
Abstract
To prepare the dominant multiepitope fusion antigen ROP2-SAG1 (RSmultiepitope) from Toxoplasma gondii in a prokaryotic system, the major immunodominant region (MIR) of the human hepatitis B virus core antigen (HBcAg(MIR)) was used as a delivery vector. The gene encoding the RSmultiepitope was inserted into HBcAg(MIR), and rHBcAg(MIR)-RSmultiepitope was prepared, purified, and administered to BALB/c mice through intradermal injection. An indirect enzyme-linked immunosorbent assay analysis based on a multiepitope peptide facilitated the specific differentiation of sera obtained from mice immunized with the rHBcAg(MIR)-RSmultiepitope protein, and high titers (greater than 1:6,400) of specific anti-RSmultiepitope antibodies were obtained. Immunized splenocytes demonstrated enhanced IFN-γ production. Based on these results, the HBcAg(MIR) vector is easily applied in vitro for targeting the RSmultiepitope and efficiently presents this target epitope for the induction of significant humoral and cellular immune responses. This study offers a novel strategy for the design of a target epitope delivery system for a toxoplasmosis vaccine.
Introduction
T
Due to the complex antigens present in T. gondii, the identification of a candidate antigen that induces immune protective effects and can be used for vaccine preparation has proved difficult. Major surface protein 1 (SAG1) and rhoptry protein 2 (ROP2) of T. gondii are considered among the two best potential candidate vaccine antigen molecules (9). In our previous work (11), we confirmed the immunological characteristics of the fusion protein ROP2-SAG1 both in vitro and in vivo and discovered the sequences responsible for potential pathological lesions in the host. In addition, given the complexity and diversity of the antigenic components of T. gondii, it is difficult to achieve a fully protective immune response and broad population coverage due to HLA restriction (3).
Accordingly, we predicted and screened certain immunodominant T and B cell epitopes from the fused antigen ROP2-SAG1 and reconstructed a combinatorial dominant multiepitope comprising dominant T and B cell epitopes. However, although prokaryotic codon optimization was performed, this multiepitope was not successfully expressed in a prokaryotic expression system. Therefore, the selection of a suitable antigen delivery vector for in vitro epitope expression is imperative.
The hepatitis B virus core antigen (HBcAg) has been developed as an effective tool for antigen presentation (2). In particular, the major immunodominant region (MIR) corresponding to amino acids 78–82 of HBcAg located on the spike tip of the virus (1,4) accepts the insertion of foreign epitopes and displays exogenous epitopes to induce strong humoral and cellular immune responses. Previous studies performed by our laboratory (10,18) identified MIR as the best fusion site for exogenous epitope insertion that also yields increased immunogenicity of the exogenous epitope. Other studies have also confirmed the generation of effective immune responses with fusion products containing the MIR and exogenous genes (5,16). Thus, MIR of the human hepatitis B virus core antigen (HBcAg(MIR)) appears to be a good epitope delivery vector.
In the present study, we inserted the ROP2-SAG1 multiepitope (RSmultiepitope) into HBcAg(MIR) to prepare recombinant HBcAg(MIR)-RSmultiepitope (rHBcAg(MIR)-RSmultiepitope) and evaluated the humoral and cellular immune responses induced by rHBcAg(MIR)-RSmultiepitope in mice. Our study presents a novel design strategy for the delivery of candidate antigen molecules that could be used as a toxoplasmosis epitope vaccine.
Materials and Methods
Restriction enzymes (SacI, BamHI) were purchased from MBI Fermentas (Burlington, Canada). Plasmid maxi preparation kits and Ni2+—chelated affinity columns were purchased from Qiagen (Hamburg, Germany). DNA Gel Extraction Kits and PCRDNA Extraction Kits were purchased from Generay Biotech (Shanghai, China). A Quick Spot™ IFN-γ ELISPOT Kit was purchased from Dakewe (Beijing, China). Horseradish peroxidase (HRP)-conjugated mouse anti-His monoclonal antibody (mAb) and HRP-conjugated goat anti-mouse IgG (H+L) were purchased from ABR, Inc.
The RSmultiepitope peptide and three T epitope peptides selected from the RSmultiepitope (peptide 1, RLRPGQPRLF [cytotoxic lymphocyte (CTL) epitope]; peptide 2, RNIELSEPVKFL [CTL epitope]; and peptide 3, TEWIFKRCRNIELSEPVKFL [CTL+Th epitope]) were synthesized separately by Hunchen Biotech, Co., Ltd. (Xi'an, China) and preserved in our laboratory at a purity of 98.9%. Purified HBcAg protein was prepared and preserved in our laboratory.
The Escherichia coli strain Rosetta was maintained in our laboratory. Six to 8-week-old female BALB/c mice (National Science Academy of Centre for Laboratory Animals, Shanghai, China) were used for the experimental procedures in accordance with protocols approved by the Wenzhou Medical College and the recommendations for the proper use and care of laboratory animals in China.
Determination of the RSmultiepitope
Based on the complex antigen ROP2-SAG1 (11), immunodominant T and B cell epitopes were predicted and screened using bioinformatics methods. Briefly, three immunodominant B cell epitopes were selected through comprehensive analyses of the transmembrane domain, hydrophilicity, surface probability, antigenicity index, polarity, and average flexibility. MHC-restricted T cell epitopes were predicted using the Internet service SYFPEITHI (
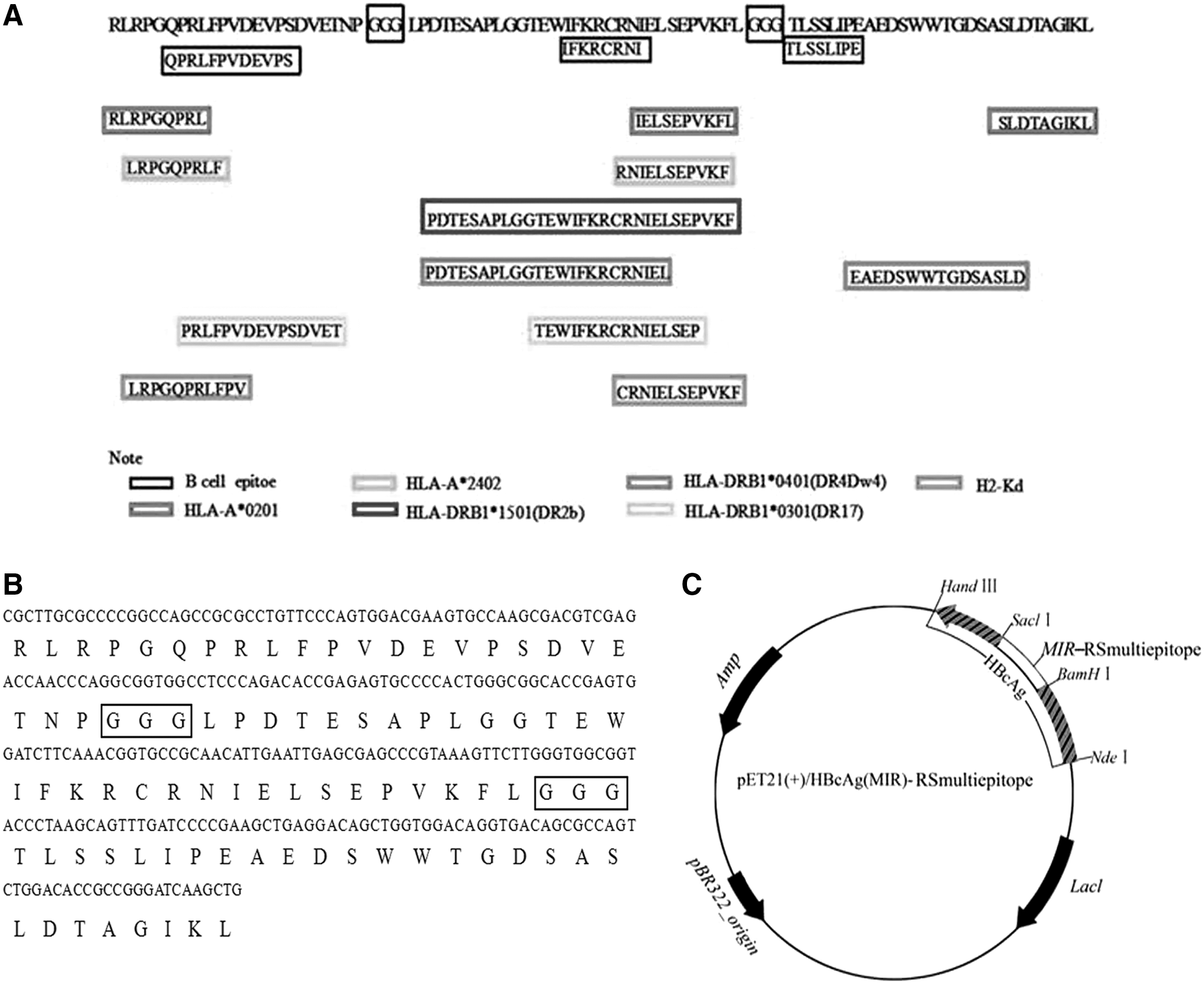
Epitope distribution and recombinant plasmid construction.
Construction of the pET21a(+)/HBcAg(MIR)-RSmultiepitope recombinant plasmid
The methods used to construct the pET21a(+)/HBcAg(MIR) plasmid have been previously described elsewhere and were used in this study with some modifications (14,15,19). In brief, BamHI and SacI restriction endonuclease sites were generated by synonymous mutations at amino acid sites from 74 to 82, which is in the region of the MIR. The mutant HBcAg gene with the universal Th epitope was synthesized by San bio-Tech (Beijing, China) and was then subcloned into the pET21a(+) vector with a 6-His tag. NedI and HindIII restriction endonuclease sites were introduced at the N and C terminals, respectively, of the HBcAg nucleic acids for purification and digestion (named pET21a(+)/HBcAg(MIR)). The recombinant plasmid was confirmed by polymerase chain reaction (PCR) and restriction endonuclease digestion. Then, pET32a(+)/RSmultiepitope (constructed and preserved in our laboratory) was designed with BamHI and SacI restriction enzymes, cloned into the pET21a(+)/HBcAg(MIR) vector, and digested with BamHI and SacI to generate the recombinant plasmid pET21a(+)/HBcAg(MIR)-RSmultiepitope. The construction map is shown in Figure 1C.
Expression and identification of rHBcAg(MIR)-RSmultiepitope in E. coli
After PCR confirmation of the recombinant plasmid, restriction endonuclease digestion, and sequencing, E. coli Rosetta strain cells were transformed with pET21a(+)/HBcAg(MIR)-RSmultiepitope, and pET21a(+) and pET21a(+)/HBcAg were used as vector controls. The transformation was followed by 16 h of shaking in Luria broth medium containing ampicillin (100 μg/mL) and overnight incubation at 37°C. After 4 h of induction with isopropyl-β-D-thiogalactopyranoside (Sigma) at a final concentration of 1 mM/L, the cells were centrifuged at 10,000 g for 5 min for collection. The bacteria were lysed through ultrasound for expression analysis. The expression products were purified through a Ni2+—chelated affinity column and confirmed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blot.
Immunization protocol
Six to 8-week-old BALB/c mice (H2-Kd) were randomly assigned (n = 5) to receive the rHBcAg(MIR)-RSmultiepitope protein, the RSmultiepitope peptide, HBcAg (the vector control), or phosphate-buffered saline (PBS; blank control). The immunization schedule involved three intradermal injections of 50 μg of purified protein or peptide at 2-week intervals. Sera were collected from the tail vein at weeks 0, 2, 4, 6, and 8 for the evaluation of antibody IgG production. The spleens were removed 1 week after the final immunization, and single-splenocyte suspensions were prepared for the detection of IFN-γ production.
Antibody detection by enzyme-linked immunosorbent assay
The production of RSmultiepitope-specific IgG in sera obtained from immunized mice was measured by enzyme-linked immunosorbent assay (ELISA). Microtiter 96-well flat-bottom plates (Corning, Lowell, MA) were coated overnight at 4°C with 100 μL (10 μg/mL) of the RSmultiepitope peptide, blocked for 1 h at 37°C with blocking buffer (5% nonfat dry milk and 0.05% Tween-20 in PBS), and subsequently incubated for another 1 h at 37°C with the serum samples (diluted 1:100). After washing with PBS, the plates were incubated with HRP-conjugated anti-mouse IgG diluted 1:3,000 in blocking buffer for 1 h at 37°C for detection of the bound antibodies. After washing with PBS-T, the chromogenic substrate
Immune sera obtained at week 4 were serially diluted twofold and used for the detection of the multiepitope-specific IgG titers according to the protocol described above. Each dilution was also run in triplicate.
Dot immunobinding assay
The specific binding characteristics of the immune sera were confirmed by dot immunobinding assay (DIBA). Four nitrocellulose membrane films were individually treated with 10 μL (1 μg/μL) of purified HBcAg, 10 μL (1 μg/μL) of purified rHBcAg(MIR)-RSmultiepitope, or 10 μL of PBS and incubated at 37°C for 30 min. The membranes were blocked with blocking buffer overnight at 4°C. After washing, the membranes were incubated with immune sera from mice immunized with the RSmultiepitope peptide, rHBcAg(MIR)-RSmultiepitope, HBcAg, or PBS for 2 h. All immune sera were diluted 1:3,000. After washing, HRP-conjugated anti-mouse IgG (diluted 1:3,000) was added for 2 h for assessment of the immune sera. Spots were visualized using a Chemiluminescence Detection Kit (Pierce, Rockford, IL) according to the manufacturer's instructions.
IFN-γ production assay
Spleen cells from immunized mice were hemolyzed in lysis buffer (0.83% NH4Cl and 0.01 M Tris-HCl, pH 7.2) for 1 min. After the cells were washed and adjusted to a concentration of 5 × 106 cells/mL, 100 μL of the cell suspension was placed into the wells of 48-well microplates precoated with an anti-IFN-γ mAb for coincubation with three T cell epitope peptides (peptide 1, 2, or 3) derived from the RSmultiepitope at a concentration of 25 μL/mL for 20 h. The subsequent steps were performed according to the instructions provided with the ELISPOT Kit. Finally, after coloration and rinsing, the enzyme-linked spots were counted using an automatic analyzer (Elispot reader 400 PRO-X; BioSys Bioreader®, Germany). All samples were assayed in triplicate.
Statistical analyses
The IgG levels are expressed as the mean measured values ± standard deviations (SD). All comparisons were tested for significance by one-way analysis of variance. Differences with p ≤ 0.05 were considered significant. Statistical analyses were performed using SPSS statistical software (SPSS, Inc., 1989–1999, Chicago, IL).
Results
Construction and expression analysis of rHBcAg(MIR)-RSmultiepitope
The constructed pET21a(+)/HBcAg(MIR)-RSmultiepitope vector was confirmed by PCR amplification and endonuclease digestion. A280-bp fragment of the RSmultiepitope was amplified (Fig. 2A, lane 3) and obtained through endonuclease digestion with SacI and BamHI in BamHI buffer (Fig. 2A, lane 2). A sequencing analysis revealed the presence of the target RSmultiepitope gene in the plasmid (data not shown), and the results indicated the successful insertion of the RSmultiepitope into pET21a(+)/HBcAg(MIR).

Plasmid identification and protein expression analysis.
A 29-kDa rHBcAg(MIR)-RSmultiepitope protein was expressed in E. coli (Fig. 2B, lane 2), and higher expression was observed in the supernatants of cellular lysates obtained by sonication (Fig. 2B, lane 3). Purified rHBcAg(MIR)-RSmultiepitope protein was obtained (Fig. 2B, lane 4) at a concentration of 300 μg/mL and was recognized by a His-tag mAb (Fig. 2C). These results indicate that HBcAg(MIR) carrying the RSmultiepitope was expressed in a prokaryotic expression system.
Antibody production is greatly enhanced by the incorporation of the RSmultiepitope into rHBcAg(MIR)
Two weeks after immunization, the RSmultiepitope-specific antibody levels in both the RSmultiepitope peptide-immunized and rHBcAg(MIR)-RSmultiepitope-immunized groups of mice began to increase significantly. In addition, the antibody levels in the rHBcAg(MIR)-RSmultiepitope group appeared to increase over time; in fact, the antibody levels at week 4 were significantly different from those at week 2 (p < 0.05), and the levels at week 6 were significantly different from those at week 4 (p < 0.05). The analysis of the RSmultiepitope peptide group revealed that the antibody levels at week 4 were significantly different from those at week 2 (p < 0.05), but no significant difference in the antibody levels was found between weeks 4 and 6 (p > 0.05) (Fig. 3A). At week 6, the maximum serum dilution for specific antibodies in the rHBcAg(MIR)-RSmultiepitope group was 1:6,400 (Fig. 3B), whereas the maximum dilution for the RSmultiepitope peptide group was 1:3,200 (Fig. 3B). These results demonstrate that HBcAg(MIR) can serve as a target epitope-carrying vector capable of greatly improving specific antibody production in vivo.
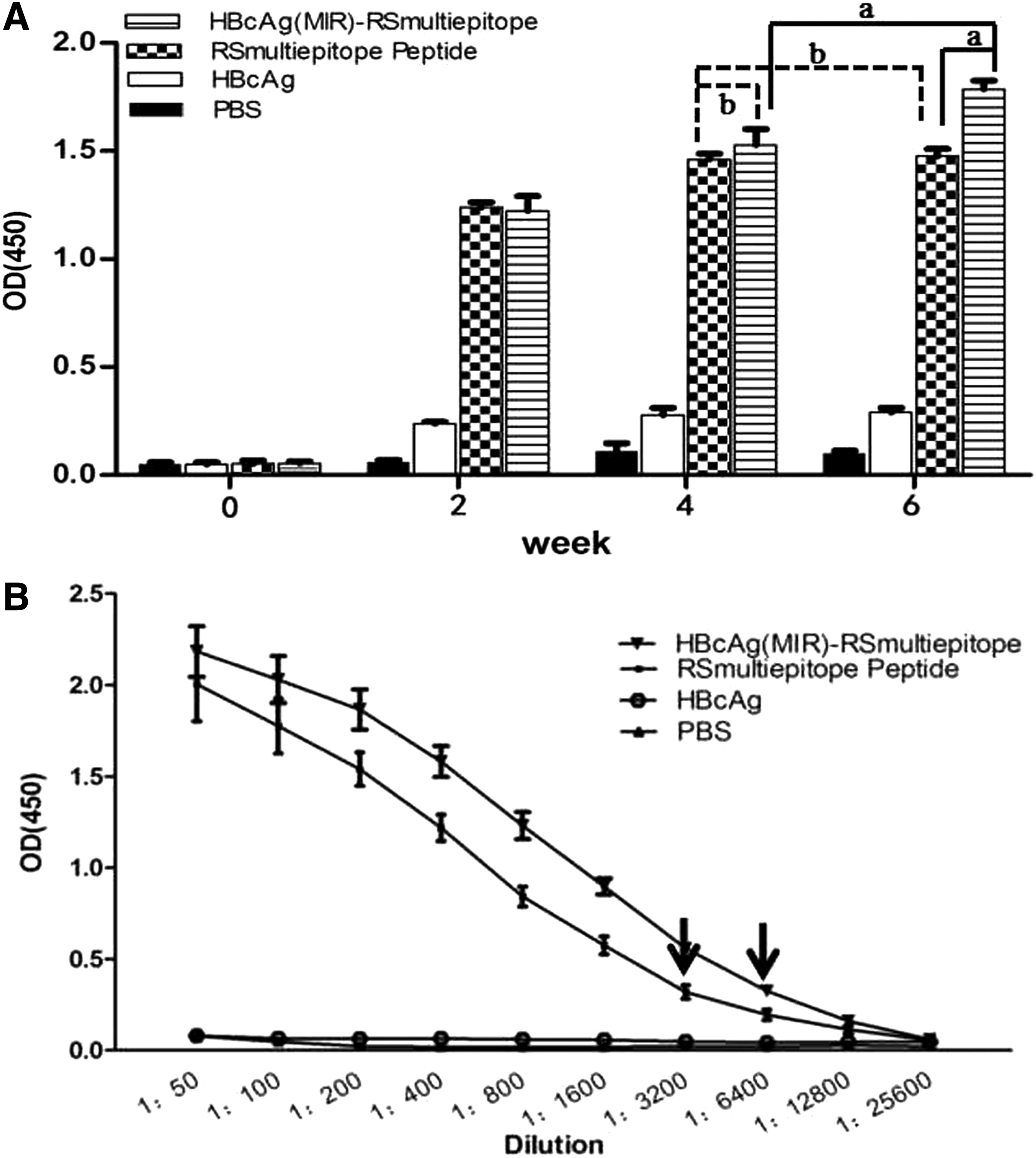
Detection of RSmultiepitope-specific antibodies by ELISA.
Dot immunobinding assay
The analysis of nitrocellulose films conjugated with PBS, HBcAg, or rHBcAg(MIR)-RSmultiepitope showed no combined dots when blotted with immune sera from the PBS group; however, when blotted with immune sera from the HBcAg group, combined dots were clearly observed at the locations corresponding to HBcAg and rHBcAg(MIR)-RSmultiepitope. When blotted with immune sera from the RSmultiepitope peptide group, combined dots were only observed at the location corresponding to rHBcAg(MIR)-RSmultiepitope, and blotting with immune sera from the rHBcAg(MIR)-RSmultiepitope group yielded combined dots at the locations corresponding to HBcAg and rHBcAg(MIR)-RSmultiepitope (Fig. 4). These results further demonstrate the specificity of immune sera from the fusion expression of HBcAg and the RSmultiepitope for the detection of both HBcAg and the RSmultiepitope.

Confirmation of antibody specificity according to DIBA. DIBA, dot immunobinding assay.
IFN-γ production in immune splenocytes
When stimulated with concanavalin A (a nonspecific mitogen), splenocytes from all groups of immunized mice produced high levels of IFN-γ, confirming the common murine lymphoid cell response (Fig. 5A). Splenocytes from the rHBcAg(MIR)-RSmultiepitope group produced higher numbers of spots corresponding to IFN-γ compared with those obtained from the RSmultiepitope peptide group stimulated with any type of peptide (peptide 1, 2, or 3) (Fig. 5A); the corresponding IFN-γ spot counts are shown in Figure 5B. In addition, stimulation with peptide 3 (CTL+Th epitope) resulted in improved IFN-γ production compared with stimulation by individual CTL epitope peptides (peptide 2 or 1; p < 0.05 for both), whereas no significant difference in the stimulating capacities of peptides 2 and 1 was detected (p > 0.05), indicating that the costimulatory effects of CTL in combination with the Th epitope peptide led to immunocyte activation.
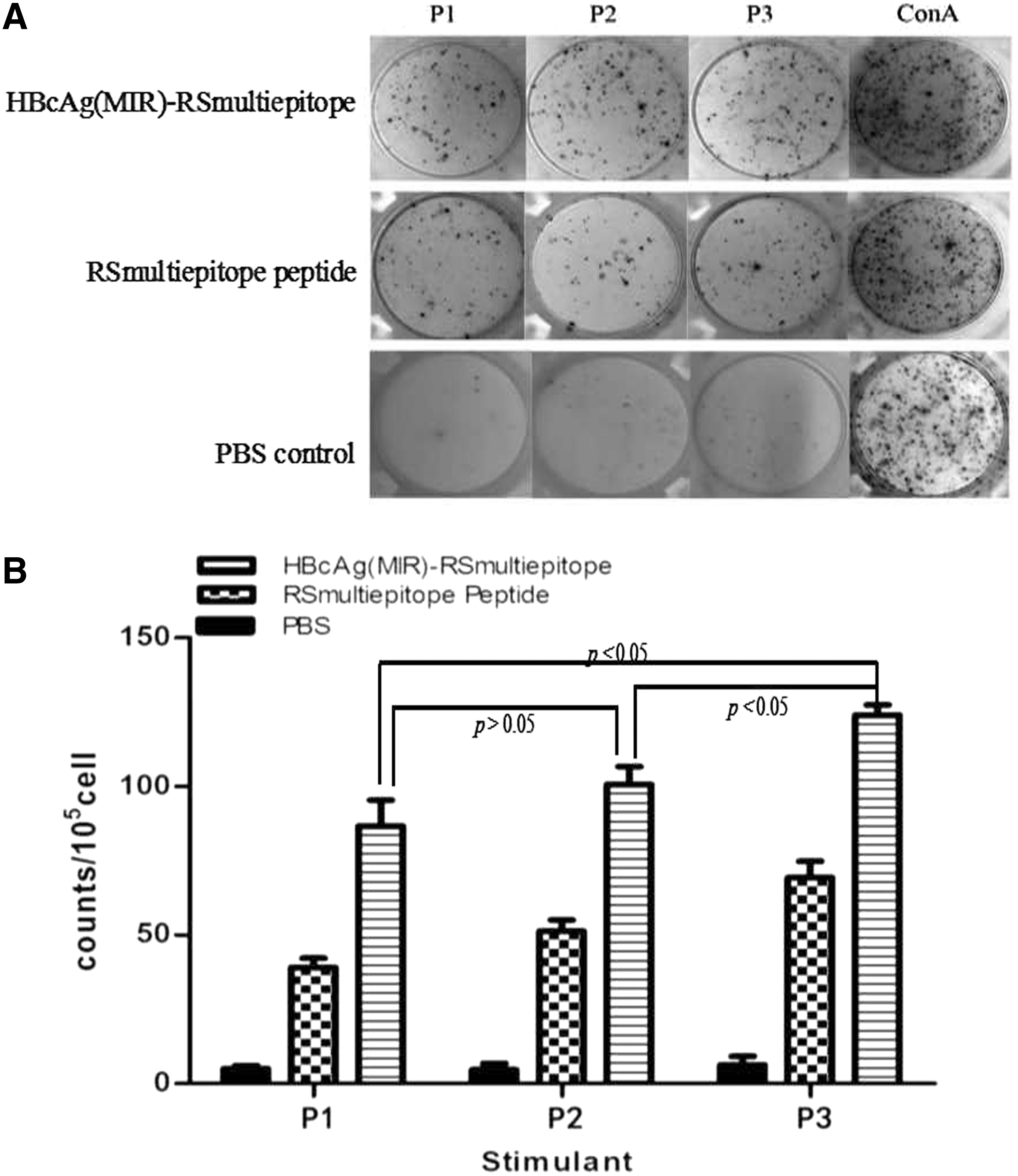
IFN-γ production by immunized splenocytes.
Discussion
An epitope is a small segment in an antigen molecule that is responsible for inducing a specific immune response. However, because immunosuppressive or immunopathological epitopes should not yield any interference, it is very important to select immunodominant epitopes that induce an effective immune response. Studies developing vaccines to combat malignant tumors and infectious diseases are now paying greater attention to epitope vaccines (6 –8), which present clear immune advantages in terms of their ability to not only induce specific immune responses but also avoid immune pathological impairments.
The antigen components of T. gondii are complex, and the parasite infection process involves a variety of antigens. SAG1 and ROP2, two major key antigens of T. gondii infection, are considered ideal target antigens for vaccine development. We predicted, screened, and determined dominant T and B cell epitopes and linked these dominant epitopes together to form an 88-amino-acid “artificial dominant epitope combination”; however, the expected dominant epitope protein could not be generated in vitro. Thus, the selection of an effective vehicle for delivering and releasing a target epitope is essential.
Our research team has focused on the development of dominant epitope vectors for many years and have constructed viral vectors in which capsid proteins act as vehicles and are optimized for coexpression with inserted exogenous genes. Such vectors include human papilloma virus (HPV) (12), hepatitis B virus surface antigen (HBsAg) (21), and HBcAg (20). Among these, HBcAg can carry and display foreign genes and expresses an ∼21-kDa capsid protein in different expression systems (e.g., E. coli, yeast, vaccinia virus, and insect cells). Its MIR, which is located on the tip of the surface spike, fuses with a foreign gene to display high-density repeat exogenous sequences and induces strong specific cellular and humoral immune responses. Therefore, HBcAg(MIR) is believed to be suitable as an epitope delivery and release system.
In this study, the RSmultiepitope was cloned into the vector pET21a/HBcAg(MIR), which was prepared in our laboratory, to successfully construct the recombinant plasmid pET21a/HBcAg(MIR)-RSmultiepitope, which expressed a 29-kDa fusion protein (HBcAg(MIR), 19 kDa; RSmultiepitope, 10 kDa) in E. coli. rHBcAg(MIR)-RSmultiepitope was recognized by a His-tag mAb, further confirming the expression of the fusion protein. Two weeks after immunization with the purified HBcAg(MIR)-RSmultiepitope protein or the RSmultiepitope peptide, RSmultiepitope-specific IgG antibodies were detected in the sera of immunized mice, and the RSmultiepitope-specific antibody levels increased over time. In particular, the mice immunized with HBcAg(MIR)-RSmultiepitope showed steady increases in antibody production, and the highest immune serum dilution for specific antibody detection reached 1:6,400, confirming the effective immunogenicity of the RSmultiepitope and the advantages of HBcAg as a good carrier.
Thus, in this study, HBcAg was selected as an exogenous gene carrier to not only achieve the expression of both the vector and the target gene, which might facilitate the effective in vitro expression of the epitope molecule, but also enhance antibody production against the target epitope. A DIBA also demonstrated the bispecificity of the immune sera from the rHBcAg(MIR)-RSmultiepitope group to both HBcAg and the RSmultiepitope. The stimulation of splenic lymphocytes from mice immunized with the rHBcAg(MIR)-RSmultiepitope protein with T cell dominant epitope peptides (peptide 1, 2, or 3) from the RSmultiepitope greatly improved the IFN-γ production levels compared with those obtained with the RSmultiepitope peptide group. These results indicated the significant immunological effects exerted by T cell epitopes found in the RSmultiepitope and revealed the dominant effects of HBcAg as a vector in enhancing cellular immune responses.
In conclusion, our findings suggest that HBcAg(MIR) is a superior carrier for exogenous epitope delivery and release in terms of not only the expression of foreign epitopes in vitro but also its ability to induce epitope-specific immune responses in vivo. This vector could be developed as a presentation platform for the additional insertion of other foreign epitopes with the aim of conferring good immunological properties.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (No. 81371668).
Author Disclosure Statement
No competing financial interests exist.