRT-PCR was performed on cDNA using SYBR Green I dye in an IcyclerQ5 (Bio-Rad) based on the established protocol previously described (15,16). Specific sequences of primers were used for the amplification of IBDV, chTLR3, IFN-β, IL8, and internal control gene GAPDH as listed in Table 1.
Western blot
Approximately 106 infected/uninfected PBMCs/CEF cells were pelleted and lysed in 100 μL of sodium dodecyl sulfate (SDS) buffer (0.5% SDS, 0.05 M Tris 7.5, 1 mM dithiothreitol). One percent of the cell extract was resolved by SDS-10% polyacrylamide gel electrophoresis, and the gel was transferred to a nitrocellulose membrane. chTLR3 and β-actin were detected by a mouse polyclonal antibody against chTLR3 (17) and antiactin antibody (Invitrogen), respectively. Immunoreactive bands were detected using an anti-mouse IgG HRP-labeled antibody (Invitrogen; 1:10,000 dilution in TBST) and DAB substrate solution. The density of the bands was scanned and analyzed by Gel Imaging System (Bio-Rad).
Statistical analysis
All values are reported as means ± standard errors (standard deviation). Statistical significance was calculated using one-way analysis of variance whenever multiple groups were compared. For individual comparisons of multiple groups, Student–Newman–Keuls post hoc test was used to calculate p values. Means with p values of <0.05 were considered significantly different. All statistical calculations were performed using Primer of Biostatistics (9)
Results
Detection of IBDV RNA in PBMCs
We sought to determine if IBDV field strain could infect and replicate in PBMCs. For this purpose, PBMC was infected with field strain NN040124 (an intermediated plus virulent strain), and the kinetics of viral RNA was detected by qRT-PCR. The results showed that the viral RNA load did not change within 1.5 hpi; however, the viral load increased significantly at 3 hpi and reached the highest level at 6 hpi with 1291.7 copies/μL. Although the viral load decreased afterward, the viral load remained at a high level until 48 hpi, as shown in Figure 1.
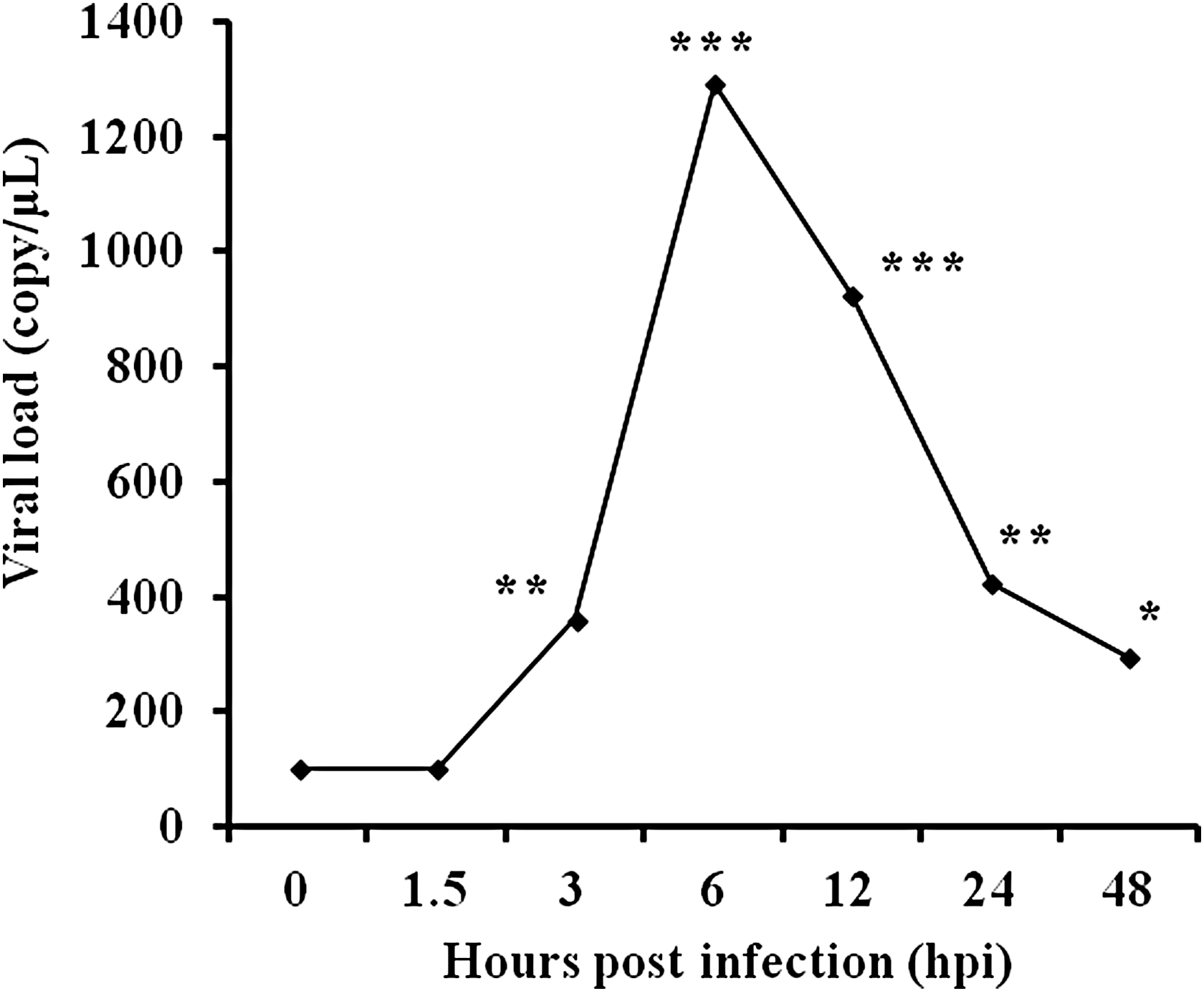
The replication kinetics of dsRNA of IBDV strain NN040124 in PBMCs. PBMCs were infected with IBDV field strain NN040124 and the cells were harvested at different time points after infection. The viral load was represented by viral RNA copies per μL. Note: the viral RNA load increased significantly at 3 hpi and reached the highest level at 6 hpi. *p < 0.05, **p < 0.01, ***p < 0.001, between the infection group and the mock-infected control, n = 5. IBDV, infectious bursal disease virus; PBMC, peripheral blood mononuclear cell.
ChTLR3 is modulated by IBDV field strain in PBMCs
ChTLR3 is an important member of pathogen-associated molecular pattern (PAMP) receptor and was reported to bind with dsRNA. IL-8 and IFN-β are important downstream effectors in the chTLR3 signaling pathway. To detect if chTLR3, IL-8, and IFN-β were expressed in response to IBDV infection in PBMCs, IBDV field strain NN040124 was used to infect PBMCs; afterward, the mRNA expression levels of chTLR3, IL-8, and IFN-β were detected by qRT-PCR. As shown in Figure 2, the expression levels of ChTLR3, IL-8, and IFN-β were all upregulated on mRNA level after NN040124 infection. The highest expression level of chTLR3 was detected to be 3138.96% of GAPDH and those of IFN-β and IL-8 were 194% and 1,087%, respectively (Fig. 2). The results indicated that IBDV infection was correlated with the chTLR3 signaling pathway. Interestingly, the expression level of IFN-β mRNA peaked at 3 hpi; however, the mRNA level of chTLR3 and IL-8 peaked at 6 hpi. The results suggested that there was an antiviral response against IBDV, as evidenced by IFN-β expression, which occurred earlier than the proinflammatory response.

The expression levels of chTLR3 and the downstream effectors in PBMCs after infection with NN040124. PBMCs were infected with IBDV field strain NN040124 and cells were harvested at different time points after infection. The expression of chTLR3, IFN-β, and IL-8 was detected by qRT-PCR. The mRNA levels were normalized to GAPDH mRNA levels and relative expression was shown as % of GAPDH. *p < 0.05, **p < 0.01, ***p < 0.001, between the infection group and the mock infection control, n = 5. GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Differential expression of chTLR3, IFN-β, and IL-8 induced by IBDVs with different virulence on mRNA level in vitro
To determine whether the expression level of chTLR3 would change after being infected with IBDVs with different virulence in vitro, PBMCs were infected with IBDVs with different virulence, respectively, and the differential replication abilities of the different IBDVs in PBMCs and the expression of chTLR3, IL-8, and IFN-β were detected after infection. We choose 6 hpi as the detecting time point since this time point had been shown to be the most efficient time point for the replication of NN040124 in PBMCs (Fig. 1).
The replication ability results are shown in Figure 3. All the three strains, intermediated virulent strain B87 (in), intermediated virulent plus strain NN040124, and vvIBDV strain NN1172, were effectively replicated in the cultured PBMCs at 6 hpi, with the vvIBDV strain NN1172 replicated as the most efficient, then was the NN040124, and the least efficient replicated strain was attenuated strain B87 (in).

Differential viral loads in PBMCs after infection with IBDVs with different virulence. PBMCs were infected with IBDVs with different virulence and the viral RNA was detected by quantitative reverse transcription-polymerase chain reaction. The viral load was represented by viral RNA copies per μL. ^p < 0.05 between B87 (in) and NN1172; $ p < 0.05 between NN040124 and NN1172; # p < 0.05 between B87 (in) and NN040124, n = 5.
The expression of chTLR3 and the downstream effectors, IFN-β and IL-8, was then detected to determine if IBDV differentially modulated the chTLR3 signaling pathway by its virulence at 6 hpi and the results are shown in Figure 4. After infection of different IBDV strains (B87, NN040124, and NN1172), the expression levels of chTLR3, IFN-β, and IL-8 were all upregulated at 6 hpi. However, the expression levels were not equivalent among strains with different virulence. The chTLR3 mRNA level in the NN1172 group was the highest, followed by NN040124 group, while the B87 (in) group showed the lowest level of expression. The trend of expression of IFN-β and IL-8 was similar to that of chTLR3 (Fig. 4). Importantly, the trend was consistent with that of viral load in PBMCs at the same time point.

Differential expression of chTLR3 and downstream molecules after infection of PBMCs with IBDVs of different virulence. PBMCs were infected with IBDVs with different virulence and the expression levels of chTLR3, IL-8, and IFN-β were detected by qRT-PCR. The mRNA levels were normalized to GAPDH mRNA levels and relative expression is shown as % of GAPDH. *Significant difference between IBDVs and control. *p < 0.05; ***p < 0.001; ^p < 0.05 between B87 (in) and NN1172; $ p < 0.05 between NN040124 and NN1172; # p < 0.05 between B87 (in) and NN040124.
Differential expression of chTLR3 induced by IBDVs with different virulence on protein level in vitro
To determine whether the change in mRNA level of chTLR3 results in change in chTLR3 protein level, ChTLR3 in PBMCs and CEF cells was detected by Western blot after infection of IBDVs with different virulence levels. As shown in Figure 5, the expression level of chTLR3 protein in PBMCs in the NN1172 group was 1.36-fold higher than those in NN040124 and B87(in) groups, and chTLR3 expression in NN040124 was slightly higher than the B87(in) group. All the chTLR3 expression levels in infected groups were higher than that in the mock infection control. Meanwhile, the expression trend of chTLR3 in CEF cells was similar to those in PBMCs. We noted that the expression of chTLR3 protein in both cell types (CEF and PBMC) in NN1172 group was significantly higher than other infection groups, which was consistent with the mRNA level detected by qRT-PCR.

Differential expression of ChTLR3 in PBMCs and CEF cells after infection with IBDVs with different virulence. PBMCs/CEFs were infected with IBDV field strains with different virulence, respectively. The cells were harvested at 6 hpi. The expression of chTLR3 was detected by Western blot. The results are shown as fold expression (chTLR3: β-actin).
Differential expression of chTLR3 and downstream effectors in the chicken bursa of Fabricius after infection of IBDVs with different virulence
To confirm whether the change of chTLR3 expression induced by IBDVs with different virulence in vivo was consistent with the change of chTLR3 expression in vitro, 28-day-old commercial Three-Yellow chickens were infected with vvIBDV strain NN1172 and attenuated strain B87(in). The results showed that NN1172 caused 100% (35/35) clinical signs (e.g., ruffled feathers, hunched posture, lethargy, anorexia, diarrhea, trembling, and dehydration, starting from 3 dpi) and gross lesions (e.g., severe edema and hyperemia of infected bursa at 3 dpi). By contrast, B87(in)-infected birds showed no significant clinical signs; however, 85.7% (30/35) of infected birds exhibited slight gross lesions in the bursa, whereas no clinical signs or deaths were observed in the mock infection control birds. The relative bursa weight of birds in each group at each time point examined is presented in Table 2. The B/BW of the infected chickens increased (p < 0.05) at 3 and 5 dpi, with the NN1172-infected group showing the highest value at both time points, and then decreased at 7 dpi (Table 2).
Data are expressed as the mean ratio ± SD (five chickens and above per group at each time point).
Significant difference between infected group and control group (p < 0.05).
Significant difference between NN1172 and B87 (p < 0.05). n = 5.
SD, standard deviation.
The chTLR3 and downstream effectors, IFN-β and IL-8, were then detected by qRT-PCR, and the results are shown in Table 3. ChTLR3 was significantly upregulated in bursa of birds infected with NN1172 at 3 dpi by 5.36-fold higher than uninfected control (p < 0.05), while no significant changes were observed at other time points. However, expression of chTLR3 in the B87(in) group was significantly downregulated at 1 dpi by 0.23-fold (p < 0.05), but with no significant change at other time points during the experiment. The chTLR3 expression level were significantly higher in the NN1172 group than the B87(in) group at 1 and 3 dpi. The expression level of IFN-β was significantly upregulated in chickens infected with NN1172 at 3 dpi, while it was then downregulated at 7 dpi (p < 0.05). On the contrary, IFN-β in the bursa of birds in the B87(in)-infected group was downregulated at 1 dpi (p < 0.01), but it was then significantly upregulated at 3 dpi. It was noted that the expression of IFN-β in B87(in)-infected group were significantly different from that of the NN1172-infected group: during the first 2 dpi, no obvious change was detected in the NN1172 group, but significant upregulation was observed in the B87(in) group from 1 to 2 dpi. The IFN-β level in the NN1172 group was higher than the B87(in) group (p < 0.05) at 2 dpi. The expression of chemokine IL-8 in the bursa of NN1172-infected birds was higher at 3 dpi by 3.26-fold (p < 0.05), which was significantly higher than B87(in) group. The expression of IL-8 in the B87(in) group was downregulated at 1 and 5 dpi. We noted that the IL-8 level in NN1172-infected birds was generally higher than the B87(in) group, excepting 2 dpi (significantly different at 3, 5, and 7 dpi). These results suggested that both viral strains NN1172 and B87 activated proinflammatory cytokine and chemokine responses and levels of immune responses were associated with the virulence of IBDV infection: stronger immune response was detected in more virulent IBDV strain.
At 1, 2, 3, 5, and 7 dpi, bursa cells were isolated from very virulent (NN1172), attenuated vaccine (B87 (in)), and control chickens and examined for chTLR3, IFN-β, and IL-8 expression by qRT-PCR. Relative expression levels of all genes were calculated as relative to the housekeeping gene GAPDH. The data are presented as fold change relative to the PBS-treated control group.
Comparison of expression between NN1172-infected and B87 (in)-infected birds.
Significant difference between infection group and control group (p < 0.05).
Significant difference between infection group and control group (p < 0.01).
Significant difference between NN1172 and B87 (in) (p < 0.05) n = 5.
The viral load in the bursa was also detected to verify if the differential expression of chTLR3 and downstream effectors correlated with virus replication. As shown in Figure 6, the quantity of viral RNA in the NN1172-infected group increased gradually from 1 to 3 dpi (peak of replication) and then gradually decreased to 7 dpi. The same trend was observed in the B87(in)-infected group. However, the replication of NN1172 in bursa was more effective than the B87(in) group at 3 dpi.
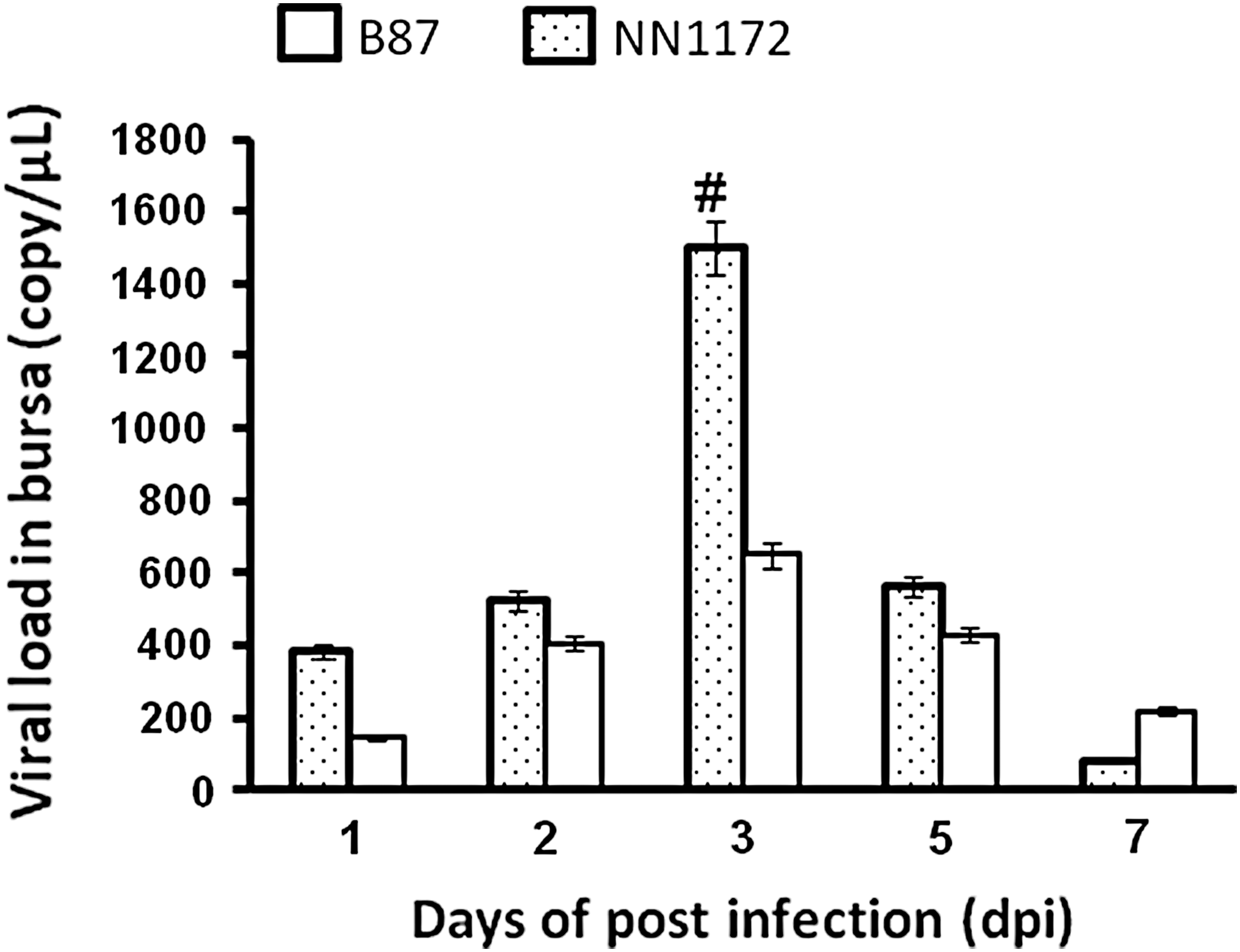
Relative expression at IBDV genome in the bursa. Bursa cells were isolated from chickens infected with IBDVs with different virulence and were examined for IBDV genome by qRT-PCR. The viral load was represented by viral RNA copies per μL. #Statistically significant difference between NN1172-infected and B87 (in)-infected groups (p < 0.05) n = 5.
Discussion
IBDV outbreaks are still common around the world, and commercial chicken flocks often experience losses due to IBDV-induced immunosuppression despite vaccination; even mass vaccination is implemented in some areas such as China. The mechanisms involved in early innate immunity triggered by interaction of IBDV dsRNA genome with chTLR3 are poorly understood both in vitro and in vivo, especially those in commercial chickens.
It has been documented that recognition of PAMPs by TLRs triggers innate host responses and promotes secretion of cytokines, which leads to elicitation of innate and adaptive immune responses (31). TLR3 plays a critical role in the outcome of a variety of viral infections in vitro and in vivo (11,22,34,39) as demonstrated in TLR3-deficient mice that exhibited unusual inflammatory responses following influenza or West Nile virus infection (10,39). TLR3 engagement is important to the outcome of virus infection because of its role in induction and regulation of antiviral mediators such as IFN-β, chemokine IL-8, and proinflammatory gene expression (22,31,36).
TLR3 is a major recognition receptor for virus-derived dsRNA in mammals. Preliminary studies have revealed that the chTLR3 signaling pathway could be activated by IBDV infection (12,32,33,41). In this study, we demonstrated that chTLR3 was upregulated both in mRNA and protein levels in PBMCs when it was infected with IBDV field strain NN040124. Consistently, the expression of chTLR3 was also detected in CEF cells when it was infected with the same IBDV strain NN040124. Intriguingly, when Three-Yellow chickens were infected with vvIBDV strain NN1172, we observed that chTLR3 was increasingly expressed by 5.365-fold (p < 0.05) in the bursa of infected birds at 3 dpi compared with the mock infection group (Table 3). These results of in vitro and in vivo investigations as described above clearly suggested that IBDV infection significantly enhanced the expression of chTLR3. In chickens, TLR3 (recognizes viral dsRNA) and TLR7 (recognizes viral single-stranded RNA) were reported to be the only TLRs implicated in antiviral response (19). In Wong's work (41), chTLR3 was also shown to be correlated with IBDV infection in DF1 cells. In Smith et al.'s (37) work, TLR3 was reported to be upregulated in the IBDV-infected bursa during the early stages of infection. Furthermore, Raj et al. (32) showed that even in different species of infected chickens, the expression pattern of chTLR3 was different after IBDV infection. Taken together, these results suggested that chTLR3 was an important chTLR that was related to IBDV infection both in vitro and in vivo.
Innate immune responses orchestrate the antiviral activities through proliferation of different effector cells, such as cytokine and chemokine-secreted cells (2). TLR3 was also reported to be associated with induction of the antiviral cytokine IFN-β during infections (22,30). In this study, we demonstrated that chTLR3 expression in PBMCs stimulated by IBDV with intermediated plus virulence was upregulated at 6 hpi, but its downstream effector molecule IFN-β was downregulated at this time point (Fig. 2). Similar results were obtained when we infected chickens with IBDV strain NN1172 (Table 3): the expression levels of chTLR3 at 1 and 2 dpi changed from 0.78- to 1.047-fold in the NN1172 group, while the expression level of IFN-β changed slightly (from 0.86- to 0.668-fold). These results were consistent with those of Eldaghayes et al. (7), whose results suggested that IBDV had downregulatory effects on the type I IFN responses, which would allow viral infection to be established. Interestingly, in chickens infected with intermediated virulence vaccine strain B87(in), both chTLR3 and IFN-β were downregulated at 1 dpi and then were upregulated after 2 dpi. We speculated that early activation of antiviral response may be interfered by IBDV to retain virus replication as well as the establishment of the acute phase of the disease. Meanwhile, another way of antiviral response from the host might be activated, which was why we detected the higher expression of IFN-β at 3 dpi as shown in Figure 2 and Table 3. Nevertheless, the replication ability of IBDV could no longer be restricted when infection is established, even a high titer of IFN-β was detected since a higher viral load was detected at later time point until the target cells were destroyed by the virus infection (as shown in Fig. 6). We speculated that cellular signal pathways might be interfered differently by IBDVs with different virulence, as shown in this study that different levels of expression of cytokines were observed between NN1172- and B87(in)-infected chickens.
IL-8 is a chemokine and one of the important downstream molecules in the TLR signaling pathway. In our study, when chTLR3 reached its highest expression level at 6 hpi in vitro and 3 dpi in vivo, IL-8 also significantly increased, showing that IL-8 responses correlated with IBDV infection. This response might be closely correlated with activation of the chTLR3 signaling pathway during IBDV infection. Most likely, IL-8 participated in the early stage of immune response after IBDV infection, which was in agreement with Chan's results when chicken was infected with avian influenza virus (6). However, further investigation is needed to confirm the exact correlation.
When IBDV strains with different virulence were used, we showed that chTLR3 expression levels in NN1172(vvIBDV)-infected PBMCs and CEFs were significantly higher than NN040124(intermediated plus virulent)- and B87(in)(intermediated virulent)-infected cells. In the in vivo experiment, we noted upregulated expression of TLR3 in the very virulent group (NN1172) at 3 dpi by 5.365-fold (p < 0.05). However, in the intermediated virulent vaccine strain (B87(in))-infected bursa, the TLR3 expression was significantly downregulated at 1 dpi by 0.23-fold (p < 0.05). This result demonstrated that NN1172-infected cells produced a higher level of chTLR3 in the bursa than the intermediated virulent IBDV (B87(in)) did. Consistently, Rauf et al. (33) also demonstrated a dramatic difference in bursal lesions, infiltration of T cells, and expression of cytokines, chemokines, and TLRs in the bursa of cIBDV- and avIBDV-infected chickens. All these results suggested that the more virulent the IBDV strain that was used for infection, the higher was the level of chTLR3 activation detected. Moreover, similar kinetics of expression was detected in IFN-β and IL-8 compared with that of chTLR3 when both in vitro and in vivo experiments were performed using IBDVs with different virulence. We showed that the expression of IFN-β in the bursa of B87(in)-infected group was downregulated at 1 dpi; however, IFN-β in NN1172- and B87(in)-infected groups were significantly increased at 3 dpi, but decreased at 5 and 7 dpi, especially significant decrease was detected in the NN1172 group at 7 dpi. The result suggested that enhancement of antiviral activities was activated at the early stage and then inhibited by the host after 3 dpi. This could be evidence that chTLR3 could be an efficient sensor to detect dsRNA from IBDV (32). Meanwhile, the expression of chemokine IL-8 in the bursa of very virulent strain NN1172-infected chickens was significantly upregulated at 3 dpi. At 1, 3, 5, and 7 dpi, the transcription of IL-8 in intermediated vaccine strain B87 (in)-infected group was significantly decreased compared with NN1172. Previously, similar upregulation of IL-8 expression was also reported by Guo et al. (12) in IBDV infection. This result suggested that IBDV strains were capable of activating proinflammatory cytokine and chemokine responses. Importantly, our data revealed that the more virulent the IBDV used for infection, the more robust was the stimulation of immune molecules detected.
So far, most studies on IBDV pathogenesis have been done in SPF layer chickens; fewer commercial chickens have been used for the investigation. Aricibasi et al. (4) documented that the genetic background of the chicken has significant impact on the disease outcome. Raj et al. (32) also reported that chTLR3 was differentially regulated by IBDV infection in commercial and indigenous chickens. The Three-Yellow chicken is one of the most popular meat-type local chicken breeds in southern China. The PBMCs, including B cell, NK cell, macrophage, and other immune cells, were reported to be susceptible to IBDV. CEF is a regular cell type for the infection of IBDV (8). In this study, commercial Three-Yellow chicken and its PBMCs and CEF cells were used as the host model for the infection of IBDV. We noted that after the PBMCs were infected with IBDV field strain NN040124, the virus amplified starting at 1.5 hpi, and after 3 hpi, viral genome copy number was significantly increased and reached the peak at 6 hpi. However, the viral copy number kept decreasing after that, but still maintained at much higher level till 48 hpi. These data suggested that IBDV could replicate in PBMCs in a time-dependent manner with speed. The viral load decreased after 6 hpi; we speculated that this might be the reason for cell apoptosis induced by IBDV infection. Apoptosis has an important role in IBDV pathogenesis and immunosuppression (29). Vasconcelos and Lam (38) had reported that a high level of apoptosis is evident in chicken peripheral blood cells infected with serotype I IBDV strain L at 6 hpi. To sum up, these results suggested that PBMC could serve as an ideal host candidate for IBDV pathogenesis' investigation.
In summary, the present study provides a basis in the activation of chTLR3 and its downstream effectors with IBDV infection both in vitro and in vivo from commercial Three-Yellow chickens. The results indicated that chTLR3 was activated by IBDV, resulting in expression of antiviral IFN-β and chemokine IL-8. Interestingly, the more virulent the IBDV strain that was used, the more efficient replication of the virus could be detected, the more pronounced was the expression of chTLR3, IFN-β, and IL-8. These results have clearly demonstrated the involvement of chTLR3 in the pathogenesis of IBDV in commercial Three-Yellow chickens, and in this process, the downstream effectors, IFN-β and IL-8, might play an important role.
Footnotes
Acknowledgments
This project was supported by the National Natural Science Foundation of China (31560706, 31660717), Guangxi Natural Science Foundation grants (2014GXNSFAA053060), and Xiangsihu Young Scholars Innovative Research Team of Guangxi University for Nationalities (2014).
Author Disclosure Statement
No competing financial interests exist.