
Abstract
Highly pathogenic avian influenza (HPAI) H5N1 virus emerged in 1997 as a zoonotic disease in Hong Kong. It has since spread to Asia and Europe and is a serious threat to both the poultry industry and human health. For effective surveillance and possible prevention/control of HPAI H5N1 viruses, it is necessary to understand the molecular mechanism underlying HPAI H5N1 pathogenesis. The hemagglutinin (HA) protein of influenza A viruses (IAVs) is one of the major determinants of host adaptation, transmissibility, and viral virulence. The main function of the HA protein is to facilitate viral entry and viral genome release within host cells before infection. To achieve viral infection, IAVs belonging to different subtypes or strains induce viral–cell membrane fusion at different endosomal pH levels after internalization through endocytosis. However, host-specific endosomal pH also affects induction of membrane fusion followed by infection. The HA protein of HPAI H5N1 has a higher pH threshold for membrane fusion than the HA protein of classical avian influenza viruses. Although this particular property of HA (which governs viral infection) is prone to deactivation in the avian intestine or in an ambient environment, it facilitates efficient infection of host cells, resulting in a broad host tropism, regardless of the pH in the host endosome. Accumulated knowledge, together with further research, about the HA-governed mechanism underlying HPAI H5N1 virulence (i.e., receptor tropism and pH-dependent viral–cell membrane fusion) will be helpful for developing effective surveillance strategies and for prevention/control of HPAI H5N1 infection.
Introduction
I
Among LPAIVs, viruses of the H5 and H7 subtypes can sometimes give rise to highly pathogenic avian influenza viruses (HPAIVs), which infect poultry (particularly chicken) with high mortality. HPAI H5 viruses have caused occasional outbreaks in poultry worldwide (84); however, since the 1997 Hong Kong outbreak, HPAI H5N1 viruses have proved to be pathogenic not only to birds but also to humans. HPAI H5N1 caused an epizootic disease in poultry in 1996 in the southern China area, which was followed by a zoonotic disease in Hong Kong in 1997 that resulted in 18 human cases (6 of whom died) (12,82). HPAI H5N1 (in this review, the term “HPAI H5N1” refers to the virus that has adapted and shown strong virulence in humans since 1997) continued to circulate in China after 1997, re-emerging in 2003; since then, HPAI H5N1 strains have spread to Asia, Europe, and Africa (8,50,97), and many different HA clades have been identified as a serious threat to both the poultry industry and human health (17,99). Therefore, it is necessary to understand the molecular mechanism(s) underlying HPAI H5N1 pathogenesis if we are to provide effective surveillance and possible prevention/control measures for HPAI H5N1 viruses.
Although host adaptation, transmissibility, and viral virulence are multifactorial processes governed by each gene expressed by IAVs, the HA protein is one of the major determinants (23,27,46,79,100). HA proteins have a trimeric structure, and each individual monomer has to be cleaved into HA1 and HA2 subunits before activation. Usually, IAVs require host serine proteases such as the trypsin-like protease tryptase Clara (42) to cleave HA0 into HA1 and HA2; this occurs through enzymatic recognition of a conserved Q/E-X-R motif at the HA cleavage site (10). Cleavage is also catalyzed by other serine proteases such as transmembrane protease serine 2 (TMPRSS2), human airway trypsin-like protease (4), miniplasmin (56), and anionic trypsin I (88). Some H5 and H7 strains have acquired insertional mutations at the HA cleavage site (i.e., R-X-R/K-R) (45,80), which are recognized by the ubiquitous protease furin and by proprotein convertases (1,18,32,70,80,85,95). This multibasic site allows viruses to activate ubiquitous intercellular cleavage of HA (from HA0 to HA1 and HA2) and systemic replication, which is why they are named HPAIVs. A recent report suggests that other serine proteases such as mosaic serine protease large form and its splice variant transmembrane protease serine 13 (TMPRSS13), which are also expressed ubiquitously (41), cleave both types of HA peptide at R/K-K-K-R (although furin cannot cleave the K-K-K-R motif, which is conserved in some HPAI H5 and H7 strains) (61). Therefore, the multibasic amino acid sequence within HA endows avian influenza viruses (AIVs) with high pathogenicity, particularly in poultry, although it is unclear to what extent this factor contributes to virulence in mammalian hosts (including humans).
The main function of the HA protein is to facilitate viral entry and viral genome release within host cells (51,109). During viral entry, influenza virus HA recognizes terminal sialic acid (SA) and galactose residues on sialylglycans expressed by host cells and binds to them before entry. Seasonal influenza viruses preferentially recognize receptors harboring terminal α2,6-linked SA (α2,6SA) residues, which are abundantly expressed by cells lining the human upper respiratory tract (although some studies imply that α2,3SA also resides in the human upper respiratory region) (52,94); by contrast, AIVs preferentially bind to receptors harboring terminal α2,3SA residues expressed mainly by respiratory and enteric cells in birds (34). After binding to a receptor, the virus is internalized and the acidic pH in the endosomal compartment induces a conformational change in the HA protein, leading to fusion of the viral and endosomal membranes, thereby allowing release of viral RNA into the cell cytoplasm.
As already mentioned, HPAI H5N1 viruses infect birds that express α2,3SA on respiratory and enteric cells; however, these viruses can also infect cells lining the human lower respiratory tract (e.g., bronchioles and alveoli) that express both α2,3SA and α2,6SA residues (74,93). This is one possible mechanism(s) by which HPAI H5N1 infects humans. Thus, HPAI H5N1 strains that have mutated to show dual tropism (both avian and human receptor recognition) are more likely to infect humans than viruses that recognize only the avian receptor because they can attach to cells lining both the upper and lower respiratory airways (105). However, virus strains isolated from patients do not always show dual tropism. In addition, many classical AIVs (in this review, classical AIV refers to viruses that only circulate in birds with or without the multibasic amino acid sequence within HA) that preferentially recognize α2,3SA rarely infect the lower respiratory region in humans, despite its abundant expression of α2,3SA and α2,6SA. Therefore, HPAI H5N1 viruses might use an alternative mechanism to overcome species barriers to infect humans. Here, we focus on the HA-mediated membrane fusion step of the HPAI H5N1 viral infection process and summarize current knowledge about the biological aspects of HPAI H5N1 acid stability, which plays different roles in host cells and in ambient environments.
HA-Mediated Membrane Fusion Mechanisms
After virus particles enter host cells through endocytosis, the low pH in the endosomal compartment induces irreversible changes in the conformation of the HA protein that culminate in insertion of the HA fusion domain into the endosomal membrane to initiate viral–cell membrane fusion (Fig. 1). This membrane fusion step is followed by release of the viral genome into the host cell cytoplasm, which triggers viral infection. The HA proteins from different subtypes and strains vary in terms of the pH at which membrane fusion is triggered (7,13,14,22,38,62,64 –66,69,112). Based on our own recent studies and those of others, the pH thresholds for HA-mediated membrane fusion are as follows: classical AIV (H1–H15), 4.8–6.2; HPAIV H5N1, 5.3–6.0; and seasonal influenza virus (H1, H2, and H3), 5.0–5.7 (7,13,14,22,38,62,64 –66,112). However, care should be exercised when comparing the results of these studies because the population sizes employed differ between studies. In addition, Okamatsu et al. (60) determined the pH threshold for 50 LPAIVs (H1–H15) and 5 HPAI H5N1 viruses. Based on the values calculated using the parameters published in their report, the mean and median pH thresholds for the 50 LPAIVs (H1–H15) are 5.34 ± 0.23 (mean ± standard deviation [SD]) and 5.30, respectively, whereas those for the 5 HPAI H5N1 viruses are 5.63 ± 0.08 (mean ± SD) and 5.60, respectively. The influenza virus lifecycle is outlined in Figure 1. In brief, viral HA proteins with a pH threshold higher than the pH of the host cell endosome show strong infectivity because membrane fusion of the acid-destabilized HA protein is more readily triggered. However, the endosomal pH could also affect the efficiency of fusion and the resulting viral infectivity. A recent report by Murakami et al. (57) suggests that a proper balance between endosomal pH and the pH threshold for HA-mediated fusion is required for efficient virus replication in host cells. Recently, we showed that the HA protein of HPAI H5N1 viruses belonging to different clades (A/Thailand/Kan353/04 [clade 1], A/Indonesia/5/05 [clade 2.1.3], A/chicken/Egypt/CL6/07 [clade 2.2.1], A/Shanghai/1/06 [clade 2.3.4], and A/crow/Kyoto/53/04 [Cw/Ky; clade 2.5]) has a higher pH threshold for viral–cell membrane fusion than for classical AIVs, which are avirulent in humans (13). We found that Cw/Ky (H5N1; clade 2.5), which has a relatively high pH threshold (5.625) for HA fusion, infects almost all populations of human small airway epithelial cells (SAECs) lining the bronchioles, whereas a classical AIV, A/duck/Hong Kong/820/80 (Dk/Hk; H5N3), which has a low pH threshold (5.375), infects only some populations of SAECs (Fig. 2) (13). This agrees with the findings of recent reports by Galloway et al. (22) and Byrd-Leotis et al. (7), showing that the pH threshold for the HPAI H5N1 virus A/Viet Nam/1204/2004 (H5N1; 5.6–5.8) is higher than that for the classical H5 AI A/tern/South Africa/1961 (H5N3; 5.4) (here they report only the pH thresholds for HA-mediated fusion and present no data regarding infectivity). Our results also agree with those of Okamatsu et al. (60). Based on the values calculated using the parameters published in their report, the mean and median pH thresholds for five HPAI H5N1 viruses (5.63 ± 0.08 [mean ± SD] and 5.60 [median]) are higher than those for 50 LPAIVs (H1–H15; 5.34 ± 0.23 [mean ± SD] and 5.30 [median]). Thus, the biological characteristics of HPAI H5N1 HA proteins with a higher threshold for fusion may be one factor that contributes to the pathogenicity and/or broad tropism (for both birds and humans) of the HPAI H5N1 virus.

Lifecycle of influenza A viruses. Virus entry is initiated by binding to host receptors (sialic acid) expressed by target cells. After the virus binds to the receptor, virions are internalized through endocytosis. The endosomal pH gradually decreased during transition from the early endosome (pH 5.9–6.5) to the late endosome (5.0–6.0) (53,91,106). The reduced pH in the endosome induces a conformational change in the HA protein that triggers viral–endosomal membrane fusion, leading to release of the segmented viral genome into the host cell cytoplasm. HA, hemagglutinin.

Expression of viral antigens by primary human SAECs infected with AIVs. Primary human SAECs were infected with the HPAI H5N1 virus Cw/Ky (H5N1) or classical AIV Dk/Hk (H5N3) at a multiplicity of infection of 10. Viral antigens were detected by immunostaining at 16 h postinfection (green). Cell nuclei were counterstained with Hoechst 33342 (blue). Scale bars = 100 μm. AIV, avian influenza virus; HPAI, highly pathogenic avian influenza; SAECs, small airway epithelial cells. This research was originally published in Daidoji et al. (13) © American Society for Biochemistry and Molecular Biology.
Our results also agree with those of other reports describing the results of experiments in chicken (14,33) and mallard (64) and partially support the results of a recent report (112) in a mammal. Based on the reports by Hulse et al. (33) and DuBois et al. (14), A/chicken/Hong Kong/YU562/2001 (H5N1; chicken-virulent strain) showing fusion activity at pH 5.7 resulted in a lower 50% chicken lethal dose (LD50) than that of A/goose/Hong Kong/437-10/1999 (H5N1; moderately chicken-virulent strain), showing fusion activity at pH 5.3. Similarly, in the report by Reed et al. (64), A/chicken/Vietnam/C58/04 (H5N1) showing fusion activity at pH 5.9 or its mutant virus (HA1-H24Q) showing fusion activity at pH 5.6 exhibited higher viral shedding from trachea and cloaca of infected mallard and higher mortality than the mutant virus (HA2-K58I) showing fusion activity at pH 5.4. Also, in the report by Zaraket et al. (112), A/Vietnam/1203/2004 H5N1 showing membrane fusion at pH 6.0 had a higher virus titer and resulted in more weight loss in ferret than its mutant virus with HA2-K58I showing fusion activity at pH 5.5. However, in their report, the mutant virus (HA2-K58I) replicated more efficiently in the nasal cavity of infected ferret than the wild type virus. This phenomenon may be related to the pH environment of the human nasal cavity. The pH value of human nasal mucosa is relatively acidic (19,98). In the acidic mammalian nasal cavity, AIVs with acid-stable HA might have an advantage because they might be able to engage in more efficient replication than AIVs with acid-destabilized HA. In contrast, in the lower respiratory regions such as the lung, AIVs with acid-destabilized HA might replicate more efficiently, because the pH of the mammalian alveolar region is less acidic than that of the nasal cavity (59). Therefore, in the case of AIVs with a high pH threshold, which is the case for many H5N1 strains, a large amount of virus entering the mammalian host might be able to bypass the barrier of the acidic nasal mucosa and reach the lower respiratory region, where survived intact viruses could engage in efficient viral replication in the mammalian lung. In contrast, in the case of AIVs with a low pH threshold, such as classical avian strains, although such viruses would be able to bypass the acidic nasal mucosa as intact virions and enter the lower respiratory tract, it would be difficult for them to replicate there because classical AIVs only partially infect lower respiratory epithelial cells, which consist of a variety of cells with distinct endosomal pH levels, as reported in our recent article (13). Nonetheless, a recent report (113) has shown that A/chicken/Vietnam/C58/04 (H5N1; fusion at pH 5.9) shows lower replication in mouse lung than the mutant virus (HA2-K58I; fusion at pH 5.4), which is inconsistent with our notion and the recent report describing results obtained in ferret (112). Therefore, further studies are warranted to reveal the relationship between acid stability of AIVs and its virulence in mammals, although it is possible that the input dose of AIVs with a high pH threshold in mammalian hosts is the most important factor affecting viral virulence, because a higher amount of virus particles would be able to reach the lower respiratory tract as intact virions and result in efficient replication. In contrast, the effective pH range of HA-mediated membrane fusion seems to be limited, although AIVs with a higher pH threshold of HA fusion, such as H5N1, replicate in avian and mammalian hosts, as was observed in recent reports demonstrating that the highly elevated pH threshold of the H5N1 HA mutant (fusion at pH 6.3) results in the attenuation of virus propagation in both avian (64) and mammalian hosts (113).
We also found that primary human airway epithelial cells mainly comprise two different populations, one with a high and one with a low endosomal pH environment (13). We generated cell lines derived from SAECs lining the bronchioles by induction with SV40 large T antigen (denoted here as SAEC-T cells); these SAEC-T clones could be categorized into two groups, each with distinct endosomal pH levels (low and high). Interestingly, these two groups of cells were highly and poorly susceptible to infection by Dk/Hk (H5N3) virus, respectively, whereas both were highly susceptible to infection by Cw/Ky (H5N1) (Fig. 3). We confirmed that the two groups of cells expressed similar patterns of α2,3 SA and α2,6 SA receptors on the surface with similar epithelial cell morphology. To investigate further this cell susceptibility, we made a recombinant Dk/Hk (H5N3) expressing the multiple basic amino acid sequence from H5N1 within its HA (rDk/Hk-RRKKR-HA) and used it to infect both highly (named 1A5 cells) and poorly (named 21E5 cells) susceptible cell clones with low and high pH values, respectively. The results showed no difference in viral antigen expression pattern in 1A5 and 21E5 cells between Dk/Hk (H5N3) and rDk/Hk-RRKKR-HA. In addition, between Cw/Ky (H5N1) and rDk/Hk-RRKKR-HA viruses, there was no significant difference in the number of virions bound to the surface of 1A5 or 21E5 cells. Also, we confirmed that rDk/Hk-RRKKR-HA showed similar internalization kinetics to Cw/Ky (H5N1) (13). These results suggest that differences in susceptibility to infection by classical AIVs are because of differences in endosomal pH levels in SAEC-Ts, and that the infectivity of AIVs depends, at least in part, on an exquisite balance between endosomal pH and the pH threshold for fusogenic activation for each viral strain (Fig. 3). To confirm this, we changed the endosomal pH of the two different cell clones with distinct endosomal pH levels (low pH: 1A5 cells; high pH: 21E5 cells) using the specific reagent BafA1, which increases endosomal pH through its inhibition of vacuolar-type H(+)-ATPase activity. BafA1 prevented endosomal acidification and inhibited viral–cell membrane fusion and resultant viral infection (63). Although BafA1 treatment (minimum concentration, 1.56 nM) of 1A5 cells remarkably inhibited Dk/Hk (H5N3) replication, BafA1 treatment did not inhibit the growth of Cw/Ky (H5N1) in 1A5 cells, even at 3.125 nM. Although higher doses of BafA1 (≥6.25 nM) prominently inhibited the growth of Cw/Ky (H5N1) in 1A5 cells, lower doses of BafA1 (1.56 nM) inhibited the growth of Cw/Ky (H5N1) in 21E5 cells (Fig. 4). Dk/Hk (H5N3) did not propagate well in 21E5 cells in the presence or absence of BafA1 (Fig. 4). These results strongly support our assumption that the pH environment of the endosomes in SAEC-Ts plays a role in determination of susceptibility to infection by H5N1 and other AIVs.

Mechanism of infection in two SAEC-T clones with different endosomal environments.
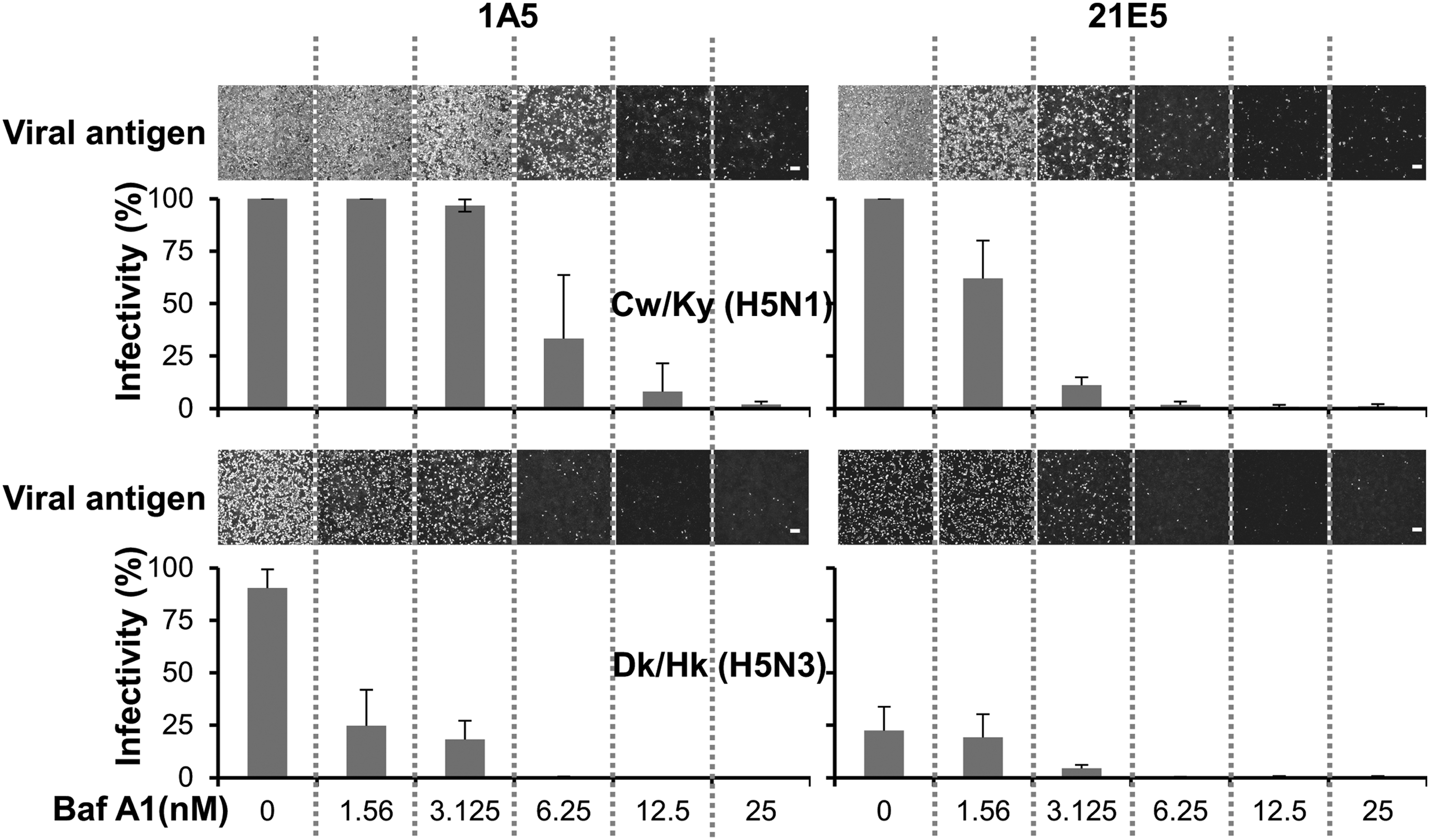
BafA1 inhibits virus infectivity. High (1A5) and low (21E5) susceptibility clones were treated with BafA1 (0–25 nM) for 2 h and then simultaneously infected with Cw/Ky (H5N1) or Dk/Hk (H5N3) at a multiplicity of infection of 10. Twelve hours later, infectivity was determined by calculating the percentage of antigen-positive SAEC-T cells. Representative micrographs of antigen-positive cells treated with BafA1 are shown. Cell nuclei were also counted (not shown as micrographs). The percentage of cells (for each SAEC-T clone) infected by the virus (the infectivity ratio) was calculated by dividing the number of antigen-positive cells by the total number of nuclei in the same field. Data are expressed as mean ± standard deviation of three independent experiments. Scale bars = 200 μm. This research was originally published in Daidoji et al. (13) © American Society for Biochemistry and Molecular Biology.
The acidity of the early and late endosome is species/tissue specific (Table 1) (43,67,75,76,90 –92,106,107,109,114). Thus, a low pH in the late endosomes of Madin–Darby canine kidney (MDCK) cells (pH 5.1–5.2) could explain the fact that, to date, MDCK cells are the most sensitive and reliable cell line for isolating and propagating IAVs (24,26,54,68). As already described, we found that human airway epithelial cells comprise two main populations, each with distinct endosomal pH levels (low and high) and that these cells show differing susceptibility to infection by AIV. The physiological heterogeneity of airway epithelial cells may arise from differences in either cell origin or differentiation stage. Club cells (originally known as Clara cells) and basal/parabasal cells (stem cells) lining the airways play important roles in the renewal of ciliated cells within the epithelium (2,3,15). Thus, rates of cellular turnover or the number of cells descended from a particular origin could result in differences in endosomal pH levels and differing susceptibility to viral infection. Furthermore, based on the fact that cell lines or primary cells from different species and/or organs have different endosomal pH levels (Table 1), it is possible that endosomal pH levels vary between the upper and lower respiratory tracts. However, further studies are required to determine endosomal pH levels in populations of epithelial cells lining the lower and upper airway tract in humans. It remains unclear why the human airway epithelium comprises physiologically different types of cells with high and low pH endosomal environments; it may be that such differences in endosomal pH protect tissues against some pathogens, including enveloped viruses and toxins that can be activated by acidic environments. Nonetheless, HPAI H5N1 viruses harboring HA proteins with a higher pH threshold for activation may overcome such putative host strategies and efficiently infect human airway epithelial cells.
Data are from the late endosome/lysosome in the original article.
CEF, chicken embryo fibroblasts; CHO, Chinese hamster ovary cells; MDCK cells, Madin–Darby canine kidney cells.
Acid Stability of HPAIV H5N1 HA and Its Biological Role in Both Ambient and Host Cell Environments
Most AIVs replicate in the intestinal tissue of waterfowl (31,40,44,101). The intestinal contents of the avian intestine are relatively acidic, although there are slight differences between species or organs/tissues (Table 2) (5,16,29,48). Thus, the balance between intestinal pH and viral acid stability is essential for persistence of classical AIVs. Classical AIVs with a lower pH threshold for fusion are acid stable, which allows them to reside within the avian intestinal tract; however, HPAI H5N1 viruses with a higher pH threshold may be inactivated because of their lower acid stability (7,13,14,22,60,64,65,112). Compared with that of other birds, the duck intestine is mildly acidic (the small intestine of the turkey is also mildly acidic) (16), which presumably enables waterfowl to act as a reservoir for most AIV subtypes (30,35,72,73,77,78,83,96). By contrast, HPAI H5N1 viruses prefer to replicate in the avian respiratory tract; indeed, oropharyngeal swabs from wild ducks contain higher virus titers than cloacal swabs (6,39). Preferences for particular replication conditions may be illustrated by the fact that many subtypes of classical AIVs (with the exception of HPAI H5N1 viruses) are found in surface water in lakes (or ponds), possibly because waterfowl excrete virus in feces, whereas HPAI H5N1 viruses are scarcely found in natural environments (30,35). The HPAI H5N1 virus is likely to prefer the nasal/oral route of transmission rather than the fecal–water–oral pathway in which the virus may be exposed to aquatic pH and/or intestinal pH environments. Although the HPAI H5N1 virus, which has an acid-sensitive HA protein, is prone to deactivation in natural environments, a higher pH threshold for membrane fusion could provide a biological benefit and allow efficient viral infection of host birds. This assumption is supported by data showing that pathogenic HPAI H5N1 virus has been isolated from a wide range of wild (e.g., mallard, goose, gull, swan, pigeon, sparrow, and crow) and domestic (duck, chicken, turkey, and quail) birds and causes occasional but severe disease in humans through direct transmission from birds; this is despite the fact that infection also depends on other factors such as multibasic cleavage sites within HA and steps in the virus lifecycle that occur after membrane fusion (Fig. 1). The polymerase activity of IAV is one of the major factors, together with the HA function, that affects virulence. Amino acid mutations within PB2 such as E627K and D701N help AIVs adapt to a mammalian host (21,27,28,47,55,104). Further studies should examine the correlation between HPAI H5N1 virulence and factors such as HA-mediated membrane fusion, multibasic cleavage sites within HA, and mammalian-adapted polymerase activity.
These parameters are taken from Bowen and Waldroup (5), Farner (16), Hewitt and Schelkopf (29), and Lin et al. (48). The parameters with no bracket are taken from the work of Farner (16).
Data for “Jejunum” and “Ileum” by Farner (16) are reported as the “upper ileum” and “lower ileum” in the original article.
This value in the original article was for the “large intestine.”
This value in the original article was for the “rectum.”
Deduced Mechanism Underlying the Emergence of HPAI H5N1 Viruses
Despite a lack of clear evidence, species-specific or tissue (cell)-specific endosomal pH environments could have assisted the ancestral H5N1 virus to acquire a higher pH threshold for fusion, which in turn would have endowed the virus with broad cell tropism. The endosomal pH values in primary cells and some cell lines are cell type or species specific (Table 1). This supports our hypothesis that selection pressures, such as species-specific or tissue (cell)-specific endosomal pH, could facilitate the appearance of the viruses such as HPAI H5N1 that have a higher pH threshold; such a virus would be inactivated in an ambient acidic environment but could still induce viral–cell membrane fusion and efficient viral replication in target cells under even higher endosomal pH conditions. Several reports argue for a possible relationship between avian host species and the pH threshold for HA-mediated membrane fusion of AIV strains. Giannecchini et al. (25) reported that LPAIV H7N3 strains isolated from turkeys had a lower pH threshold than related strains isolated from mallard. Okamatsu et al. (60) found no correlation between the HA pH threshold in 50 LPAIV strains isolated from ducks and chicken. Similarly, Galloway et al. (22) found no clear relationship between the acid stability of AIV HA proteins and host bird species such as duck and mallard and terrestrial birds including turkey, chicken, gull, and tern. Further studies should explore the possible mechanisms underlying the generation of human pathogenic virus such as HPAI H5N1 virus from the view point of HA gene stability. Also, it would be interesting to examine possible relationships between viral infectivity and the pH dependency of HA activation in AIV subtypes such as H5N6, H7N3, H7N7, H7N9, H9N2, H10N7, and H10N8, which have also been transmitted directly from birds to humans (9,20,37,49,81,89,108,110).
Closing Remarks
Influenza virus harbors a multifunctional HA protein that facilitates both viral entry and viral genome release after receptor binding and viral–cell membrane fusion, respectively. The balance between receptor tropism and HA acid stability has allowed influenza viruses to evolve to exhibit various characteristics, as observed for highly transmissible strains such as H1N1pdm or highly pathogenic strains such as HPAI H5N1. Accumulated knowledge, together with further research, about the HA-governed mechanism of HPAI H5N1 virulence (i.e., receptor tropism and pH-dependent viral–cell membrane fusion) will be helpful for developing effective surveillance strategies and for prevention/control of HPAI H5N1 infections.
Footnotes
Acknowledgments
This work was supported by Grants-in Aid for Scientific Research 22790953, 24591480, and 15K09576 from the Ministry of Education, Culture, Sports, Science and Technology; and by JSPS Core-to-Core Program, B. Asia-Africa Science Platforms, a grant from Takeda Science Foundation, and a grant from the Japan Foundation for Pediatric Research (Grant No. 16-013).
Author Disclosure Statement
No competing financial interests exist.