
Abstract
Bluetongue is a ruminant infectious disease that is characterized by hyperpyrexia, leukocyte decrease and mucosal ulcerative inflammation changes. In this study, three segments of Bluetongue virus (BTV)-8 VP2 protein (BTV-8A, 8B, and 8C) were cloned into pET-28a (+) and pMAl-c5X vectors and expressed in Escherichia coli BL21 (DE3) as histidine (His)-tagged (His-8A/8B/8C) and maltose-binding protein (MBP)-tagged (MBP-8A/8B/8C) fusion proteins. After purification, His-8A/8B/8C were used to immunize BALB/mice and MBP-8A/8B/8C were used to screen for monoclonal antibody (mAb)-secreting hybridomas. Two mAbs (8B-5D4 and 8B-3G11) that could recognize BTV-8 were obtained. Two competitive enzyme-linked immunosorbent assays with good specificity and sensitivity using mAbs 8B-5D4 or 8B-3G11 as competitive antibody were established. With the joint reaction of these methods, serum samples containing anti-BTV-7 or anti-BTV-8 antibody could be screened out, suggesting that these methods would be useful for the diagnosis and epidemiological studies of BTV-8.
Introduction
B
BTV is classified into 27 serotypes (BTV-1, -2, -3, etc) based on the genetic and antigenic features of the main neutralization antigen VP2. The distribution of serotypes shows a peculiar geographic pattern. For example, among the 26 serotypes, at least five BTV serotypes (BTV-1, 2, 4, 9, and 16) have been described as the most common serotypes in the Mediterranean area (10), but since 2006 a new emerging and highly virulent serotype, BTV-8, has caused severe outbreaks in Northern Europe, not only in sheep, but also in cattle (16). BTV-8 emerged in Northern and Central Europe in August 2006 and it was characterized by a high virulence in sheep and, notably, in the bovine populations (2,14). The recent BTV outbreaks caused by BTV-8, the introduction and circulation of additional serotypes, the finding of Toggenburg Orbivirus (21), and the extensive vaccination against BTV in Europe emphasize the need for a reliable neutralization test protocol. This serotype is also able to cross the ruminant placenta causing fetal infections, in contrast to the other described serotypes in which this type of infection is unusual (3,6). Recent works have also documented the oral route as a way of experimental transmission of BTV-8 (15,17).
Serological detection methods have been considered to be the most widely used clinical detection test, including enzyme-linked immunosorbent assay (ELISA), agar gel immunodiffusion (AGID), and virus neutralization test (VNT). In 1962, Klontz established AGID detection method against BT (4). As a rapid and effective detection method, ELISA has been reported to be used in the clinical detection of BTV (22). In 1992, Du Plessis established an indirect ELISA method detecting anti-BTV serum (5). In 1990, House et al. developed a competitive ELISA (C-ELISA) method using monoclonal antibody (mAb) against BTV for the first time (7). The reliability of C-ELISA has also been reported by other clinical research (9).
In the present study, three segments of VP2 of BTV-8 were expressed and used as immunogen for the preparation of mAb-secreting cell lines. Finally, two mAbs that could recognize BTV-8 were obtained and characterized. Two C-ELISA methods were established by using both mAbs as competitive antibodies, which could be useful for routine examinations of field samples.
Materials and Methods
The animal protocol for this research was reviewed and approved by the Animal Care and Use Committee of Northeast Agricultural University.
Reagents, antibodies, vectors, and kits
Commercial enzymes, including T4 DNA ligase, restriction enzymes (BamHI and XhoI), DNA Ligation Kits (6022Q), and isopropyl-thio-D-galactoside (IPTG) were purchased from TaKaRa (Dalian) Co., Ltd. (Dalian, China). Escherichia coli BL21 (DE3) was used for fusion protein expression. pET-28a (+) vector was stored by our laboratory and pMAl-c5X was given by Yongning Zhang. Mouse anti-histidine (His) MAb (MA1-21315) and anti-maltose-binding protein (MBP) MAb were purchased from ZSGB-Bio Co., Ltd. (Beijing, China). The Mouse mAb Isotyping Reagent Kit was purchased from Sigma-Aldrich (St. Louis, MO).
Construction of recombinant plasmids
Full-length open reading frame of the VP2 gene of BTV-8 (GenBank accession no. AM498052.2) with BamHI and XhoI restriction sites at the 5′ end and SalI restriction site at the 3′ end was synthesized by Genewiz (Beijing) Biotech Co., Ltd. (Beijing, China). Three partial overlapping recombinant VP2 was cloned into pET-28a (+) and pMAl-c5X vectors and the positive recombinant plasmids were identified by enzyme digestion, sequencing, and named pET-28a-8A, pET-28a-8B, and pET-28a-8C and pMAl-c5X-8A, pMAl-c5X-8B, and pMAl-c5X-8C.
Expression and purification of recombinant BTV-8 VP2 protein
E. coli strains BL21 (DE3) with pET-28a-8A, pET-28a-8B, and pET-28a-8C or pMAL-c5x-8A, pMAL-c5x-8B, and pMAL-c5x-8C were grown in 200 mL of LB medium (supplemented with 50 μg/mL kanamycin or 30 μg/mL ampicillin) at 37°C to an optical density at 600 nm of ∼0.6. The protein expression was induced by adding IPTG to 1 mM final concentration. Then, additional 4 h incubation at 37°C with agitation was performed. After the cells were harvested by centrifugation at 5,000 g for 10 min at 4°C, the pelleted bacteria were suspended in 10 mL phosphate-buffered saline (PBS). Afterward, the pelleted bacteria were lysed by sonication in an ice water bath.
The recombinant protein His-8A, His-8B, and His-8C were purified from their inclusion bodies by The Ni-NTA agarose (Qiagen, Hilden, Germany). The kit was used to purify His-8A, His-8B and His-8C according to the manufacturer's instructions, with slight modifications. Briefly, the inclusion bodies were suspended with binding buffer (0.1 M NaH2PO4, 0.01 M Tris-Cl, 5 mM imidazole, 8 M urea, pH 8.0). The supernatant was transferred to a fresh tube. Two milliliters of Ni–NTA agarose pre-equilibrated with the binding buffer was added to the supernatant and shaken gently at 4°C for 1 h. The lysate–agarose mixture was loaded into an empty column, which was sequentially washed with the wash buffer (0.1 M NaH2PO4, 0.01 M Tris-Cl, 0.5 M imidazole, 8 M urea, pH 6.3). Then His-8A, His-8B, and His-8C, respectively, were eluted using 10 mL of elution buffer (0.1 M NaH2PO4, 0.01 M Tris-Cl, 8 M urea, pH 4.5).
The recombinant protein MBP-8A, MBP-8B, and MBP-8C were purified by the MBP kits. The MBP-8A, MBP-8B, and MBP-8C were purified by affinity chromatography on amylose resin following the manufacturer's protocol (New England Biolabs, Inc., Ipswich, MA). In brief, the supernatant was diluted with the column buffer (20 mM Tris–HCl, 200 mM NaCl, 1 mM EDTA, 1 mM azide, 1 mM DTT, pH 7.4) in a ratio of 1:7. The diluted supernatant was loaded slowly into a 1.6 × 5 cm column, in which the amylose resin had been equilibrated with five column volumes of the column buffer. The column was then washed with 10 column volumes of the column buffer at a rate of 4 mL/min. Finally, MBP-8A, MBP-8B, and MBP-8C were eluted with 30 mL of the column buffer supplemented with 10 mM maltose. The purified protein solutions were concentrated using centrifugal filter concentrators (10 kDa cutoff; Millipore). The concentrations of the purified recombinant protein were determined by a spectrophotometer (Thermo Scientific, Wilmington, DE).
Preparation and screening of mAbs against BTV-8 VP2 protein
To prepare mAb against BTV-8 VP2, 6-week-old BALB/c mice were intraperitoneally immunized with 50 μg purified recombinant VP2 segments expressed in E. coli and emulsified with an equal volume of Freund's complete adjuvant (Sigma-Aldrich) and boosted with the same dose emulsified with an equal volume of Freund's incomplete adjuvant 2 weeks later. Three days after the final booster, spleen cells from the immunized mice were fused with myeloma cells SP2/0, and hybridoma supernatants were screened by indirect ELISA as described below. Cells whose supernatants produced positive results in the ELISA were subcloned by limiting dilution and further characterized by western blot.
SDS-PAGE and Western blot
The samples added with His-8A/8B/8C loading buffer containing DDT were boiled for 15 min and were fractionated by sodium dodecyl–sulfate polyacrylamide gel electrophoresis (SDS-PAGE) in 12% gels, then transferred to nitrocellulose filter membrane. Then, the membrane was blocked with 5% skim milk in PBS for 2.5 h at 37°C, followed by incubation with either anti-His, anti-MBP, or prepared mAbs at 4°C overnight. After a thorough washing with phosphate-buffered saline Tween-20 (PBST), the membrane was incubated with a 1:5,000 dilution of HRP-conjugated goat anti-mouse IgG antibody for 1 h at 24°C. Color was developed with Luminata™ Forte Western HRP Substrate and the reaction was read by a ChemiScope (Clinx Science Instruments Co., Ltd, Shanghai, China).
Indirect ELISA
The microplate was coated with 100 μL of a predetermined optimal concentration (0.5 mg/well) of purified recombinant BTV8 VP2 segments incubated overnight at 4°C. After being blocked with 5% skim milk in PBS for 2 h at 37°C, the plates were incubated with 200 μL/well of hybridoma supernatants for 2 h at 37°C. Then, the plates were incubated with a 0.04 μg/well (1:2,500 dilution) of HRP-conjugated goat anti-mouse IgG antibody (Sigma) for 1 h at 37°C. A substrate solution containing o-phenylenediamine was added for color development. Following a 15-min incubation, the enzymatic reaction was stopped by the addition of 100 μL/well of 2 M H2SO4 and the absorbance (OD490 value) was measured at 490 nm using a Microplate Reader (BioTek Instruments, Inc, Winooski, VT).
Indirect Immunofluorescence
Briefly, indirect immunofluorescence assay (IFA) experiments were performed using BHK-21 cells cultured in 96-well plates. Cells were infected with BTV-8. At 48–72 h postinfection, cells were fixed using cold 75% ethanol for 30 min at 4°C. The fixed cells were then incubated with the hybridoma supernatant and positive/negative mouse serum (positive/negative control) for 1 h at 37°C. A FITC-conjugated goat anti-mouse IgG antibody (Sigma) was used as a secondary antibody at a 1:500 dilution, and plates were viewed at a magnification of 20× on a fluorescence microscope (Leica, Wetzlar, Germany).
Sandwich ELISA
Briefly, rabbit anti-BTV serum was first coated onto the wells of a 96-well microplate by incubation for 1 h at 37°C. Plates were coated in BTV of 24 distinct serotypes for 1 h and then incubated with hybridoma supernatants that were diluted 1:5 with a blocking buffer for 1 h at 37°C. The plates were washed thrice with PBST and incubated with a 1:8,000 dilution of HRP-conjugated goat anti-mouse IgG antibody (Sigma-Aldrich) in a blocking buffer for 1 h at 37°C. After a thorough washing with PBST, 3,3′,5,5′-tetramethylbenzidine (TMB) substrate solution (Becton Dickinson and Company, Franklin Lakes, NJ) was added to each well. The reaction was stopped by the addition of a half volume of 2 M H2SO4. The optical density (OD490 value) was measured at 490 nm using a Microplate Reader (BioTek Instruments, Inc.).
Optimal C-ELISA reaction conditions
The reaction conditions of C-ELISA were optimized using His/MBP-8B as the antigen, the standard anti-BTV-8-positive serum, and the mouse mAb as the competitive antibodies. The microplate was coated with 1, 2, 5, or 10 μg/well of His/MBP-8B for 1, 2, or 3 h. After being blocked with 5% skim milk in PBS for 2 h at 37°C, the plate was incubated with a mixture of standard anti-BTV-8-positive serum (diluted at 1:10, 1:20, 1:50, 1:100, or 1:200) and the mAb (diluted at 1:500, 1:1,000, 1:2,000, or 1:4,000) for 1, 2, or 3 h at 37°C. Then, color was developed using TMB for 7, 10, or 15 min. The optimal conditions of C-ELISA were determined according to the outcomes of the reactions with different reagent concentrations.
Determination of percent of inhibition
Percent of inhibition (PI) = (12) × 100% (ODmax: OD value of the empty control, ODx: OD value of serum samples, ODmin: OD value of negative control). The mean value and standard deviation (SD) of 50 serum samples were calculated, +2SD was designated as the cutoff value. Samples with PI>+ 2SD were considered as positive samples.
Specificity, sensitivity, and repeatability of C-ELISA
The cross-reaction with other BTV serotypes was detected by using 10 positive and 10 negative serum samples of each BTV serotype (diluted at 1:20) in C-ELISA. The standard anti-BTV-8-positive serum was diluted at 1:5, 1:10, 1:20, 1:40, 1:80, 1:160, and 1:320 and used as the competitive antibody in C-ELISA. Ten serum samples were tested by C-ELISA, each one of which was performed three times. Then, the repeatability of C-ELISA was assessed.
Field samples
Seventy serum samples of sheep, goat, and cattle from two farms were detected by the BT Antibody ELISA Kit (Jiya Biotech Co., Ltd. Shanghai, China), after which the positive samples were detected by C-ELISA developed in this article.
Results
Expression and purification of recombination protein
After SDS-PAGE analysis and Coomassie Brilliant Blue staining, thick protein bands with molecular weight around 51, 47, and 46 kDa were detected in IPTG-induced pET-28a-8A, pET-28a-8B, and pET-28a-8C-transformed E. coli BL21 (DE3) (Fig. 1A). Moreover, thick protein bands with molecular weight around 87 kDa, 82 kDa, and 81 kDa were also detected in IPTG-induced pMAL-c5x-transformed E. coli BL21 (DE3) (Fig. 1B).
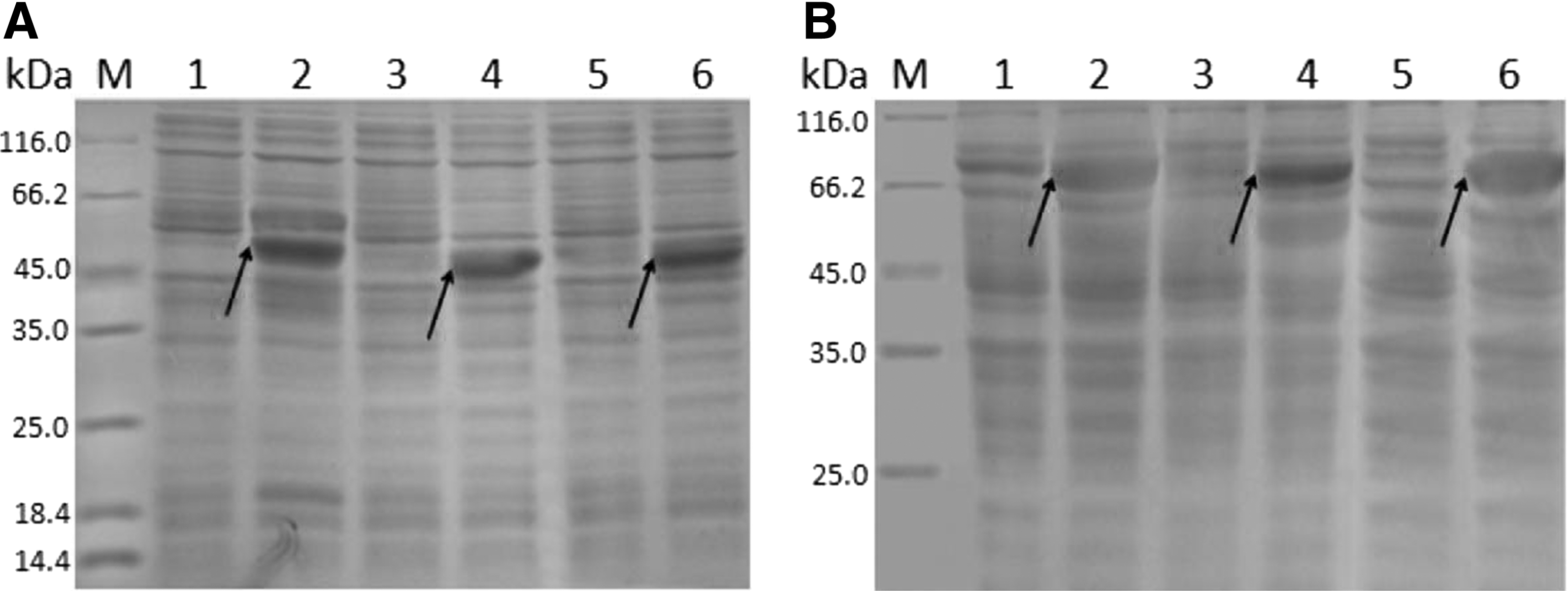
SDS-PAGE analysis of recombinant BTV-8 VP2 segments expressed in Escherichia coli BL21 (DE3).
His-8A/8B/8C were expressed mainly in the inclusion bodies form, the precipitation of whole-cell lysates from IPTG-induced bacteria was used for protein purification using Ni–NTA agarose. The concentrations of purified His-8A/8B/8C were 0.54, 0.66, and 0.58 mg/mL. MBP-8A/8B/8C were expressed mainly in soluble form, the supernatants were used for protein purification using amylose resin affinity chromatography. The concentration of purified MBP-8A/8B/8C was 0.61, 0.55, and 0.58 mg/mL. Western blot analysis indicated that both His-8A/8B/8C and MBP-8A/8B/8C were specifically reactive with anti-His antibody and anti-MBP antibody, respectively (Fig. 2).
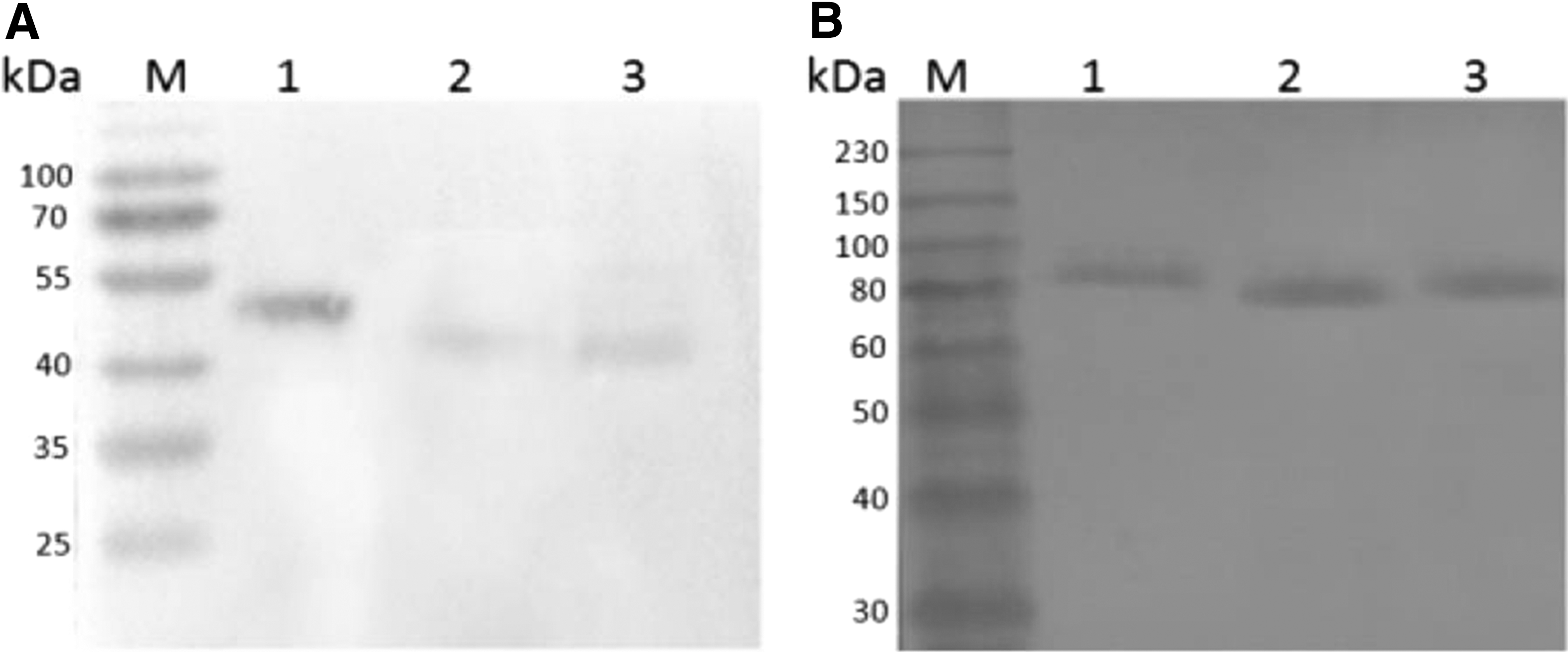
Western blot identification of the purified recombinant His-8A/8B/8C and MBP-8A/8B/8C.
Preparation and identification of VP2-specific mAbs
Among the BALB/c mice immunized with purified recombinant His-8A/8B/8C, respectively, the mouse with the highest antibody titer was selected for cell fusion. Ten hybridoma cell lines stably secreting mAbs against the BTV-8A/8B/8C protein were obtained.
To characterize the reactivity of the mAbs with VP2 segments, western blot analysis was correspondingly performed using 10 mAbs as the primary antibody. The mAbs were all shown to react with His-8A, His-8B, or His-8C (Fig. 3). Double antibody sandwich ELISA results showed that 8B-5D4 and 8B-3G11 could recognize BTV-8, whereas other mAbs did not have reaction with BTV-8 (data not shown). In addition, the IFA results confirmed that mAb 8B-5D4 and 8B-3G11 had positive reactions with BHK-21 cells infected by BTV-8 (Fig. 4). The results of antibody isotyping showed that 8B-5D4 and 8B-3G11 belonged to IgG2a and IgG2b, respectively.
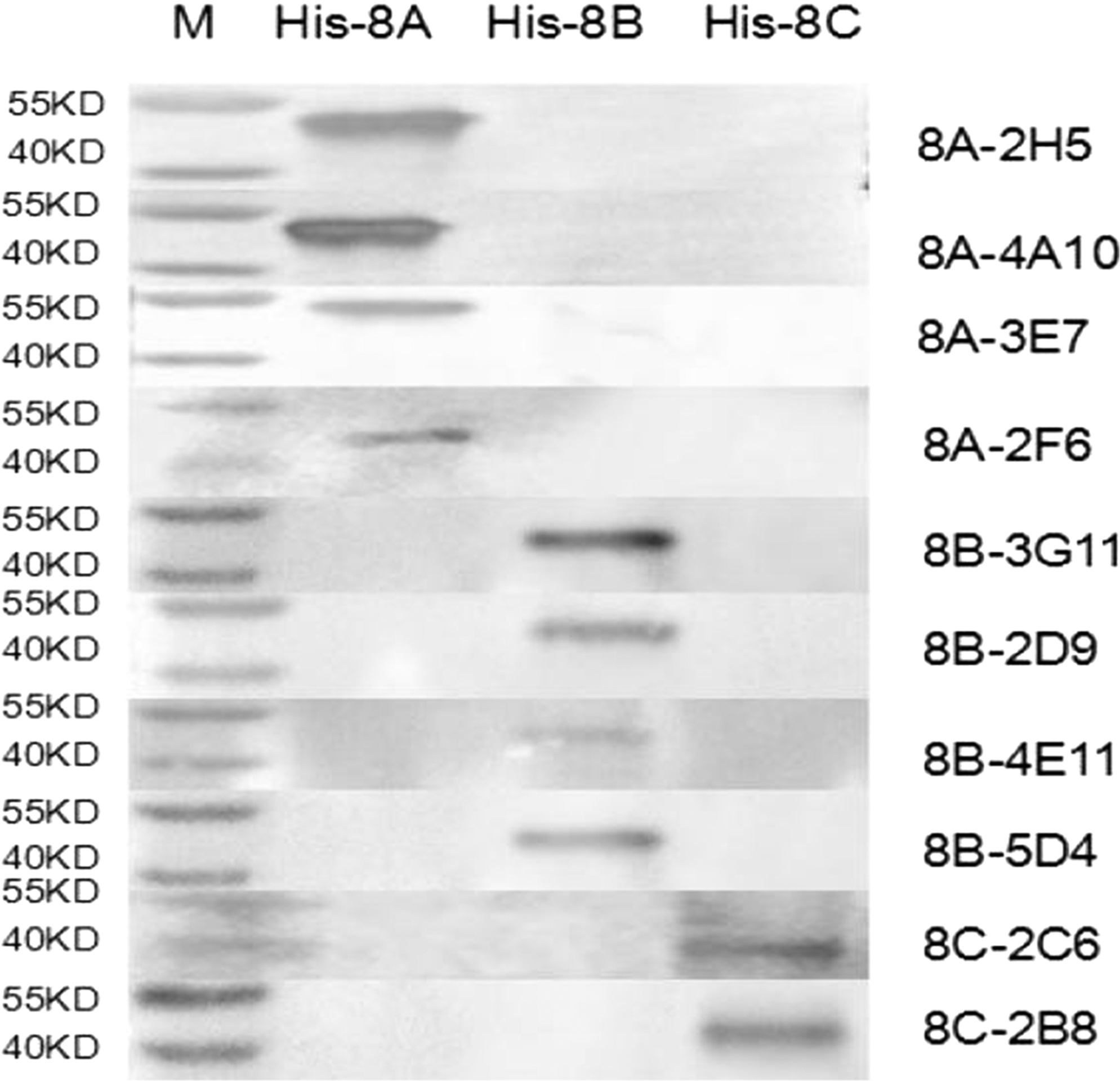
Western blot identification of mAbs with His-8A, His-8B, and His-8C protein.
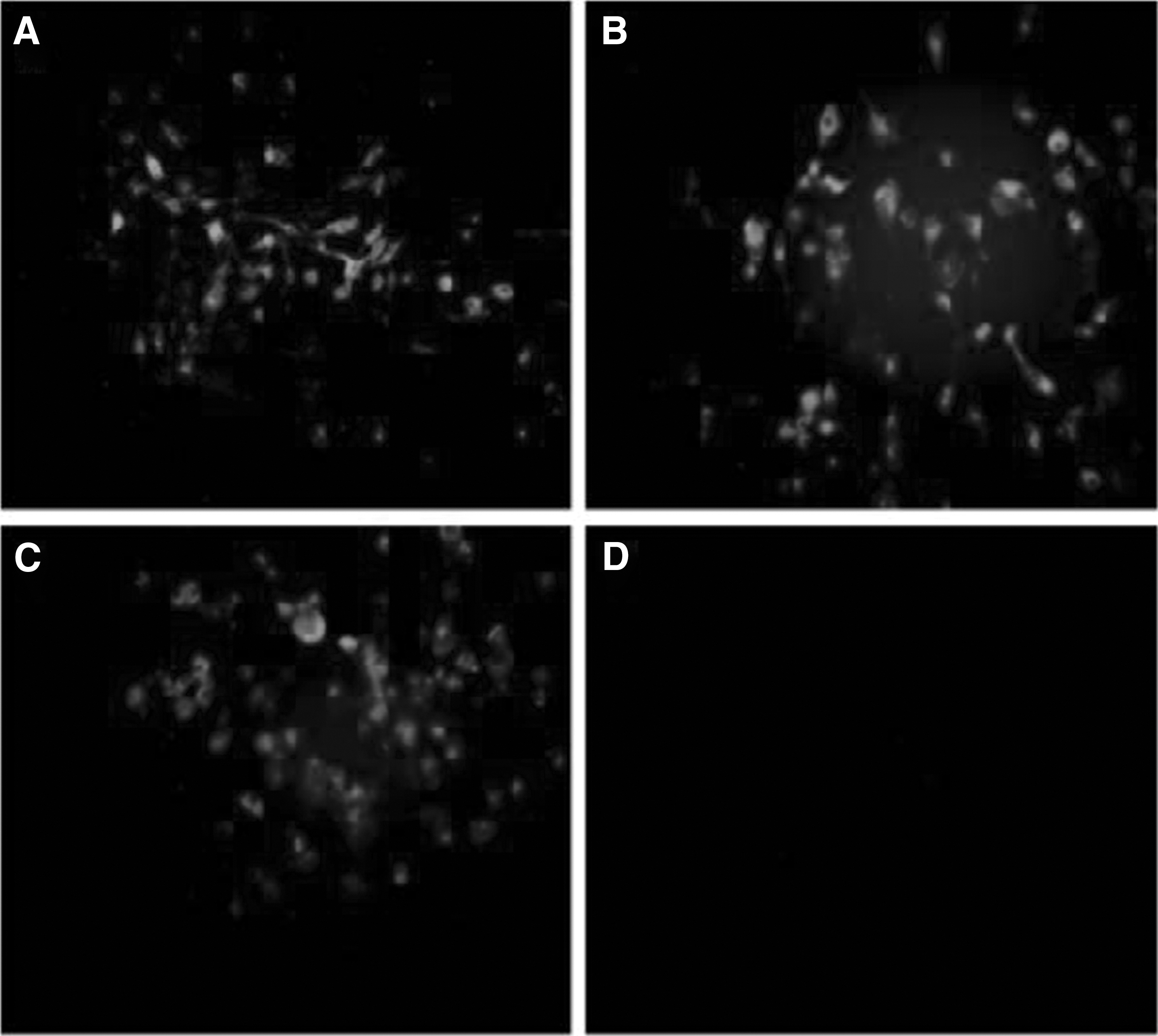
Reactivity of mAbs with BTV serotypes 8-infected BHK-21 cells by IFA.
Optimal conditions of C-ELISA
C-ELISA by checkerboard titration was performed to determine the optimal conditions for test samples. His/MBP-8B of 5 μg/well coated at 37°C for 1 h, sample serum dilution of 1:20, mAb dilution of 1:1,000 and 1:2,000 for 8B-3G11 and 8B-5D4, incubation at 37°C for 2 h, and color development of 10 min with TMB of 100 μL/well were selected as the optimal conditions (Tables 1 and 2).
BTV, Bluetongue virus; C-ELISA, competitive enzyme-linked immunosorbent assays; TMB, 3,3′,5,5′-tetramethylbenzidine.
Determination of PI value
Fifty negative anti-BTV serum samples were detected by C-ELISA of 8B-3G11 and 8B-5D4 to determine the PI. The PI value of 8B-3G11 and 8B-5D4 were 44% and 49% (Figs. 5 and 6).
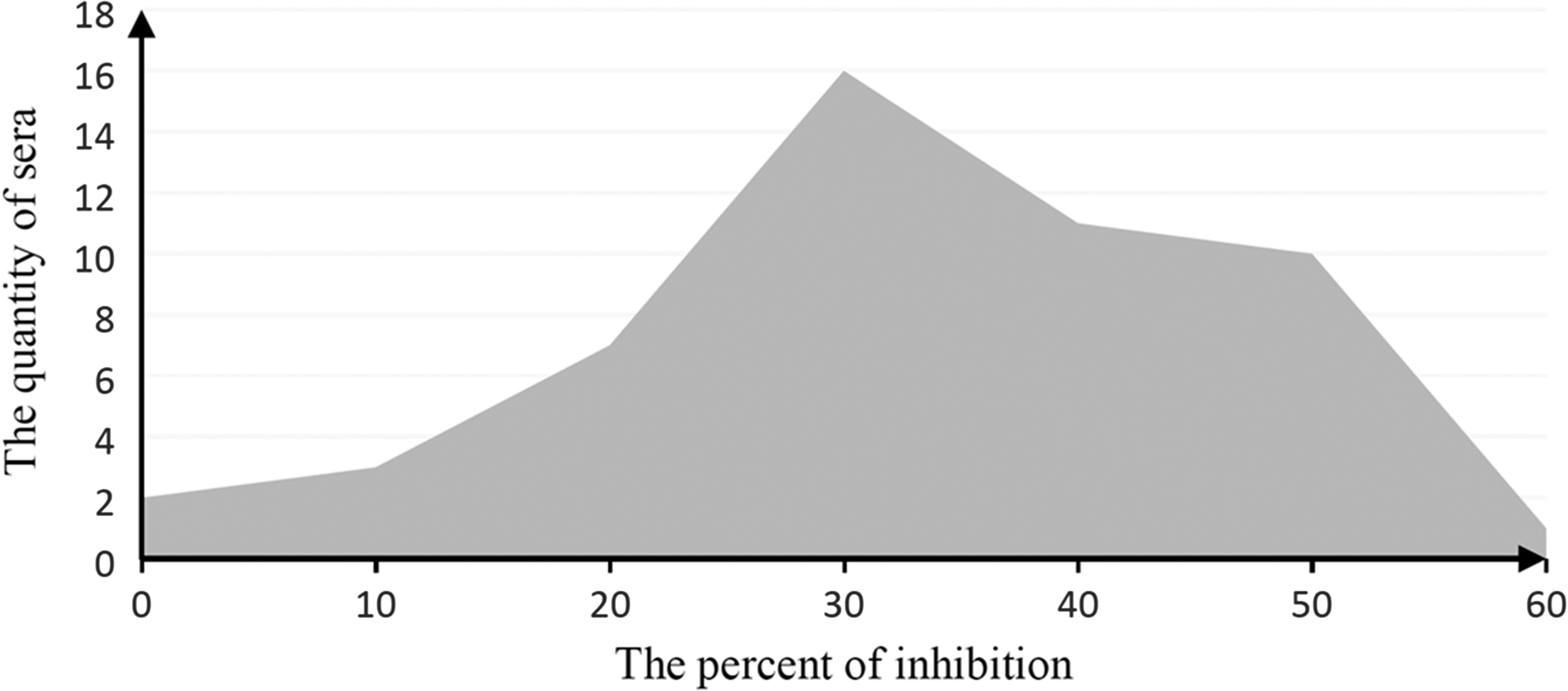
The assay of negative serum samples of 8B-5D4 C-ELISA. C-ELISA, competitive enzyme-linked immunosorbent assays.
The assay of negative serum samples of 8B-3G11 C-ELISA.
Specificity, sensitivity, and repeatability of C-ELISA
Specificity results showed that 8B-3G11 C-ELISA cross-reacted with anti-BTV-7 and anti-BTV-10 sera, whereas 8B-5D4 C-ELISA crossreacted with anti-BTV-1, -4, -6, -7, -14, -18, -19, -21, and -23 sera (Table 3). The standard anti-BTV-8 serum was diluted from 1:5 to 1:320, the sensitivity of 8B-3G11 C-ELISA and 8B-5D4 C-ELISA were determined to be 1:80 and 1:160. Ten serum samples were tested by both C-ELISA methods from three different batches, the results of which showed insignificant difference with the variable coefficient during 4–10%.
Field samples
Among the 20 positive samples detected by BT the group-specific ELISA Kit, one positive sample was confirmed by 8B-3G11 C-ELISA, whereas four positive samples were confirmed by 8B-5D4 C-ELISA, including the one confirmed by 8B-3G11 C-ELISA.
Discussion
BT is an insect transmission viral disease of domestic and wild ruminants, especially sheep. It is characterized by inflammation, hemorrhage, ulceration, and cyanosis of the mucous membranes, laminitis, coronitis, and edema of the head and neck (1). Twenty-six distinct serotypes of BTV have been detected around the world, between latitudes in the Americas, Africa, Asia, Australia, and Europe (11). Due to the limited crossprotection between different serotypes, the effectivity of BTV vaccine is unsatisfactory. Therefore, developing serological diagnosis methods against different BTV serotypes is an important way for BT prevention. Compared with indirect immunofluorescence assay and neutralization test that need to be performed in P3-level laboratory, real-time PCR method of high cost, and AGID method of low specificity, C-ELISA method based on mAbs has been considered as the optimal method for its low cost, high specificity, and simplicity of performance.
In the present study, His-8A/8B/8C were used as immunogen for the preparation of mAb against BTV-8, and two mAbs (8B-5D4 and 8B-3G11) that could recognize BTV-8 were obtained. As a mature operating system, prokaryotic expression system is widely used in diagnostic reagent preparation, protein function research, and production of subunit vaccine. To retain the antigenic and immunogenic properties of His-8A/8B/8C, and to overcome protein refolding issues, various induction conditions, including IPTG concentration, induction temperature, and induction time were optimized (data not shown). Ultimately, inducing of the bacteria with 1 mM IPTG for 4 h at 37°C was determined as the optimal conditions for His-8A/8B/8C induction. To avoid screening hybridoma cells that secrete mAbs against the His tag, MBP-8A/8B/8C were expressed, purified, and then used as coating antigens in ELISAs to screen for positive hybridomas specific for the VP2 segments.
To establish the C-ELISA method, mAbs 8B-5D4 and 8B-3G11, respectively, were used as the competitive antibody. To improve the sensitivity and specificity, several reaction conditions were optimized, including the concentration of coating antigen, dilutions of sample serum and mAb, reaction temperature, and time. Due to the homology among VP2 proteins of different BTV serotypes, 8B-5D4 C-ELISA could screen out anti-BTV-1, -4, -6, -7, -8, -14, -18, -19, -21, and -23 sera, whereas 8B-3G11 C-ELISA could screen out anti-BTV-7, -8, and -10 sera. Our additional research found that the epitope recognized by 8B-5D4, 192LKP194, exists in BTV-1, -6, -7, -14, -18, -19, -21, and -23 as well, and the epitope recognized by 8B-3G11, 283LL284, exists in BTV-7, which further explains the reason of the crossreaction of both C-ELISA methods. However, with the joint reaction of the two methods, serum samples containing anti-BTV-7 or anti-BTV-8 antibody could be screened out.
In conclusion, mAbs 8B-5D4 and 8B-3G11 that could recognize BTV-8 were produced. Two C-ELISA methods based on the mAbs were established, which are suitable for screening a large number of samples. This research provides us with useful tools that can be implemented in the diagnosis and serological study of BT.
Footnotes
Acknowledgments
This study was supported by the National Key Technology R&D Program of China (Grant no. 2013BAD12B01). The authors gratefully thank Dr. Jianbo Zhu from Yunnan Animal Science and Veterinary Institute for the specificity testing of mAbs with 24 serotypes of BTV.
Author Disclosure Statement
No competing financial interests exist.