
Abstract
Endocannabinoid system plays an important role in pathophysiologic processes such as immune functions and impacts on disease severity. Our previous study showed that cannabinoid receptor 2 (CB2) affects clinical course of respiratory syncytial virus (RSV) infection. In this study, we investigated the role of cannabinoid receptor 1 (CB1) in RSV immunopathology and its therapeutic potential in mice model. To study the role of CB1 receptors in the immunopathology of RSV, CB1 was blocked daily with AM281 as a selective antagonist in Balb/c mice and were infected by intranasal inoculation of RSV-A2 24 h following the first dose of antagonist administration. The potential pharmacological therapeutic effects of cannabinoid receptor activation during RSV infection were studied using JZL184 as a selective indirect agonist, 24 h after infection. Mice were sacrificed on day 5 after infection and experimental analyses were performed to study the CB1 receptor expression, airway immune cell influx, cytokine/chemokine secretion, lung histopathology, and viral load. RSV infection of airways significantly induced the expression of CB1 receptors in lung cells of mice. Blockade of CB1 receptors using AM281 enhanced immune cell influx and cytokine/chemokine production, and aggravated lung pathology. Activation of cannabinoid receptors using JZL184 decreased immune cell influx and cytokine/chemokine production, and alleviated lung pathology. This study and our previous finding indicated that endocannabinoid signaling regulates the inflammatory response to RSV infection, and is a potential therapeutic candidate for alleviation of RSV-associated immunopathology.
Introduction
R
Cannabinoids are a diverse group of endogenous and exogenous/synthetic compounds that function through the activation of cannabinoid receptors (34). Two types of G protein-coupled cannabinoid receptors have been cloned and were classified as cannabinoid receptor 1 (CB1) and cannabinoid receptor 2 (CB2) (29), which principally expressed in the central nervous system (CNS) and gastrointestinal tract. However, other tissues, including spleen, thymus, and leukocytes, were also shown to express these receptors (19,26,28). The high-level expression of CB2 receptors in immune cells as well as the upregulation of these receptors in inflammation support the notion that the anti-inflammatory and immunomodulatory activity of cannabinoids can occur in a CB2 receptor-dependent manner (9). However, as suggested by some studies, CB2 receptors may not be the only cannabinoid-based mediator of the immune system; therefore, CB1 receptors may also act as an anti-inflammatory and immunomodulatory factor (4,6). Indeed, in vivo studies on knockout mice highlighted the essential role of both CB1 and CB2 receptors in cannabinoid-mediated beneficial effects (7,27).
The effects of cannabinoid receptors upon the immune system gained many attentions over the past years, as they have the potential to compromise host immunity and may directly and/or indirectly impacts on pathogenic agents (18,33). In this regard cannabinoid signaling was shown to interact with several key metabolic pathways, enzymes, receptors, and transcription factors involved in inflammation (19,34). Given the potential effects of cannabinoid-mediated regulation of immune function and the complicated RSV-associated immunopathology, is tempting to speculate two hypotheses: (i) the endocannabinoid signaling might participate in the RSV infection outcome and (ii) targeting of cannabinoid receptors may be a novel approach for alleviation of RSV immunopathogenesis. We have recently shown that the functional CB2 genetic polymorphism (CB2 Q63R) is associated with the risk of hospitalization in children with acute RSV infection (31). In addition, CB2 receptor was found to protect against experimental RSV disease by controlling immune responses in mice model (31). In the light of these findings, this study was aimed to investigate the role of CB1 receptors in a commonly used animal model of RSV infection, Balb/c mice. We applied selective CB1 antagonist (AM281) to investigate the plausible CB1 signaling involvement in the clinical outcome of RSV pathogenesis. Furthermore, JZL184 as a potent and selective indirect cannabinoid agonist was used to test the activation effects of cannabinoid receptors on clinical outcome. Our findings further highlighted the essential role that cannabinoid signaling can play in RSV infection outcome.
Materials and Methods
Animals and drugs
Female Balb/c mice (6–9 weeks old, weighing 15–18 g) were purchased from the Institute Pasteur of Iran (Karaj, Iran) and kept in-house for 1 week before experiment. Mice were housed in individual cages in a controlled environment with access to standard laboratory food and water ad libitum. This experiment was approved by the animal ethics committee of the Tehran University of Medical Sciences (No. 34743).
The AM281 as a potent and selective CB1 receptor antagonist/inverse agonist and JZL184, a potent and selective inhibitor of monoacylglycerol lipase (MAGL) known as cannabinoid receptors indirect agonist, were obtained from Cayman Chemical. The drugs stocks were dissolved in dimethyl sulfoxide (DMSO) and administered intraperitoneally (i.p.) in normal saline 0.9%.
Experimental procedure
Mice were randomly assigned into six groups: controls (PBS-PBS, PBS-RSV, AM281-PBS, and PBS-JZL184) and experimental (AM281-RSV and RSV-JZL184) groups. The AM281 was injected i.p. twice a day at a dose of 2 mg/kg. One day after the first antagonist treatment, mice were anesthetized with ketamine/xylazine and intranasally infected with RSV-A2 at 5 × 106 pfu/50 μL/mice dose. The JZL184 was injected i.p. 24 h after viral infection (5 × 106 pfu/50 μL/mice) twice a day at a dose of 4 mg/kg. Drugs were administrated daily until 5 days after viral infection. The control groups received a similar volume of phosphate-buffered saline (PBS).
Mice were anesthetized and sacrificed on day 5 after infection (the peak day of immune cell influx and viral load) and bronchoalveolar lavage fluid (BALF) and lungs were obtained to measure CB1 receptors expression, airway immune cells influx, cytokine/chemokine secretion, lung histopathology, and viral load. In this experiment, virus stock was propagated on HEp-2 cells and purification was performed as described previously by our groups (20). It also, lung were lavaged through a catheter inserted into the trachea and by flushing with ice-cold PBS according to our previous work (20).
CB1 receptor expression
Total RNA was extracted from bronchoalveolar lavage (BAL) cells and homogenized left lung lobe using RNX reagent (SinaClone), and complementary DNA (cDNA) was synthesized using a high-capacity cDNA Reverse Transcription kit (Applied Biosystems, Warrington, United Kingdom). The expression level of CB1 receptor gene was determined by a Real-time reverse transcription-polymerase chain reaction (RT-PCR) with an ABI PRISM 7900 Sequence Detection System (Applied Biosystems) using the SYBRs Premix ExTapTM II (Takara).
The reaction conditions were as follows: 95°C for 30 sec, followed by 40 cycles of denaturation at 95°C for 5 sec, annealing at 55°C for 30 sec, and extension at 60°C for 30 sec. Melting curve conditions were as follows: 95°C for 15 sec and 65°C for 1 min. The relative level of gene expression was determined by the comparative threshold cycle method as described by the manufacturer. Levels of CB1 gene expression were normalized to those of the housekeeping gene (β-actin) using the 2−ΔΔCt method and expressed in the graphs as “relative expression.” The following primer pairs were used: CB1: 5′-CTCTGCTTGCGATCATGGTG-3′ (F), 5′-TCTCAGGTCCTTGCTCCTCA-3′ (R), and β-actin: 5′-GCTCTGGCTCCTAGCACCAT-3′ (F), 5′-GCCACCGATCCACACAGAGT-3′ (R).
Airway immune cell influx
BALF was obtained 5 days after infection, and total number of cells and differential cell counts were performed subsequently. Briefly, the lavage fluids were centrifuged, and cells were suspended in 200 μL of PBS. The total number of cells present in the BALF was counted with the aid of a light microscope in Neubauer chambers. For morphological assessment, an aliquot of 5 × 104 cells/100 μL was centrifuged onto glass slides using cytospin (Tashkhis Co.), and differential leukocyte counts were performed on smears stained with Giemsa dye using standard morphological criteria.
Cytokine/chemokine secretion
The cytokines IFN-γ, MIP-1α, and IL-10 were determined in the BALF supernatant with enzyme-linked immunosorbent assay (ELISA) kits (PeproTech), according to the manufacturer's instructions. The concentration of each cytokine/chemokine was determined by interpolation from the standard curve. The threshold of sensitivity for IFN-γ, MIP-1α, and IL-10 was 16, 4, and 39 pg/mL, respectively.
Lung histopathology
Histology slides were prepared as described previously to study lung pathology (24,25). Briefly, right lung lobes were fixed in 10% phosphate-buffered formalin overnight and embedded in paraffin. Tissue sections (5 μm) were prepared on silane-coated glass slides and stained with hematoxylin and eosin. Prepared slides were evaluated by light microscope and histopathologic score was established on the basis of the numbers and distribution of inflammatory cells within the tissues. A mean score with the standard error of the mean (SEM) was calculated for each treatment group.
Viral load
Nucleic acid was extracted from supernatants of BALF using a viral high pure nucleic acid extraction kit following the manufacturer's instructions (Roche). Real-time RT-PCR was performed on an ABI PRISM 7900 Sequence Detection System (Applied Biosystems) using TaqMan polymerase chain reaction (PCR) Master Mix (Primer Design). Known concentrations of RSV-A2 were used to make a standard curve, and results were expressed as copy number/mL.
Statistics
Statistical analysis was performed using GraphPad Prism v6.0 (GraphPad Software, Inc., San Diego, CA). Data are expressed as mean ± SEM. The difference between control and challenged groups was evaluated by using a Student's t test. The criterion for significance was taken to be p < 0.05.
Results
CB1 expression during primary RSV infection
To determine the effect of primary RSV infection on CB1 receptor expression, we measured the CB1 messenger RNA (mRNA) expression in the lung and BAL of mice, 5 days after viral infection. Our results showed that CB1 gene was significantly expressed in lung cells of RSV-infected mice compared with the uninfected control group (p < 0.05). However, no significant difference was found in the amount of the CB1 expression between infected and uninfected mice in BAL cells (Fig. 1).
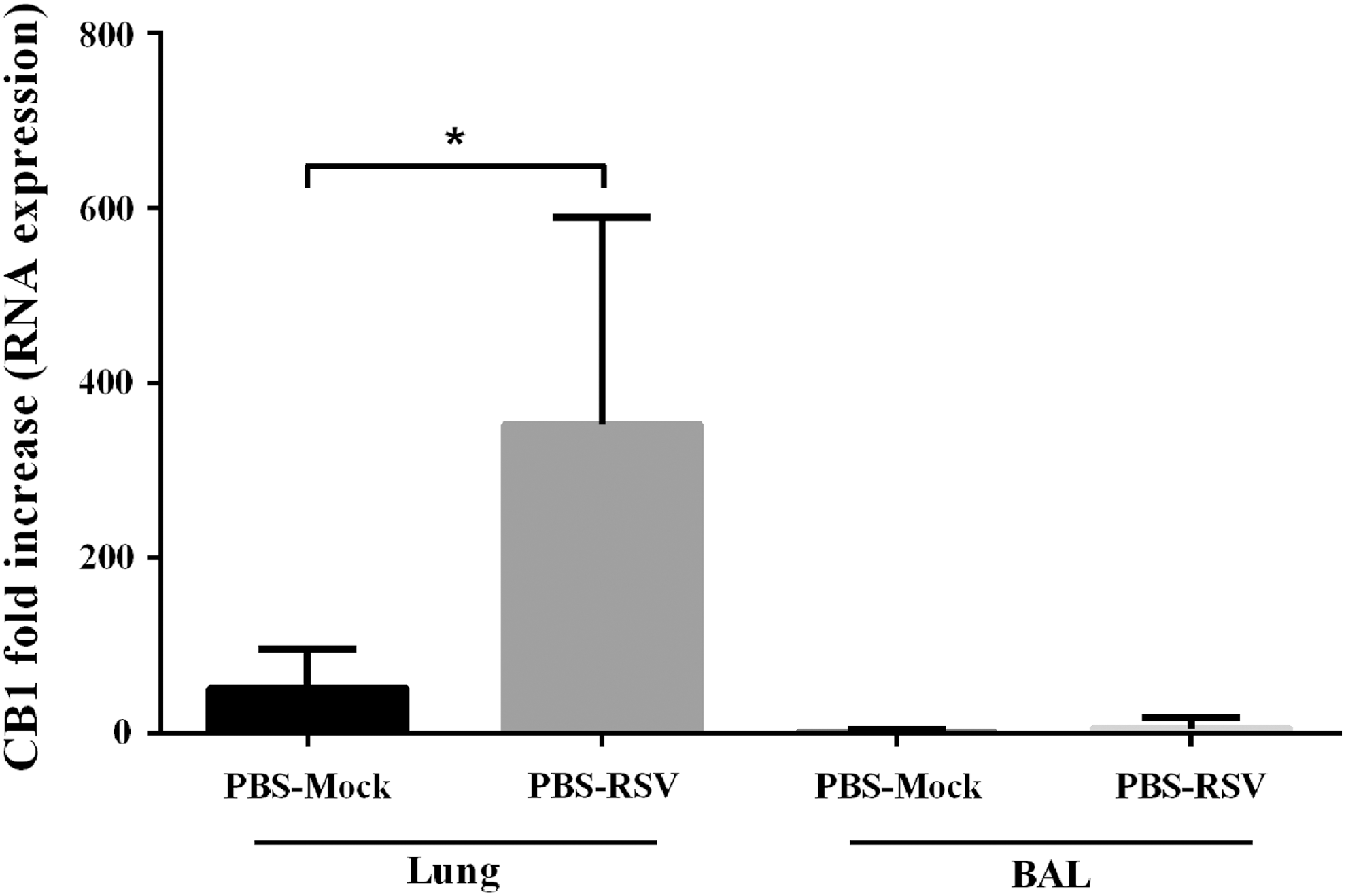
The effect of RSV infection on CB1 receptor expression. Relative expression of CB1 receptors was evaluated in lung and BAL of mice 5 days after RSV or mock infection, using specific primers targeting the CB1 receptors gene, and normalized to those of the housekeeping gene (β-actin). Results represent the mean ± SEM of six animals for each group (*p < 0.05). BAL, bronchoalveolar lavage; CB1, cannabinoid receptor 1; RSV, respiratory syncytial virus; SEM, standard error of the mean.
CB1 blockade during RSV infection
Primary RSV infection elicits an inflammatory response consisting of a mixed population of leukocytes that infiltrates the pulmonary airways. The magnitude of the cell infiltrate was significantly enhanced in CB1 blockade mice following RSV infection (p < 0.05). Differential analysis of leukocytes found in the BALF of mice also showed a significant enhancement of monocytes following CB1 blockade by AM281 (p < 0.05) (Fig. 2). Secreted inflammatory mediators were also measured as an indicator of immune cell function in response to RSV infection. While the level of IFN-γ significantly increased, we found a significantly reduced level of IL-10 production (p < 0.01 and p < 0.05, respectively) (Fig. 3).
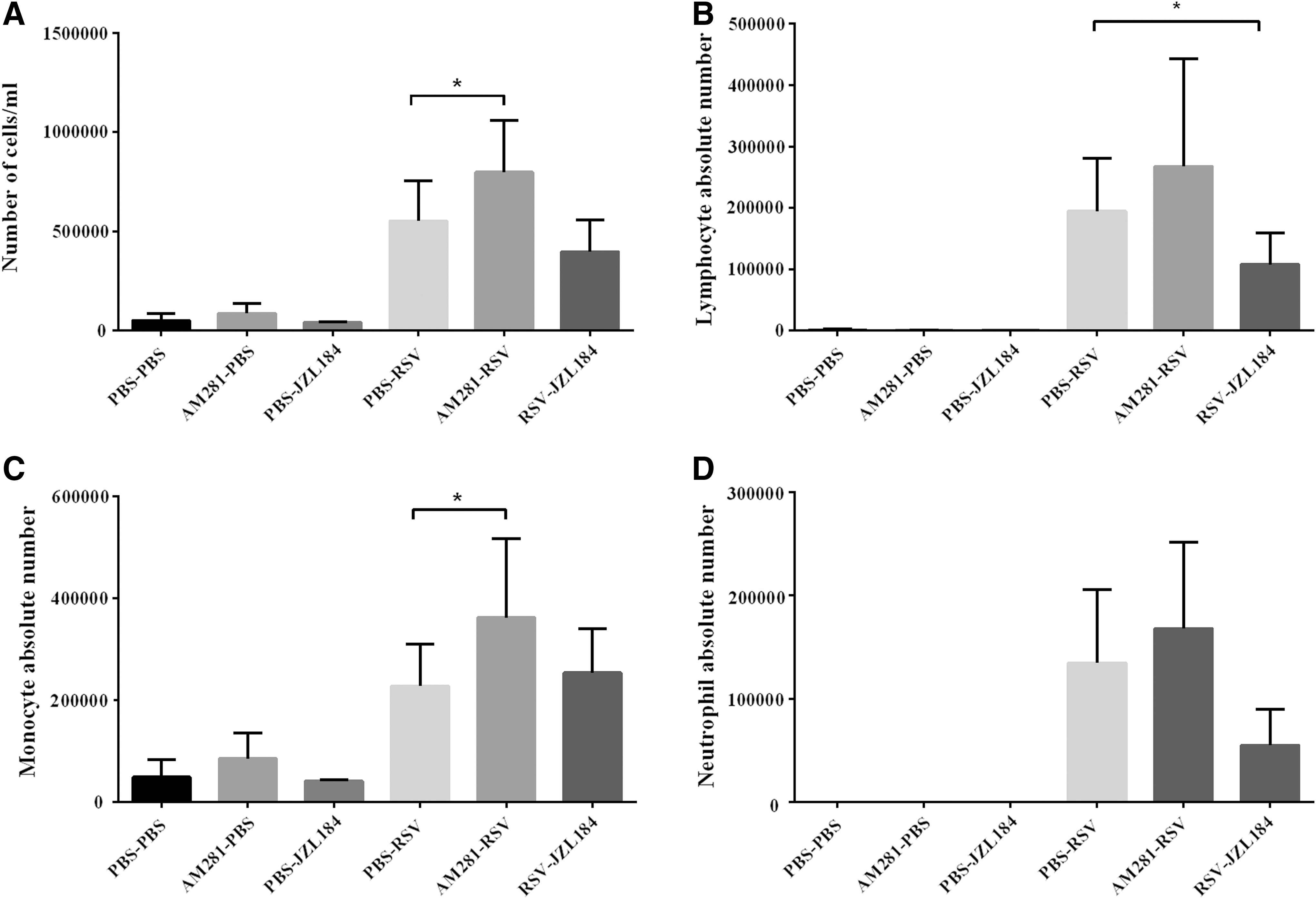
The effect of CB1 receptors on immune cell influx following RSV infection. CB1 receptors were blocked or activated daily and the total cell counts
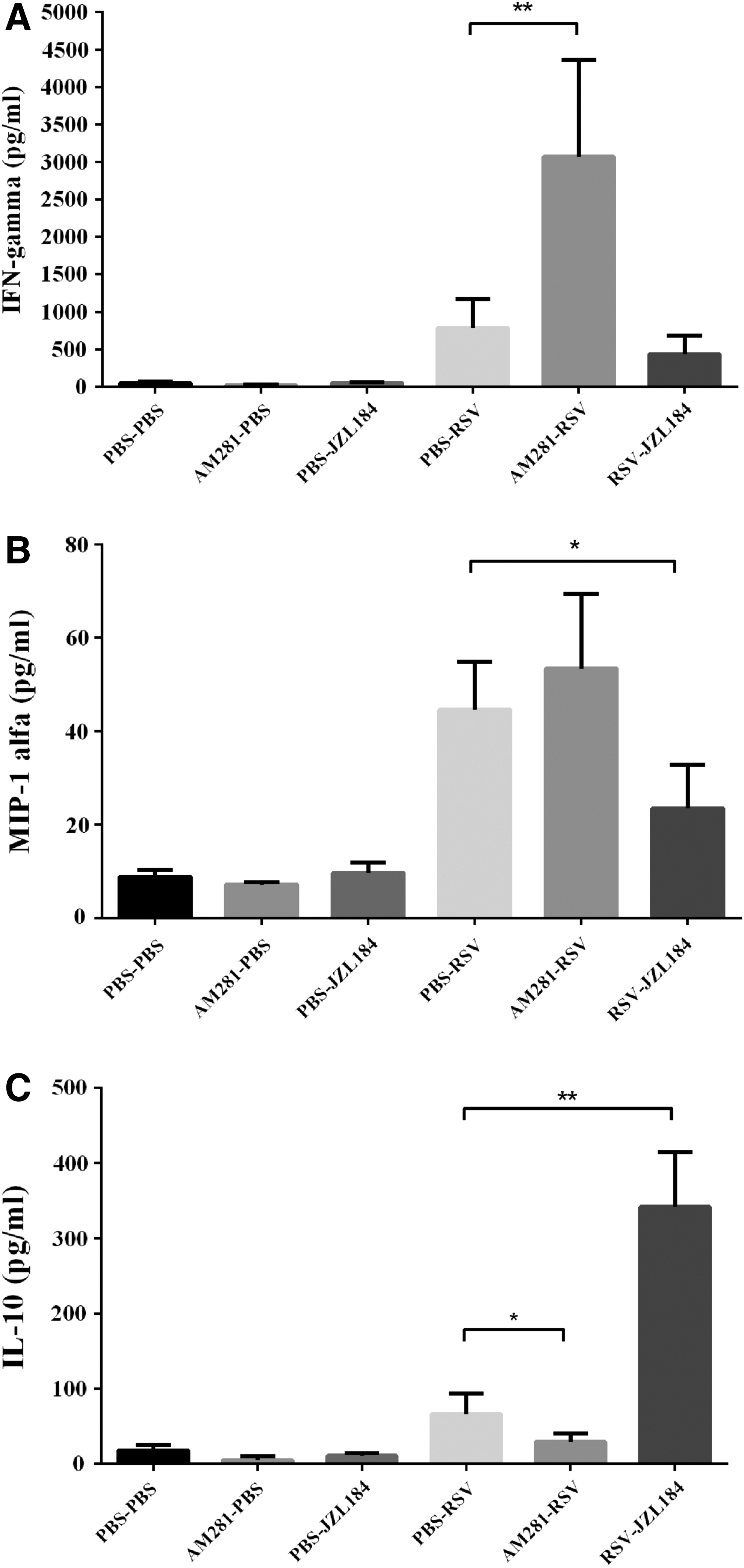
The effect of CB1 receptors on chemokine/cytokine production following RSV infection. CB1 receptors were blocked or activated daily and the IFN-γ
In consistent with the immune cell influx following RSV infection, CB1 blockade subsequently exacerbated lung pathology. Indeed, CB1 blockade had a significant accumulation of immune cells in the peribronchial and perivascular spaces of the lung when compared to control groups (p < 0.05) (Fig. 4). To determine whether CB1 blockade by AM281 has an impact on disease severity, weight loss was daily followed and CB1 blockade was not associated with more weight loss following RSV infection (Fig. 5). Furthermore, based on the enhanced immune cell infiltration and increased tissue injury in RSV-infected lungs of CB1 blockade mice, RSV viral load was assessed using quantitative real-time PCR. Although not significant, CB1 blockade appears to decrease viral replication to some extent (Fig. 6).
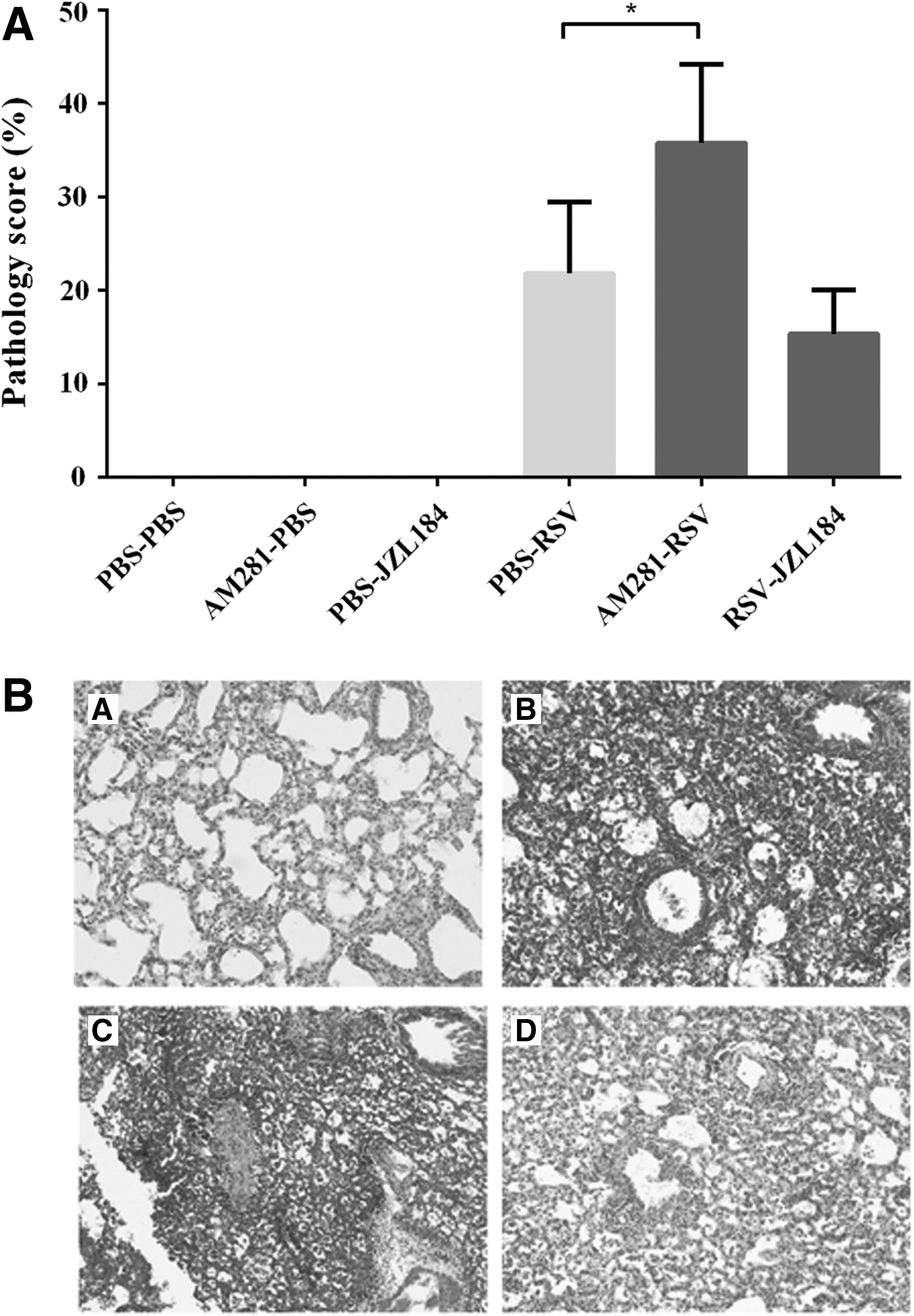
The effect of CB1 receptors on lung pathology following RSV infection. CB1 receptors were blocked or activated daily and the lung pathology was determined on day 5 after infection.
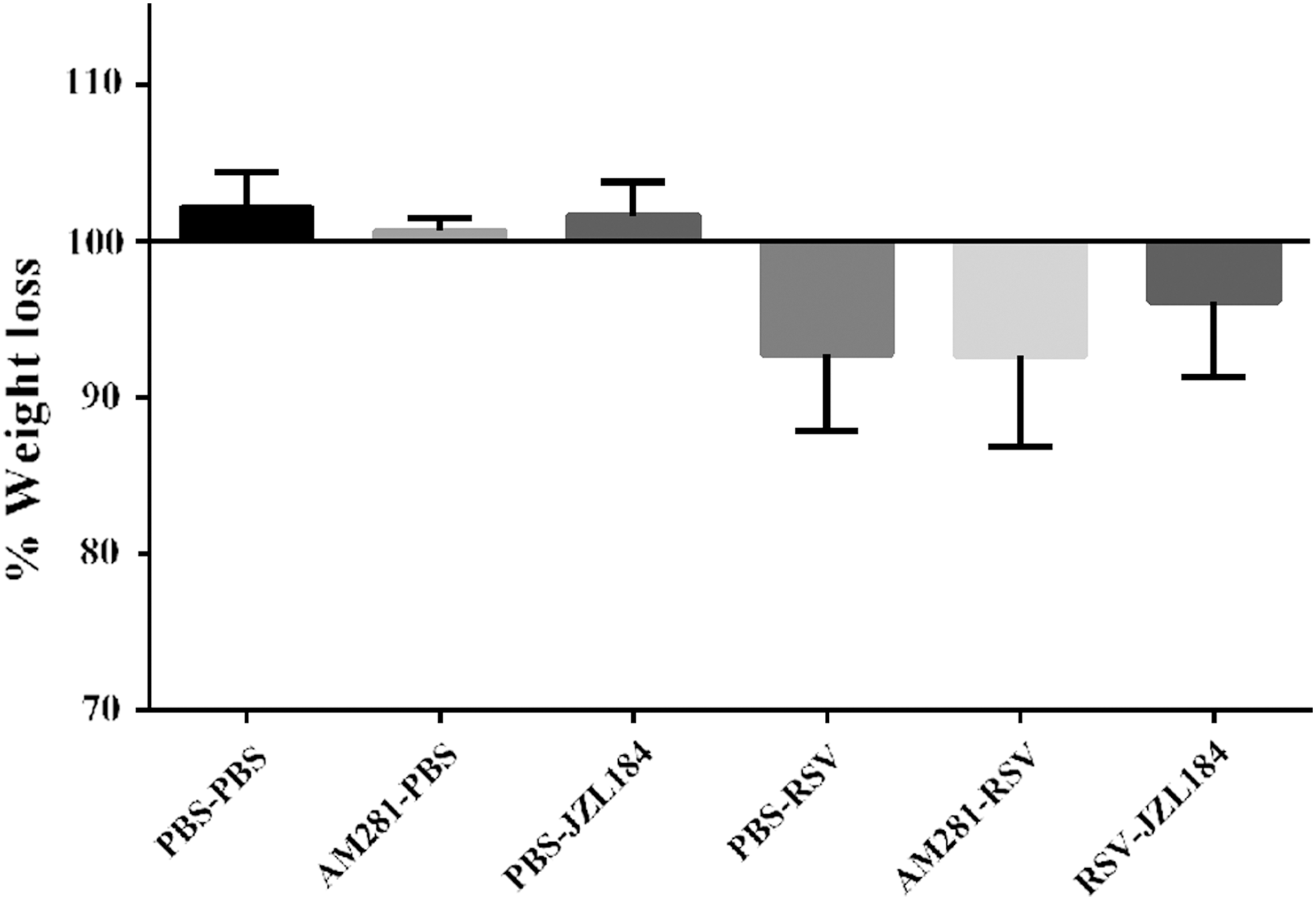
The effect of CB1 receptors on mice weight loss following RSV infection. CB1 receptors were blocked or activated daily and mice weight was measured during RSV infection. The graph shows changes in body weight 5 days after the primary RSV or mock infection.
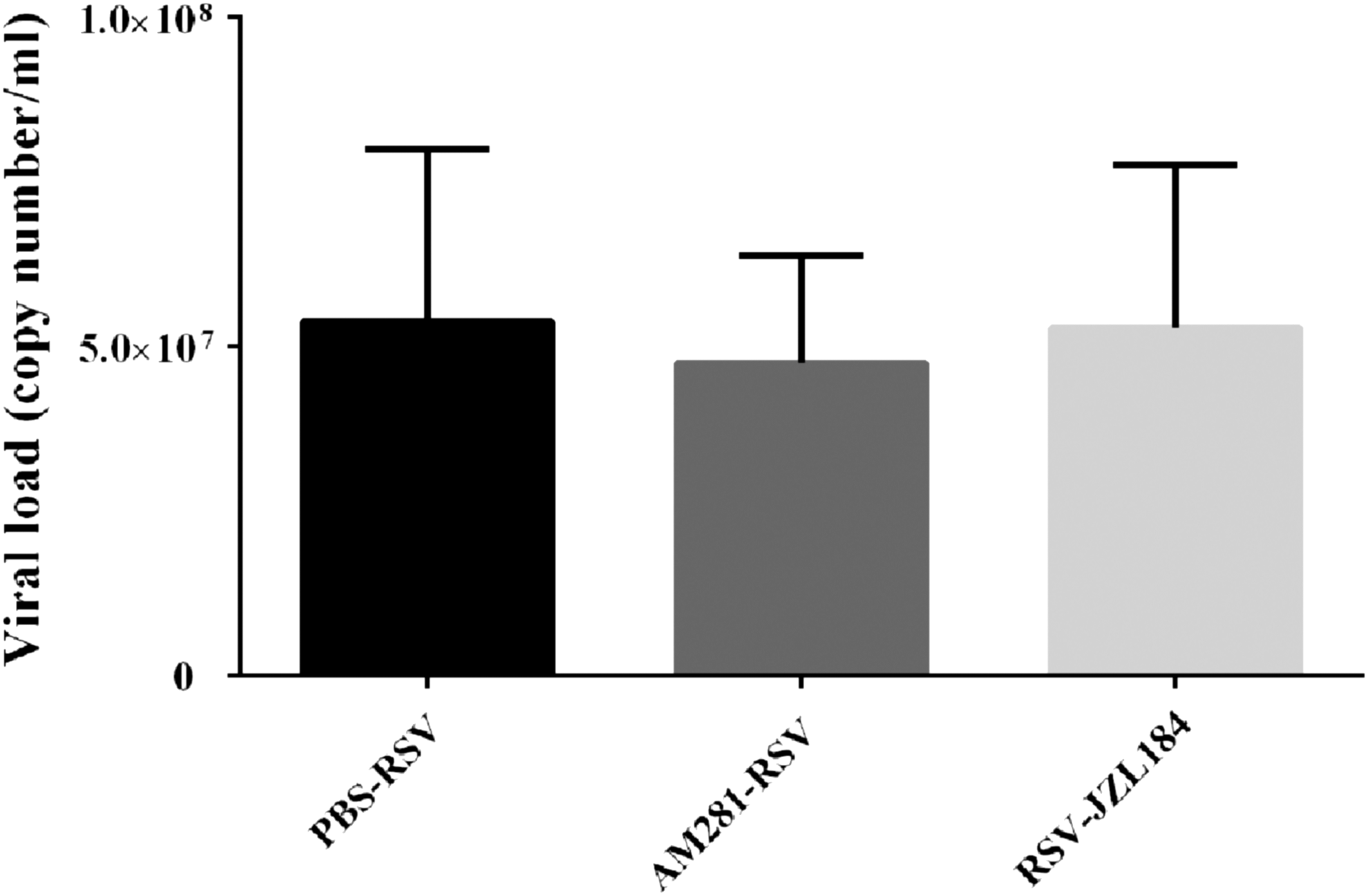
The effect of CB1 receptors on RSV titer following RSV infection. CB1 receptors were blocked or activated daily and the virus copy number was determined by quantitative real-time polymerase chain reaction in BALF supernatant on day 5 after infection. Results represent the mean ± SEM of six animals for each group.
Cannabinoid receptor activation during RSV infection
The roles of endogenously cannabinoid receptor activation in the control of RSV immunopathology were further analyzed using JZL184, a cannabinoid receptors indirect agonist. The data showed that JZL184 treatment decreased the RSV-induced leukocyte migration into the lungs. Compared to the PBS-RSV group, JZL184 treatment significantly decreased lymphocyte counts following RSV infection (p < 0.05) (Fig. 2). While RSV infection significantly induced IFN-γ and MIP-1α, and decreased IL-10 production, JZL184 treatment displayed the opposite role, in which the levels of IFN-γ and MIP-1α decreased, and IL-10 increased after RSV infection, although only IFN-γ and IL-10 were statistically significant (p < 0.05 and p < 0.01, respectively) (Fig. 3).
In agreement with the attenuation of BALF cell infiltration, JZL184 treatment subsequently decreased lung pathology following RSV infection, as evaluated on the basis of reduced accumulation of immune cells in the peribronchial and perivascular spaces of the lung (Fig. 4). Our data also showed that JZL184 treatment as indirect agonist restored weight loss after RSV infection (Fig. 5). However, JZL184 treatment did not show any impact on viral replication (Fig. 6). In this experiment, exposure of mice to the PBS or antagonist and agonist alone did not result in differences within the control groups.
Discussion
Our knowledge of RSV immunopathogenesis and disease severity has increased over the past decades. Understanding how the cannabinoid system, as an interacted system with immune responses, impacts on diseases severity in RSV infection is helpful to facilitate the development of safe and effective therapeutic interventions. The CB1 receptor is profoundly expressed in the CNS and exerts psychoactive function; however, in this study, we found that RSV infection of airways can also affect the CB1 expression in the lung (which prominently contain epithelial cells) and importantly displays immunomodulation. Moreover, RSV infection had no effect in the amount of the CB1 expression in BAL (which prominently contains immune cells)—this is in line with its low-level expression by immune cells (19). We have already shown that RSV infection can affect the CB2 expression in BAL cells, but not in lung cells—that is in line with its high-level expression by immune cells and the inducible expression in inflammatory condition (31). Given the high expression of cannabinoid receptors following RSV infection, they may impact disease severity.
In this study, we demonstrated that CB1 blockade is associated with enhanced bronchoalveolar cellular influx, increased IFN-γ production, and decreased IL-10 production, together with exaggerated RSV-induced lung histopathology. Less data are available regarding the anti-inflammatory and immunomodulatory activity of CB1 receptors. However, our results suggest that the CB2 receptor is not the only cannabinoid-based mediator of the immune response. These results are in line with our previous reports indicating the association of CB2 blockade with enhanced RSV-induced immunopathology (31), consistent with previous reports on the role of endocannabinoid system in influenza infection outcome, in which CB1/CB2 blockade was found to significantly enhance inflammatory response to influenza infection (3,10). Taken together, these results suggest that endocannabinoid signaling can play a significant role in maintaining immune homeostasis and controlling the magnitude of immune response to RSV infection.
Cannabinoid receptor agonists have potential utility as anti-inflammatory and immunomodulatory agents for the treatment of many inflammatory diseases (33). In line with this notion, treatment with JZL184, as cannabinoid receptors indirect agonist, lead to reduced bronchoalveolar cellular influx and MIP-1α production, and enhanced IL-10 production, together with alleviated RSV-induced lung pathology and weight loss. The JZL184, as an indirect agonist, increases the levels of 2-arachidonoylglycerol (2-AG), an endocannabinoid that acts on the cannabinoid receptors, by inhibiting MAGL, the main enzyme responsible for hydrolysis of 2-AG (12). These observations were consistent with the suppressive effects of JZL184 treatment on lung inflammation parameters and colitis reported by others (1,5). In addition, it has been shown that JZL184 treatment decreases neutrophil and macrophage infiltration and proinflammatory cytokine expression in skeletal muscle contusion in rats (8). In this study, AM281 and JZL184 treatment had no significant effect on viral load. This is in line with our previous report, which indicated that CB2 manipulation has no significant effect on RSV replication (31). As indicated previously, the opioid/cannabinoid receptors do not necessarily have a direct effect on viral load; the relevance of cannabinoid receptors on viral infection appears to mainly happen through immune response manipulation by infiltrating the immune cells to the affected tissues (32,33). Importantly, Balb/c mice is a semipermissive model for RSV replication, which may explain the nonsignificant effect (17). Although endocannabinoid signaling reduces the magnitude of the RSV-induced immune response, it might provide protection against excessive airway injury in the lung from exacerbated immunopathology.
Previous reports have shown the association of RSV infection and acute neurologic symptoms such as apnea and seizures (11,30). Since cannabinoid receptors, especially CB1 receptor, are profoundly expressed in the CNS, any alteration in their expression or function following RSV infection might be considered underlying mechanisms behind the RSV-induced neurological symptoms (2). Although our results may support the previously proposed idea that RSV can alter CNS function, however, more studies are required in this area.
In conclusion, our results clearly indicate that (i) RSV infection of airway cells induces CB1 receptor expression in the lung which, may impact RSV immunopathology, (ii) RSV-induced immunopathogenesis can be modulated, in part, by endocannabinoids, as observed by blocking of CB1 receptors, (iii) RSV-induced immunopathogenesis can be modulated, in part, by cannabinoid receptor activation, as observed by receptors activation. These results suggest that CB1 receptors alongside CB2 receptors have anti-inflammatory and immunomodulatory properties and might serve as a therapeutic target in RSV infection by alleviation of virus-associated immunopathology. However, data should be interpreted with caution as different treatment approaches may be needed due to complex immunopathology and mysterious functions of endocannabinoid systems. Although these findings shed light on the pathogenesis of RSV, further comprehensive investigations are warranted in this regard. Based on the results of this project, providing the possible consequence of CB1 receptor activation/inhibition in other cytokine/chemokine level during RSV infection would be interesting. In addition, various RSV challenge doses may better translate the RSV infection seen in children with different disease severity, and it is important to ensure that the JZL184 effects are attenuated with antagonists in future studies.
Footnotes
Acknowledgments
The authors appreciate the financial support of School of Public Health, Tehran University of Medical Sciences (No. 34743). This project was extracted from a PhD thesis.
Author Disclosure Statement
The authors have no potential conflicts of interest to disclose.