
Abstract
To evaluate the association of the -308 and -238 tumor necrosis factor alpha (TNF-α) gene polymorphisms with clinical manifestations of dengue and TNF-α serum levels in a northwestern Mexican population. The study populations included dengue fever (DF) and dengue hemorrhagic fever (DHF) patients, and a group of healthy controls (HCs) without history of dengue. Polymerase chain reaction–restriction fragment length polymorphism and Enzyme-Linked Immunosorbent Assay were performed to determine genotypes and serum concentration of TNF-α, respectively. There were no significant differences in alleles, genotypes, and haplotype frequencies between patients and HCs. However, when patients were separated into DF and DHF, there was an increased prevalence of the -308 GA genotype in HCs compared to DHF (odds ratio [OR] = 0.129, 95% confidence interval [CI] = 0.018–0.945, p = 0.025), as well as the GG haplotype (OR = 0.49, 95% CI = 0.273–0.880, p = 0.01757) in DF. The genotypes of both polymorphisms were not associated with hematologic manifestations. Serum TNF-α levels were significantly higher in patients than in HCs (p = 0.004). Our results suggest a minimal effect of the -308 and -238 TNF-α gene polymorphisms in dengue patients and that their increased serum levels of TNF-α are independent of genotypes.
Introduction
A
The pathogenesis of dengue has been the focus of intense biomedical research for several decades. It is well known that both the virus and the human host possess biological features, which can be considered predisposing, or resistance factors to infection or clinical progression. Factors for predisposition or resistance include a secondary infection with a heterologous serotype (23), differences in the virulence of virus strains (41), primary infection in infants born to dengue-immune mothers (6,16), and an appropriate antiviral inflammatory response combined with an effective immune regulation (43); the latter depends on a complex set of molecules such as vitamin D (30) and cytokine receptors (5), lectins (40), antimicrobial peptides (9), free radical scavengers/oxidative stress inducers (10), and proinflammatory and anti-inflammatory mediators (20).
Tumor necrosis factor alpha (TNF-α) is a key proinflammatory cytokine that participates in many processes, such as lipid metabolism (13), host defense (28), coagulation (14), regulation of apoptosis (36), endothelial function (39), and transcriptional induction of some cytokines (42). Regarding the latter process, the two main genetic polymorphisms of TNF-α (-308G/A and -238G/A) have been studied regarding their involvement in the pathogenesis and consequences of dengue virus infection with controversial and contrasting results, ranging from associating them with severe manifestations, with a protective effect and a neutral effect in different populations (1,4,15,17,19 –21,25,38,39,46, 48,53).
This work studied the relationship between TNF-α gene polymorphisms and their circulating serum levels, and the susceptibility to dengue virus infection and its different clinical and laboratory manifestations in an endemic region of Mexico. We also reviewed the available literature about the possible influence of the -308 and -238 TNF-α gene polymorphisms on the outcomes of dengue infection.
Materials and Methods
Study subjects
This case–control study was conducted at the Hospital General de Culiacan in the state of Sinaloa, Mexico, during the period 2014–2016. Following the 1997 guidelines for Dengue by the WHO, the study included 251 adult patients admitted to the emergency services, 182 of whom had DF and 69 of whom had DHF. Acute dengue infection was confirmed by the SD BIOLINE Dengue Duo rapid test (NS1 Ag and IgM/IgG).
The results of the clinical evaluation and laboratory analysis, demographic data, and other parameters related to the dengue infection, except the infecting dengue virus serotype, were obtained from electronic medical record systems and entered into a database.
The control group included 275 healthy blood donors negative for IgM and IgG antidengue antibodies, Human Immunodeficiency Virus, and Hepatitis C Virus, and who had no known history of hospitalization for dengue-like illnesses.
To be able to determine TNF-α levels, any person presenting a history of inflammatory and chronic degenerative disorders was discarded as healthy control (HC).
All participants were fully informed of the study protocol before signing the written informed consent. The study was approved by the Institutional Ethics and Research Committee of the Hospital General de Culiacan. The parents and grandparents of patients and HCs were born in the state of Sinaloa, ensuring ancestral homogeneity.
DNA extraction and TNF-α genotyping
Genomic DNA was obtained from peripheral venous blood samples, as previously described (26), and stored at −20°C until processed. A polymerase chain reaction (PCR)–restriction fragment length polymorphism (RFLP) assay was used for genotyping. For the -308G/A polymorphism, the primers used were 5′-AGGCAATAGGTTTTGAGGGCCAT-3′ and 5′-TCCTCCCTGCTCCGATTCCG-3′; for the -238G/A polymorphism, the primers used were 5′-AGAAGACCCCCCTCGGAACC-3′ and 5′-ATCTGGAGGAAGCGGTAGTG-3′ (48).
The PCR reaction yielded a 107 bp amplicon for the -308G/A polymorphism, which was cut with the NcoI restriction enzyme (fragments of 87 and 20 bp for G allele and 107 bp for A allele); for the -238G/A polymorphism, the PCR generated a 152 bp amplicon, which was subjected to digestion with the MspI restriction enzyme (fragments of 133 and 19 bp for G allele, and of 152 bp for A allele).
The PCR-RFLP product of the TNF-α genotyping was visually examined in polyacrylamide gel (9%) stained with silver nitrate. In each PCR-RFLP run, a known heterozygous DNA sample was used as restriction control (these DNA samples were kindly donated by PhD. Nora Magdalena Torres-Carrillo, from the Biomedical Research Center, University of Guadalajara). The genotyping results were confirmed by a double-blind evaluation by an independent observer who randomly selected 40% of GG genotypes and 100% of GA + AA genotypes, and obtained the same results.
TNF-α levels
Sample collection and processing
To determine serum TNF-α levels, venous blood samples were taken from patients with dengue (13 DF and 3 DHF) and patients with other febrile illnesses (11 other febrile illnesses [OFIs]) but negative to NS1, IgM, and IgG. All samples were taken during the acute phase (up to 5 days after the onset of symptoms). In addition, we analyzed samples from 33 healthy volunteers without antibodies against the dengue virus.
Serum levels of TNF-α were measured using a commercial kit (Platinum ELISA kit; eBioscience) and calculated based on a curve generated by plotting the optical density values of the known standards on log–log graph paper.
Statistical analysis
A Chi-square test was used to evaluate the association between the qualitative variables (distribution of symptoms and signs according to the clinical type of dengue, and of genotypes according to bleeding manifestations), and a Mann–Whitney U test was used to analyze quantitative variables with a nonnormal distribution (comparing the laboratory findings between DF and DHF, and the TNF-α levels with genotypes). The allele and genotype frequencies were calculated by direct counting. To estimate the Hardy–Weinberg equilibrium (HWE) and the association between polymorphisms and disease, we used the online DeFinetti program (
Also, STATA software version 13 was used for search of association between polymorphisms and haplotypes with disease. The correlation between the means of the quantitative variables was evaluated using Dunn's multiple comparison post hoc test (comparing TNF-α levels between patients, OFIs, and HCs), after the Kruskal–Wallis test. The IBM SPSS Statistics 20.0 was used to analyze qualitative variables and GraphPad prism 6.0 was used for quantitative variables. A significance level of p < 0.05 was considered statistically significant.
Results
Demographics, clinical presentations, and laboratory findings in patients
The Table 1 describes and compares the distribution of demographic, clinical, and laboratory data of patients with dengue (182 DF and 69 DHF). As expected, retro-orbital pain, abdominal pain, vomiting, and exanthema were more common in DHF patients, as well as high activated partial thromboplastin time, high hematocrit, and low platelet count (p < 0.05).
Demographic and laboratory data are expressed by mean and SD, p-value was obtained using χ2 test. Symptoms and signs are expressed in absolute value and percentages, and were analyzed by the Mann–Whitney U test.
CI, confidence interval; DF, dengue fever patients; DHF, dengue hemorrhagic fever patients; ns, nonsignificant; OR, odds ratio; PTT, partial thromboplastin time; SD, standard deviation.
TNF-α -308 and -238 polymorphisms, and dengue infection
In patients, the alleles and genotypes are in HWE, with the exception of the -308G/A genotype. Table 2 shows the allelic and genotypic distribution. For the -308 polymorphic site, 12 patients were not included in analysis due to technical problems in amplification process. For -308A allele, the 6.8% of the general population are carriers, similar to the frequency observed in the patients who participated in the study (6.3% for total patients; 5.2% for DF and 9.0% for DHF), while that the frequencies of the -238A allele were 5.3% and 5.6% (4.4% and 8.7% for DHF), respectively. The comparisons of these results were not statistically significant, neither the genotypic distributions between patients and healthy controls for the two polymorphisms. However, when the patients were separated into DF and DHF, the -308G/A heterozygous genotype had a significantly higher prevalence in healthy controls compared to patients with DHF (odds ratio [OR] = 0.12, 95% confidence interval [CI] = 0.018–0.945, p = 0.025 with DeFinetti and 0.04 with STATA programs). For polymorphism at position -238, we do not find any differences between the groups of study.
Single nucleotide polymorphism.
In agreement with DeFinnetti and STATA programs, the TNF-α-308 GA genotype frequency was significantly higher in healthy controls than DHF patients (p = 0.02, p = 0.04, respectively). Both programs yielded identical CIs and OR (95% CI 0.018–0.945; OR = 0.129).
p-Value was calculated using Chi-square test with DeFinetti online program (
Four haplotypes were formed with significant linkage disequilibrium (D′ >0.812, R2 = 0.002). The GG was the most common haplotype observed (Table 3); its prevalence was higher in DF compared to DHF patients (OR = 0.49; 95% CI = 0.273–0.880, p = 0.0175). Thus, the -308G/A heterozygous genotype and GG haplotype can be considered protective factors against DHF.
p-Value was calculated using Chi-square test with STATA software version 13.
The GG haplotype frequency was significantly higher in DF than in DHF patients (p = 0.017) that is in accord with the SHEsis program.
Serum levels of TNF-α in study subjects
Figure 1 shows the mean level of serum TNF-α in 16 dengue patients, 11 OFIs, and 33 HC: 21.94, 23.13, and 14.71 pg/mL, respectively. Significant differences were found between patients with dengue (p = 0.004) and OFIs (p = 0.020) compared to healthy controls, as well as between DF and DHF (p = 0.026); conversely to the first two groups that showed no significant differences. In this small sample, we did not find a relationship among TNF-α levels and genotypes (data not shown).
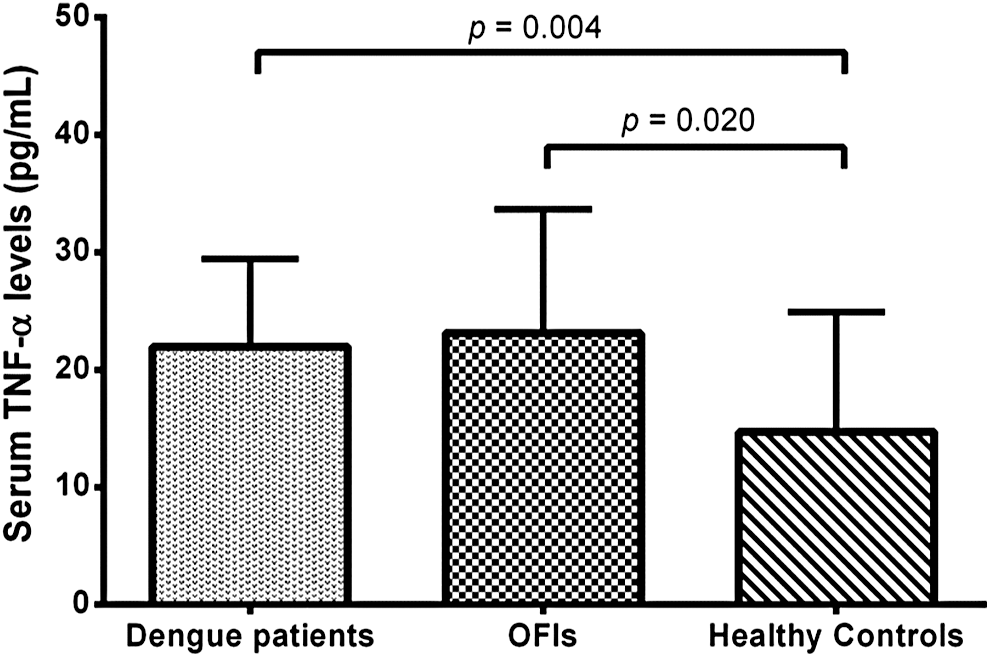
TNF-α serum levels in dengue patients, OFIs, and healthy controls. The p-value was calculated by the Kruskal–Wallis Dunn's multiple-comparisons test. OFI, other febrile illness; TNF-α, tumor necrosis factor alpha.
Discussion
Tumor necrosis factor alpha is a key molecule in human innate and adaptive immunity, and has been studied in numerous diseases such as infections, autoimmune and inflammatory diseases, and cancer (42). In the last 10 years, there has been an increased interest in studying the possible participation of TNF-α in genetic susceptibility/resistance to dengue infection and its progression to more severe forms of illness. Unfortunately, only a few populations have been studied (1,4,15,17,19 –21,25,38,46,53). This study demonstrated the association of -308 GA heterozygote genotype and GG haplotype of the TNF-α gene with dengue in an endemic area in northern Mexico. The above could be interesting because it suggests similar and contrasting conclusions, which are discussed below.
The results of this study indicated that GA genotype of the -308 TNF-α polymorphism is associated with protection against severe forms of dengue in a specific population. Similar results have been reported in a Malaysian population (46), but those results differed with several previous studies with Indian, Thai, Mexican, and Brazilian populations, which discarded this genotype as a strong modulator of susceptibility or resistance to dengue (1,15,25,53). In contrast, in Sri Lanka, the GG genotype (21), and the A allele in Venezuela (20) and Cuba have been associated with susceptibility to DHF (38); however, these results are inconsistent because healthy controls deviated from the HWE.
Regarding the -238 polymorphism, only the heterozygous genotype showed an important close-up to difference in percentage terms (17.4 vs. 9), but nonstatistically significant (OR = 2.18; 95% CI = 0.075–4.894, p = 0.05), between DF versus DHF patients, which allow us to discard that there is an imminent risk for developing DHF in those infected with dengue virus.
Unfortunately, there is very little scientific information about the participation of this polymorphism in the pathogenesis of dengue. García-Trejo et al. (25) in a study of a small sample of patients in the state of Morelos, in South-Central Mexico, suggested that the A allele could confer protection against dengue and the wild-type G allele could increase the risk of dengue (although the latter was not published), but Sam et al. (46), in a study of a Malaysian population, argued that the predisposition to severe forms of dengue is found in carriers of the A allele and the GA genotype, in agreement with the results of Vejbaesya et al. (49) for ethnic Thai children, who found a significant association between the TNF-4 haplotype, which includes the -238A allele (and the -308G and +488G alleles), and the severity of dengue (OR = 2.38). In this Thai population, the TNF-3 haplotype, comprised by GGA alleles (-308, -238, and +488 positions, respectively), was associated with protection to dengue (OR = 0.08 and p = 0.019); this is similar to the findings of this study, in which the prevalence of the GG haplotype is lower in DHF than in DF patients (OR = 0.493 and p = 0.0175), suggesting its association with a reduced risk of severe forms of dengue.
The similarity and the differences between the results of different studies could be explained by the differences in sample size, but the population structure could also be a confounding factor (32). Rubi-Castellanos et al. (44), using STRs (Short Tandem Repeat) for human identification, found important differences in ancestral components of Mexican-Mestizos; more recently, Mendoza-Carrera et al. (33) described an increasing frequency gradient of allele -308A toward northwestern Mexico (9.7% for Culiacan and 6.9% for Monterrey) and a decreasing gradient toward southeastern Mexico (6.7% for Guadalajara, 3.3% for Puebla, and 4.1% for Veracruz). The similarity between the findings of our study and the study of the Malaysian population, including the frequency of the -308A allele of 10%, could be explained by the sample size (46).The above is of great value from an anthropological point of view because the main non-Amerindian ancestry of Mexican-Mestizos comes from Europe, mainly Spain (with a frequency of 10% for the -308A allele) (31). However, the Morelos study included a reduced sample size of patients (85 with DF and 45 with DHF) and also found a reduced frequency of the -308A allele in healthy controls (of 4.3%) (25), similar to Puebla, its neighboring state.
In summary, to the best of our knowledge, there are 11 reports about the influence of the -308A polymorphism on the predisposition/resistance to dengue, five of which discard a potential association (three in Brazil and two in India) (1,4,17,19,53). The other studies suggest that the A allele and GG genotype are protective factors in Malaysia and Cuba, respectively. The results of this study also suggest a protective effect of the GA genotype (38,46). Regarding the progression to DHF, the A allele in Venezuela and Cuba (20,38) and the GG genotype in Malaysia and Sri Lanka have been suggested as predisposing factors. With respect to the polymorphic site -238 (25,46), the A allele and GA genotype and G allele have been suggested as risk factors to developing DHF in Malaysia and dengue per se in Morelos, Mexico, respectively. These findings are not seen in this study (25,46).
This integrative analysis of the possible association of dengue with the TNF-α gene led us to discard that these two polymorphisms play a preponderant role as genetic modulators of dengue infection and its outcomes. Moreover, due to their low frequency, they cannot be used as biomarkers for individual susceptibility to dengue, at least in the studied population. Only a minimal effect can be expected, in addition to the effect of many other genetic polymorphisms such as HLA (4,8,18,23,24,34,49), DC-SIGN (2,37,45,51,53), VDR (3,31), and the FcγRIIa alleles (17,23,24,35,37), which seem to play a role in the pathogenesis of dengue.
The association of TNF-α gene polymorphisms with bleeding manifestations was discarded when they were grouped together. However, the epistaxis and rectal bleeding were associated, in an isolated way, with the polymorphisms. We consider this as a spurious association that can be explained by the small sample size. To say, eight patients with epistaxis had the GG genotype and four patients had the GA + AA genotype; in the case of rectal bleeding, three patients had the GG genotype and three patients had the GA + AA genotype (data not shown). Our findings contrast with the results of Chuansumrit in Thai children with bleeding manifestations, who tended to have a higher frequency of the 308A allele (15). Such differences can be explained by the different characteristics of the studied populations. It is well known that children are particularly vulnerable to a high burden of disease and secondary DENV (Dengue virus) infection is a risk factor for severity in children (27); last, but not least, there are serotype-specific differences (7,22).
There are several hypotheses explaining the pathogenesis of the severe disease caused by the dengue virus, and they all coincide in attributing a highly relevant role to cytokines. TNF-α levels were associated with DHF for the first time in 1991 (50). A possible role as modulator of the pathophysiological changes observed in DHF/DSS was also proposed for TNF-α in that year (54). Although the sample size was small in both studies, their results marked the beginning of a new era of research in dengue disease. In this study, TNF-α serum levels were quantified and compared between dengue patients, OFIs, and healthy controls; they were higher in patients with dengue per se (p = 0.004) and OFIs (p = 0.020) compared to healthy controls, but similar between the first two groups (p > 0.5).
The distribution of two polymorphisms among the studied groups showed no differences. This is an expected finding, considering that the integrity of the TNF-α system could improve the survivability and management of DENV infection (35).
The comparison of mean level of serum TNF-α between 13 DF (23.26 pg/mL) and 3 DHF (16.21 pg/mL) was significant (p = 0.026). This contrasts with the results of studies in Thailand and India (11,29), but it matches in relation to levels significantly elevated, found in the disease group compared to the control group. Unfortunately, we studied, in addition to 11 OFIs and 33 healthy controls, only 16 patients (13 with DF and 3 with DHF). This reduced number is a great limitation, and so, these results deserve to be corroborated. We used such a small sample because the outbreak was ending when the study started and we decided not to use frozen samples.
The similarity of the results between patients with dengue and OFIs, coupled with their lack of correlation with genotypes in this study and others (15), as well as the fact that time of specimen collection for measuring cytokines could affect the actual values of cytokines, led us to discard the utility of TNF-α levels as biomarkers for monitoring the severity of this disease. Sinaloa is a subtropical state of Mexico with other endemic febrile illnesses. It is very possible that the patients with OFIs have actually brucellosis, salmonellosis, or leptospirosis, which can cause a cytokine waterfall characterized by high levels of TNF-α, among other immunological mediators (47).
Our results suggest a minimal effect of the -308 and -238 TNF-α gene polymorphisms in susceptibility/predisposition to dengue in this Mexican population. Furthermore, the increased TNF-α serum levels seen in dengue patients are independent of genotypes; however, previous studies support a possible association of the TNF-α system with vascular endothelial homeostasis.
Footnotes
Acknowledgments
We are grateful to PROFAPI/UAS 2014/225 for the grant given to Dr. Raul Borrego Gaxiola, for the constant support of the Hospital General de Culiacan “Bernardo J. Gastélum,” and especially to all the participating subjects. M.G.S.L. was a Research Fellow at the Master of Biomedical Sciences, Faculty of Chemistry and Biological Sciences, Sinaloa Autonomous University, and received a National Council of Science and Technology MD scholarship number 364040. We also thank Dr. Norma Torres-Carrillo of the Biomedical Research Center, University of Guadalajara, for the kind donation of the TNF-α -308 and -238 heterozygous DNA samples used as controls.
Author Disclosure Statement
No competing financial interests exist.