
Abstract
Toll-like receptor (TLR)3 signaling pathway is known to induce type 1 interferons (IFNs) and proinflammatory mediators leading to antiviral response against many viral infections. Double-stranded ribonucleic acid (dsRNA) has been shown to act as a ligand for TLR3 and, as such, has been a focus as a potential antiviral agent in many host-viral infection models. Yet, its effectiveness and involved mechanisms as a mediator against low pathogenic avian influenza virus (LPAIV) have not been investigated adequately. In this study, we used avian fibroblasts to verify whether dsRNA induces antiviral response against H4N6 LPAIV and clarify whether type 1 IFNs and proinflammatory mediators such as interleukin (IL)-1β are contributing to the dsRNA-mediated antiviral response against H4N6 LPAIV. We found that dsRNA induces antiviral response in avian fibroblasts against H4N6 LPAIV infection. The treatment of avian fibroblasts with dsRNA increases the expressions of TLR3, IFN-α, IFN-β, and IL-1β. We also confirmed that this antiviral response elicited against H4N6 LPAIV infection correlates, but is not attributable to type 1 IFNs or IL-1β. Our findings imply that the TLR3 ligand, dsRNA, can elicit antiviral response in avian fibroblasts against LPAIV infection, highlighting potential value of dsRNA as an antiviral agent against LPAIV infections. However, further investigations are required to determine the potential role of other innate immune mediators or combination of the tested cytokines in the dsRNA-mediated antiviral response against H4N6 LPAIV infection.
Introduction
A
Although various control measures, including biosecurity, surveillance, and mass euthanasia of affected and in contact poultry, are practiced (85), transmission of AIV to public time to time is a continuous challenge (11,78). Vaccination against AIV also has been introduced in some developing countries, but without a success (78) since, these vaccines are less efficacious, vaccination does not prevent the infection of the vaccinated birds and cross protection between subtypes of AIV are weak (12,30,70). Moreover, immune pressure may lead to selection of mutations in the AIV genome leading to novel strains (9,13,14,21,50). Novel strains may not be vulnerable to vaccine-induced immune responses as such changes in vaccine composition often required (1,27,45,50,65).
Similar to mammals, avian immune system consists of two arms: innate and adaptive. Innate host responses elicited based on detection of highly conserved pathogen-associated molecular patterns. Such patterns encoded by the pathogens such as double-stranded ribonucleic acid (dsRNA) are detected by pattern recognition receptors, particularly by toll-like receptor (TLR)3 (54).
Polyinosinic–polycytidylic acid [Poly(I:C)] is a synthetic analog of dsRNA and, as such, could act as a ligand for TLR3, which is expressed on the endosomal membrane of host cells (82). This binding of dsRNA with TLR3 (49) can activate toll-interleukin-1 (IL-1) receptor (TIR) domain-containing adaptor-inducing interferon β (TRIF) pathway, leading to the production of type 1 interferons (IFNs). Two other receptors expressed in the cytosol as well recognize dsRNA: the retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) and melanoma differentiation-associated gene 5 (MDA5). Following detecting intracellular dsRNA, the transcription factors such as nuclear factor (NF)-κB and IFN regulatory factor (IRF)3 are induced, leading to the production of type 1 IFNs and proinflammatory cytokines (47).
IFN-α and IFN-β are members of the type 1 IFN family and signal through binding to cell surface receptors with two subunits known as IFN alpha receptor (IFNAR)1 and IFNAR2 expressed on most of the host cells (17,20,75) to elicit antiviral responses as one of the functions. The binding of the type 1 IFNs to these subunits activates the Janus Kinase and Signal Transducer and Activator of Transcription (JAK-STAT) pathway, inducing the transcription of IFN-stimulated genes (ISGs) (71). ISGs are the primary effector molecules of type 1 IFN signaling pathway that functions to elicit antiviral responses (19,72). Although both members of the family bind to the common receptor subunits, their downstream activities, for example antiviral activities, are not similar (10,23,25,40,49).
TLR3 signaling plays a decisive role in the outcome of a variety of viral infections in vitro and in vivo (28,67,69,79,81). In chickens, TLR3 signaling dependent antiviral response has been shown against Newcastle disease virus (NCDV), where overexpression of chicken TLR3 in Henrietta Lacks (Hela) cells and chicken fibroblasts has been shown to decrease NCDV titers (16). RB1B strain of Marek's disease virus (MDV) replication has been shown to be reduced in chicken embryo fibroblasts (CEFs) treated with dsRNA (37). Intramuscular delivery of dsRNA in chickens has also been shown to decrease cloacal and oropharyngeal shedding of LPAIV (76).
Also, it has been shown that dsRNA can increase mRNA expression of type 1 IFNs in various cells (22,42,76,80), as well as ISGs in primary B cells (4). Furthermore, recombinant IFN-α has been shown to inhibit exogenous avian leukosis virus (18) and AIV (39) in vitro. dsRNA-induced IFN-β mRNA expression has been shown to be abrogated when small interfering RNA has been used to reduce the mRNA expression of TLR3 (42).
However, it is unknown whether dsRNA-mediated antiviral response against LPAIV infection correlates with the expression of TLR3, type 1 IFNs, and IL-1β. We hypothesized that these mediators are associated with dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblast cells. The objective of our study was to uncover the mechanism of dsRNA-mediated induction of antiviral response in avian fibroblasts against H4N6 LPAIV infection. Our findings imply that dsRNA has antiviral activity against H4N6 LPAIV infection in avian fibroblasts correlating with the increased expression of type IFNs and IL-1β, although the antiviral response is not attributable to type 1 IFNs or IL-1β.
Materials and Methods
Eggs and embryos
The use of 9–11 day old specific pathogen-free (SPF) chicken eggs for the purpose of propagating H4N6 LPAIV and making CEFs was approved by the Health Science Animal Care Committee (HSACC). The SPF eggs were purchased from the Canadian Food Inspection Agency (CFIA), Ottawa, Canada, and incubated at Health Research Innovation Center (HRIC), University of Calgary, in egg incubators (Rcom Pro 20 and 50, Kingsuromax 20 and Rcom MARU Deluxe max; Autoelex Co., Ltd., GimHae, GyeongNam, Korea) until the embryos were 9–11 days old.
Viruses and reagents
H4N6 LPAIV (A/Duck/Czech/56) was obtained from Dr. Eva Nagy (University of Guelph, Canada), propagated in embryo day (ED) 9–11 SPF eggs, and titrated on monolayers of Madin-Darby Canine Kidney (MDCK) cells. The vesicular stomatitis virus encoded with green fluorescent protein (VSV-GFP) was provided by Dr. Markus Czub (University of Calgary, Canada). The VSV-GFP was propagated and titrated on African green monkey kidney (Vero) cells.
The ligand for TLR3, synthetic dsRNA analogue Poly(I:C) HMW (large dsRNA strands of the sizes 1.5–8 kbp) VacciGrade™ was purchased from InvivoGen (San Diego, CA) and reconstituted as prescribed by the manufacturer. Tumor necrosis factor (TNF) receptor-associated factor (TRAF) family member-associated NF-κB activator (TANK)-binding kinase 1 (TBK1) inhibitor, BX 795, which was used for blocking IRF3 hence type 1 IFN production, was also purchased from InvivoGen. Chicken IL-1 receptor antagonist (IL-1Ra) was purchased from Kingfisher Biotech, Inc., MN, USA. It is known that IL-1Ra binds to IL-1 receptor and prevents receptor engagement of IL-1β (5).
Cells and cell culture
The Douglas Foster (DF)-1 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA). The DF-1 cells are free of endogenous retrovirus sequences since they have been immortalized spontaneously (35). DF-1 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco; Life Technologies, Burlington, ON, Canada) supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL), and maintained in a 5% CO2 incubator at 40°C.
Vero and MDCK cell lines were purchased from ATCC and cultured in DMEM supplemented with 10% FBS, penicillin (100 U/mL), and streptomycin (100 μg/mL), and maintained in a 5% CO2 incubator at 40°C.
Primary CEFs were isolated from ED 9–11 eggs following a standard protocol. Briefly, SPF eggs at ED 9–11 were candled to isolate embryos. The embryos without the head, viscera, and limbs were minced and washed with sterile phosphate-buffered saline (PBS) to remove residual blood. The minced tissue was trypsinized using TrypLE™ Express Enzyme (1 × ) (Gibco; Life Technologies) for 5 min at 37°C. The cells were sent through 70 μm cell strainer (Sigma, Saint Louis, MO) into a centrifuge flasks containing 5% FBS. The trypsinization was repeated three to four times until adequate cells were obtained. The collected cells were centrifuged for 10 min at 350 g. The supernatant was discarded and the pellet was suspended in 100 mL of DMEM supplemented with 10% FBS, penicillin (100 U/mL), and streptomycin (100 μg/mL).
The cell numbers were quantified and viability of the cells was determined using trypan blue dye exclusion. The cells were seeded at 1 × 106 cells/mL in tissue culture flasks and maintained in a 5% CO2 incubator at 40°C.
Experimental design
All our experiments were performed in vitro using avian fibroblasts. AIVs are known to infect avian fibroblasts (51) and these cells have been used commonly to study avian type 1 IFN signaling pathways (26,52,77).
Determination whether dsRNA mediates antiviral response against H4N6 LPAIV infection in avian fibroblasts
The DF-1 and primary CEFs were cultured in 12-well plates at a cell density of 5 × 105 cells/well for 24 h. Then the cells were stimulated with dsRNA in Opti-minimum essential medium (MEM) I reduced serum medium (Gibco; Life Technologies) at a concentration of 50 μg/mL (86), along with a control that received the same media alone, and the plates were incubated in a 5% CO2 incubator at 40°C.
After 24 h, the confluent monolayer of cells in 12-well plates were washed with warm Hank's balanced salt solution (HBSS) and inoculated with H4N6 LPAIV at a multiplicity of infection (MOI) of 0.1. After 30 min of incubation at 37°C, the inoculum was removed, washed with warm HBSS, fresh Opti-MEM I reduced serum medium was added, and the plates were incubated for further 24 h in a 5% CO2 incubator at 40°C. The H4N6 LPAIV infections of DF-1 and primary CEF were determined by plaque assay done on MDCK cells, as has been described previously (2).
Determination whether dsRNA increases TLR3 expression in avian fibroblasts
DF-1 cells were cultured in six-well plates at a concentration of 1 × 106 cells/well for 24 h in a 5% CO2 incubator at 40°C. The cells were treated either with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) or only with the medium as a control, and all the plates were incubated for 24 h in a 5% CO2 incubator at 40°C. For the immunostaining, we employed the same procedure described for staining for IFN-α and IFN-β, except the primary antibody, which was rabbit anti-chicken TLR3 polyclonal antibody (Creative Diagnostics, Shirley, NY) diluted at 1:200 in the blocking buffer.
Determination whether dsRNA increases IFN-α and IFN-β expressions in avian fibroblasts
DF-1 cells were cultured in six-well plates at a concentration of 1 × 106 cells/well for 24 h in a 5% CO2 incubator at 40°C. The cells were treated either with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) or only with the medium as a control. After 6 h, for the purpose of blocking secretion of type 1 IFNs, ebioscience™ protein transport inhibitor cocktail 500 × (Invitrogen Life Technologies, Carlsbad, CA) was added at the rate of 2 μL/mL for all the wells and were incubated for further 18 h in a 5% CO2 incubator at 40°C. For the immunostaining of type 1 IFNs, the plates were washed once with HBSS, fixed with 4% paraformaldehyde for 20 min, washed thrice with HBSS, permeabilized with 0.2% Triton 1 × for 30 min, washed thrice with HBSS, and blocked with 5% horse serum for 1 h at the room temperature.
The used primary antibodies were rabbit anti-chicken IFN-β (Bio-Rad Laboratories, Mississauga, ON, Canada), rabbit anti-chicken IFN-α (Bio-Rad Laboratories), or isotype control (rabbit IgG), and were diluted at 1:10 in the blocking buffer and incubated for 1 h at room temperature. For the secondary antibody, VectaFluor™ Excel Amplified DyLight® 594 Anti-Rabbit IgG Kit (Vector Laboratories, Inc., Burlingame, CA) was used following the manufacturer's instructions. Nuclear staining of the cells was done with Hoechst 33342 (Image-iT™ LIVE Plasma Membrane and Nuclear Labeling Kit [I34406]; Invitrogen, Eugene, OR) following the manufacturer's instruction. Plates were read and analyzed using an epifluorescent microscope (Olympus IX51, Markham, Ontario, Canada).
Determination whether IFN-β expression is enhanced during dsRNA-mediated antiviral response against H4N6 LPAIV replication in avian fibroblasts
DF-1 cells were cultured in two 12-well plates at a concentration of 0.2 × 106 cells/well for 24 h and incubated in a 5% CO2 incubator at 40°C. The cells were treated either with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) (n = 3) or only with the growth medium as a control (n = 3). The plates were incubated for further 24 h in a 5% CO2 incubator at 40°C. The wells from each group were infected with H4N6 LPAIV at MOI = 0.1 for 30 min at 37°C, 1 mL of Opti-MEM I reduced serum medium was added after washing the inoculum out, and incubated for 6 h in a 5% CO2 incubator at 40°C, while maintaining the rest as uninfected controls. After 6 h of incubation, for the purpose of blocking secretion of IFN-β, eBioscience protein transport inhibitor cocktail 500 × (Invitrogen Life Technologies) was added at the rate of 2 μL/mL for all the wells.
For the immunostaining of IFN-β, after further 18 h, the plates were washed once with HBSS, fixed with 4% paraformaldehyde for 20 min, and proceeded with immunofluorescent staining as described in the previous section.
Determination whether the dsRNA-induced antiviral response against H4N6 LPAIV is attributable to type 1 IFNs
First, to determine whether TBK1 inhibitor is able to inhibit type 1 IFN activity downstream of dsRNA-TLR3 signaling, DF-1 cells were seeded in 12-well plates at a concentration of 0.2 × 106 cells/well for 24 h in a 5% CO2 incubator at 40°C.
Three wells of cells were first treated with TBK1 inhibitor (BX 795) at 4 μM/well for 2 h employing three wells of untreated controls. TBK1 plays a role in the dimerization and translocation of IRF3 (24), hence production of type 1 IFNs downstream of TLR3-dsRNA signaling. After the inhibition, both groups of cells were treated with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies). Three additional sets of controls such as the growth medium as a control, DMSO, or BX 795 were also employed and the plates were incubated for 24 h in a 5% CO2 incubator at 40°C. All the cells were infected with VSV-GFP at an MOI of 0.1 for 30 min at 37°C with uninfected controls, and then fresh Opti-MEM I reduced serum medium was added to the cells and were incubated for 24 h in a 5% CO2 incubator at 40°C.
All the plates were washed twice with HBSS and fixed with 4% paraformaldehyde for 20 min at room temperature, washed again with HBSS, and nuclear stained with DAPI. To determine the percentages of cells infected with VSV-GFP, the plates were scanned using In Cell Analyzer 2000 live C TEMP/LH/EC high light analysis system (GE Healthcare Sciences, Mississauga, Ontario, Canada).
Second, to determine whether the expression of IFN-β is inhibited following TBK1 treatment of avian fibroblasts downstream of dsRNA-TLR3 signaling, DF-1 cells were seeded in 12-well plates at a concentration of 0.2 × 106 cells/well and incubated in a 5% CO2 incubator at 40°C for 24 h. Six wells were treated with TBK1 inhibitor (BX 795) at 4 μM/well for 2 h. Then, one group of cells was treated with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) (n = 3, TBK1 inhibitor treated; n = 3, without TBK1 inhibitor) and the other groups acted as the controls (n = 3, TBK1 inhibitor alone; n = 3 without TBK1 inhibitor). The plates were fixed and immunofluorescent assay was performed for determination of expression of IFN-β as described earlier.
Third, to determine whether dsRNA induced antiviral response against H4N6 LPAIV in avian fibroblasts is attributable to type 1 IFNs, DF-1 cells were seeded in 12-well plates at a concentration of 0.2 × 106 cells/well for 24 h in a 5% CO2 incubator at 40°C. Six wells of cells were first treated with TBK1 inhibitor (BX 795) at 4 μM/well for 2 h employing six wells as untreated controls. After the type 1 IFN production inhibition step, three wells from each group of cells were treated with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies). After 24 h, all the wells were infected with H4N6 LPAIV at MOI = 0.1 for 30 min at 37°C, 1 mL of Opti-MEM I reduced serum medium was added after washing the inoculum out, and incubated for 24 h in a 5% CO2 incubator at 40°C, while maintaining uninfected controls.
Twenty-four hours postinfection, the supernatants were collected and the plaque assay was done on MDCK cells to enumerate number of plaque produced by H4N6 LPAIV.
Determination whether dsRNA induces IL-1β expression in avian fibroblasts
The DF-1 cells were cultured on coverslips in 12-well plates with 0.2 × 106 cells/well and protein transport inhibitor (2 μL/mL) (Invitrogen Life Technologies) was added to culture medium after 6 h. After 24 h of culture, the cells were stimulated with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) or only with the growth medium as a control (nine replicates per treatment). Following 24 h of treatment, immunofluorescent staining was performed after fixation with 4% paraformaldehyde (three well per treatment). In immunofluorescent staining, unlabeled rabbit polyclonal antibody specific for chicken IL-1β (Bio-Rad Laboratories) was used as a primary antibody in 1:10 dilutions and biotinylated goat anti-rabbit IgG(H+L) (Vectashield; Vector Laboratories Inc.) was used as the secondary antibody in 1:400 dilution.
Determination whether dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts is attributable to IL-1β expression
DF-1 cells were cultured in 12-well plates at a concentration of 0.2 × 106 cells/well for 24 h and incubated in a 5% CO2 incubator at 40°C. The cells were treated either with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium (Gibco; Life Technologies) (n = 6) or only with the growth medium as a control (n = 6). The plates were incubated for further 24 h in a 5% CO2 incubator at 40°C. The culture supernatants were collected and transferred (450 μL) to another DF-1 cell monolayer in a 12-well plate. Before being treated with cell culture supernatants, six wells were incubated with 1.0 μg/mL of IL-1Ra (Kingfisher Biotech, Inc.) for 30 min, while maintaining the six wells as controls. Following 24 h, all wells in the second DF-1 cell monolayer were infected with H4N6 LPAIV at MOI = 0.1 maintaining uninfected controls.
At 24 h postinfection, the culture supernatants were collected and transferred to MDCK cell monolayer in 10-fold serial dilution. The resulting plaques were counted after staining with 1% crystal violet at 48 h postinfection.
Data analyses
Analyses of immunofluorescent signals
To count the percentage of cells expressing VSV-GFP, the plates were scanned with the In Cell Analyzer 2000 live C TEMP/LH/EC high light analysis system (GE Healthcare Sciences), which captured pictures of 12–24 fields from each well. The images were then analyzed through multitarget analysis using In Cell Analyzer workstation 1000 software version 3.7 (GE Healthcare Sciences). To quantify the fluorescent intensity and % positive cells, five fields/well were captured using an epifluorescent microscope and analyzed by ImageJ software (National Institute of Health, Bethesda, MD).
Statistical analysis
Student's t-test (GraphPad Prism Software 5, La Jolla, CA) was used for identifying the differences between two groups. When multiple groups were present in an experiment, one-way ANOVA followed by Tukey's test was used to compare group differences. The differences between groups were considered significant when p < 0.05.
Results
TLR3 ligand, dsRNA elicits antiviral response against H4N6 LPAIV infection in avian fibroblasts
The culture supernatants collected from DF-1 cells or CEFs following 24 h of dsRNA or PBS treatment, followed by 24 h of H4N6 LPAIV infection, were titered in MDCK cells. Briefly, DF-1 and CEFs were cultured for 24 h before being treated with dsRNA along with a control. After 24 h of treatment, the culture supernatants were collected and transferred on to MDCK cells. After adsorption of the virus for 30 min at 37°C, the inoculum was removed and washed with warm HBSS. After 30 min of incubation at 37°C, the plates were overlaid with serum-free 2 × MEM media containing the equal volume of 2.4% avicel® (FMC BioPolymer, Philadelphia, PA) and 1 μg/mL of bovine l-tosylamide-2-phenylethyl chloromethyl ketone (TPCK)-trypsin (Sigma). The inoculated plates were incubated for 2 days at 37°C and 5% CO2.
The visible plaques were counted under an inverted microscope after staining with 1% crystal violet. We observed that dsRNA reduces H4N6 LPAIV replication in both DF-1 cells (Fig. 1a, p < 0.0001) and CEFs (Fig. 1b, p = 0.0489). The extent of inhibition in DF-1 cells was more pronounced in DF-1 cells, which is a cell line when compared to that observed in primary cells (CEFs).
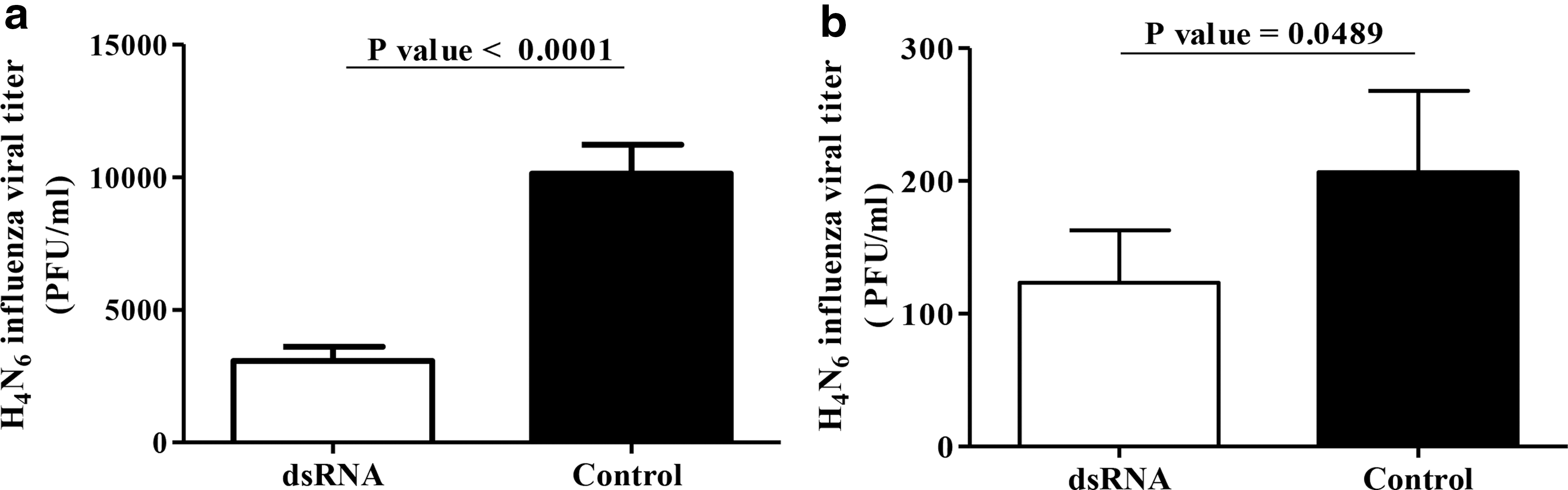
TLR3 ligand, dsRNA, induces antiviral response against H4N6 LPAIV infection in avian fibroblasts.
TLR3 ligand, dsRNA increases TLR3 expression in avian fibroblasts
Since dsRNA mediates antiviral response by type 1 IFN production signaling through TLR3, we immunoassayed the expression of TLR3 following dsRNA or control treatments and observed that dsRNA treatment of avian fibroblasts leads to significantly higher expression of TLR3 in avian fibroblasts as assessed by fluorescent intensity (Fig. 2a, b, p = 0.0018). The % of cells expressing TLR3 was also higher in dsRNA-treated fibroblasts when compared to the controls (Fig. 2c, p = 0.0451).
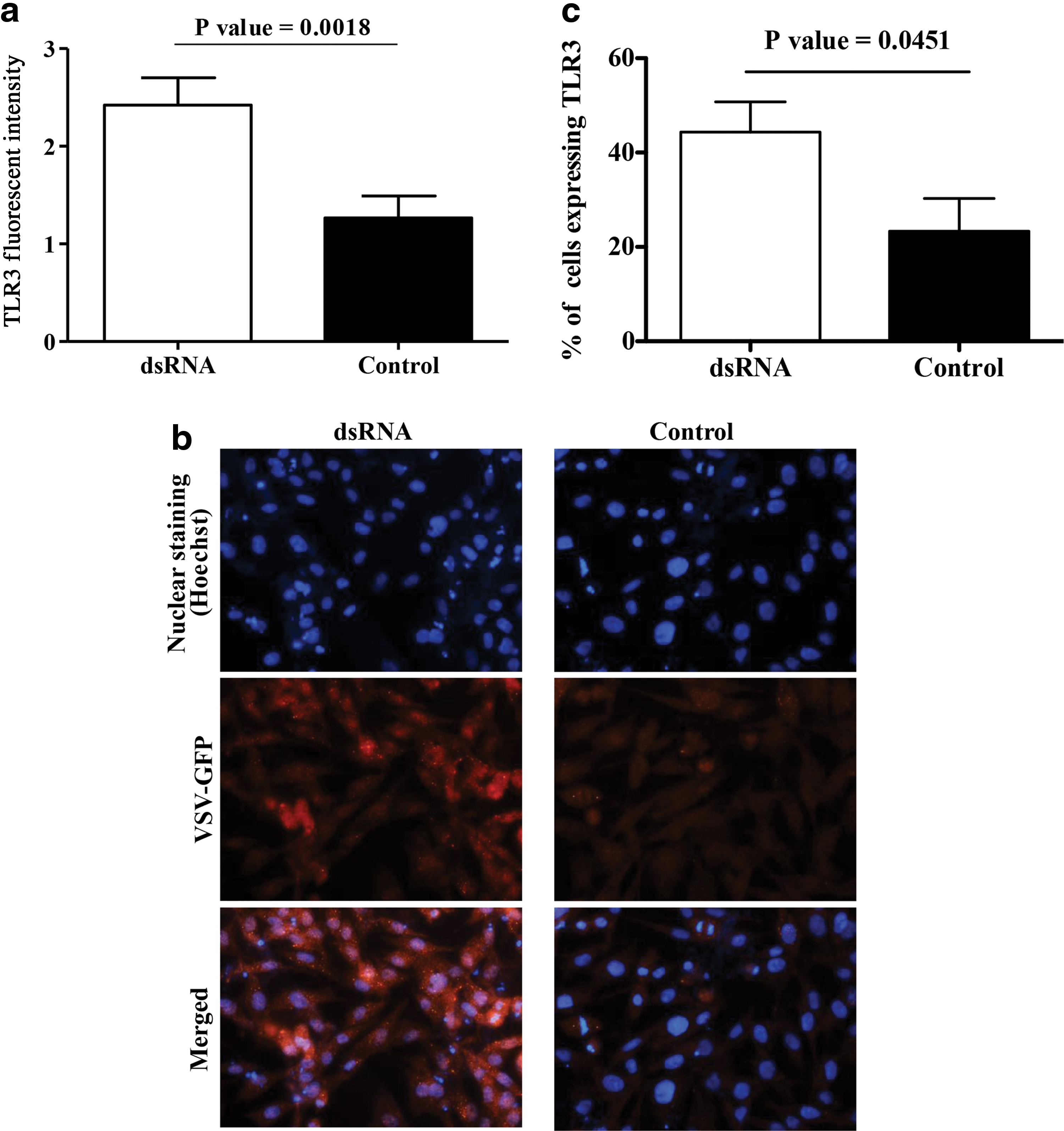
dsRNA increases TLR3 expression in avian fibroblasts. Avian fibroblasts (DF-1) were cultured in six-well plates at 1 × 106 cell/well for 24 h, treated with dsRNA at 50 μg/mL (n = 3) or Opti-MEM medium (n = 3) for 24 h, and the cells were fixed and immunofluorescent assay was done to determine the expression of TLR3 using primary rabbit anti-chicken TLR3 polyclonal antibody. The treated and control plates were imaged and analyzed using ImageJ software. The quantitative fluorescent intensity data
TLR3 ligand, dsRNA increases expressions of IFN-α and IFN-β
To determine the type 1 IFN molecules involved in the dsRNA-mediated type 1 IFN activity in avian fibroblasts, first, we performed immunostaining for IFN-β following dsRNA treatment of avian fibroblasts and found that dsRNA induces expression of IFN-β as assessed by fluorescent intensity (Fig. 3a, b, p = 0.0303) and % cells expressing IFN-β (Fig. 3c, p = 0.0012). We also found that dsRNA treatment increases expression of IFN-α as assessed by fluorescent intensity (Fig. 3e, f, p = 0.0117) and % cells expressing IFN-α (Fig. 3g, p = 0.0286).
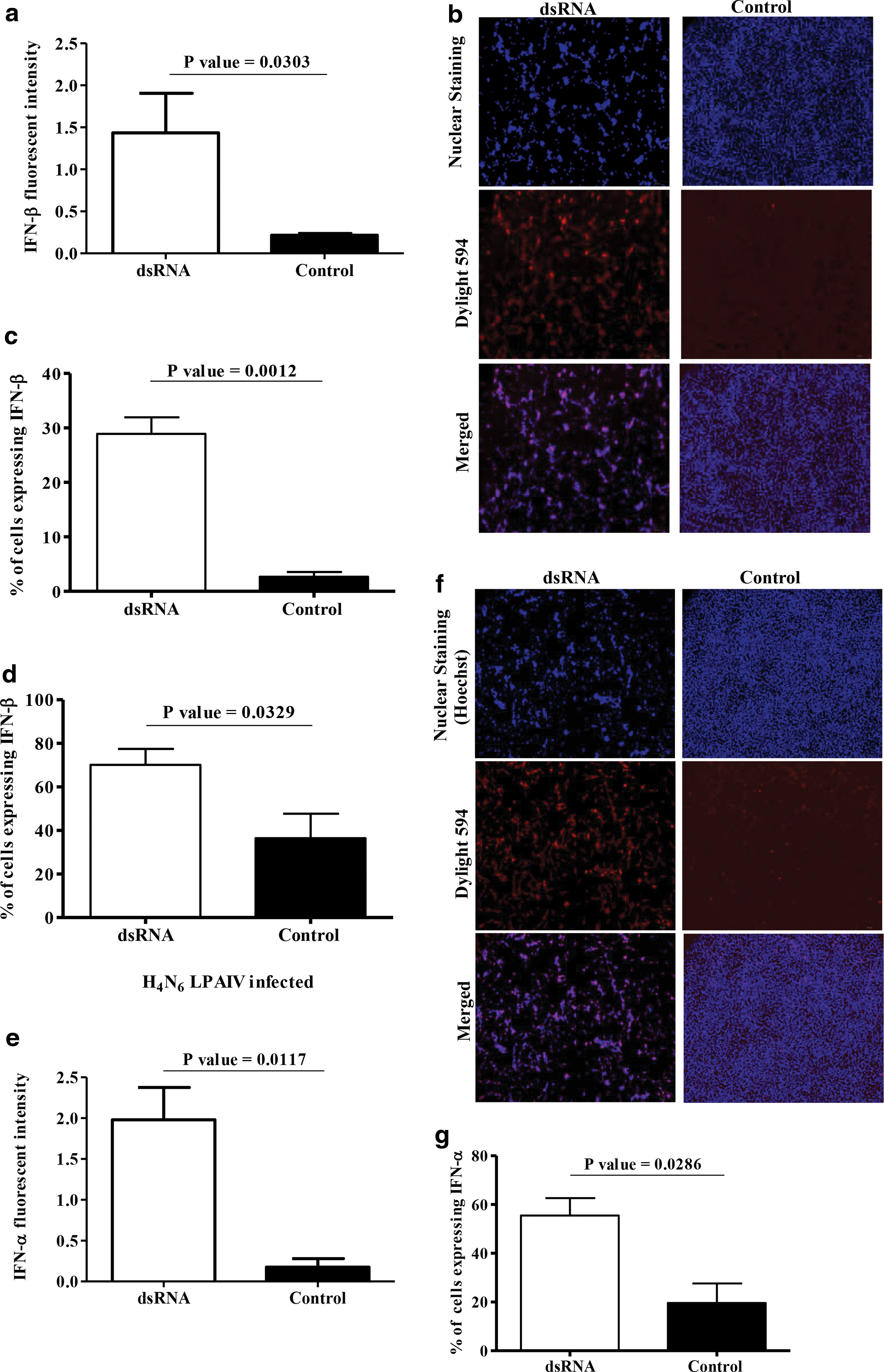
dsRNA increases IFN-α and IFN-β expressions in avian fibroblasts and IFN-β expression is enhanced during dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts. Avian fibroblasts (DF-1 cells) were cultured in six-well plates at 1 × 106 cells/well for 24 h and treated with dsRNA at 50 μg/mL (n = 3) or Opti-MEM medium (n = 3). The protein transport inhibitor was added to both the groups at 2 μL/mL after 6 h of incubation and incubated for further 18 h. The cells were fixed and immunofluorescent assay was performed for determination of expressions of IFN-β and IFN-α. Images were acquired by a fluorescent microscope and analyzed by ImageJ software. The quantitative fluorescent intensity data for IFN-β
IFN-β expression is enhanced during dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts
Since we observed that dsRNA treatment increases IFN-β expression, we then evaluated the expression of IFN-β in fibroblasts treated with dsRNA and infected with H4N6 LPAIV. We found that IFN-β expression in fibroblasts that were treated with dsRNA and infected with H4N6 LPAIV was significantly higher when compared to the group that was untreated and infected (Fig. 3d, p = 0.0329). The % of cells expressing IFN-β in dsRNA-treated uninfected cells was much lower when compared to that observed in dsRNA-treated and H4N6 LPAIV-infected fibroblasts (Fig. 3c, d).
TLR3 ligand, dsRNA-induced antiviral response against H4N6 LPAIV infection in avian fibroblasts is not attributable to type 1 IFN activity
To determine whether dsRNA-mediated antiviral response is indeed attributable to type 1 IFN activity, we used BX 795 as an inhibitor of TBK1, hence type 1 IFN production. As illustrated in Figure 4a, when BX 795 was used to inhibit the type 1 IFN production in the dsRNA-treated and control fibroblasts, VSV-GFP expression increased when compared to the cells that were treated only with dsRNA (p < 0.0001) and growth media.
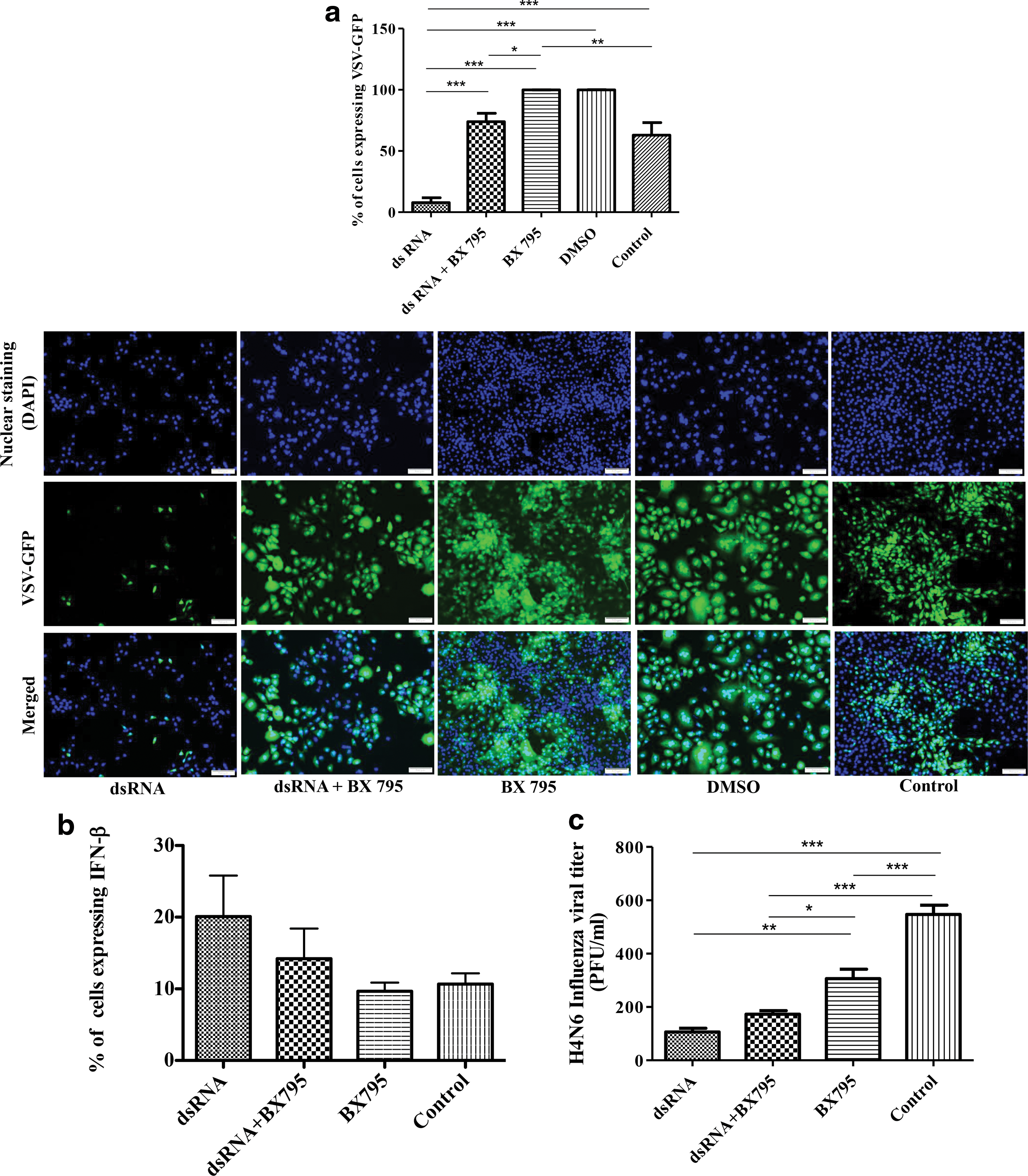
TLR3 ligand, dsRNA-induced antiviral response against H4N6 LPAIV infection in avian fibroblasts is not attributable to type 1 IFNs. First, to determine whether TBK1 inhibitor is able to inhibit type 1 IFN activity downstream of dsRNA-TLR3 signaling, DF-1 cells were cultured in 12-well plates at 0.2 × 106 cells/well for 24 h and treated with TBK1 inhibitor (BX 795) at 4 μM for 2 h, followed by treatment of dsRNA at 50 μg/mL (n = 3). The other groups were BX 795 alone (n = 3), dsRNA alone (n = 3), growth medium as a control (n = 3), or DMSO (n = 3). Following 24 h of incubation, the cells were infected with VSV-GFP (MOI = 0.1), and incubated the cells for further 24 h. These type 1 IFN bioassay plates were scanned using the In Cell Analyzer 2000 machine and analyzed by In Cell Analyzer 1000 software. The quantitative data along with representative images for each group are given
In other words, when type 1 IFN production was inhibited by BX 795, anti-VSV-GFP response was abrogated (Fig. 4a) indicating that anti-VSV-GFP responses induced by dsRNA and controls in avian fibroblasts were attributable to type 1 IFNs. The fibroblasts that were treated only with dsRNA showed the lowest VSV-GFP expression when compared to all the dsRNA-untreated cells (Fig. 4a, p < 0.0001). When we used TBK1 inhibitor, we did not observe significant blocking of the IFN-β production (Fig. 4b, p > 0.05), although the anti-VSV-GFP response was abrogated following treatment of dsRNA-treated fibroblasts, and control fibroblasts were treated with TBK1 inhibitor.
To determine whether dsRNA-mediated antiviral response against H4N6 LPAIV is attributable to type 1 IFN production, we inhibited the type 1 IFN production using TBK1 inhibitor and found that antiviral response against H4N6 LPAIV is not attributable to type 1 IFN production (Fig. 4c). Specifically, we did not observe significant abrogation of antiviral response against H4N6 LPAIV when dsRNA-mediated and basal level of type 1 IFN productions were blocked using TBK1 inhibitor (Fig. 4c, p > 0.05).
TLR3 ligand, dsRNA induces IL-1β expression, yet dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts is not attributable to IL-1β expression.
Since dsRNA-mediated antiviral response is not attributable to type 1 IFNs, we evaluated whether proinflammatory cytokines such as IL-1β are produced following dsRNA or control treatments of avian fibroblasts. We observed that dsRNA treatment of avian fibroblasts leads to significantly higher expression of IL-1β in avian fibroblasts as assessed by fluorescent intensity (Fig. 5a and b; [p = 0.0033]). The % of cells expressing IL-1β was also higher in dsRNA-treated fibroblasts when compared to the controls (Fig. 5c, p = 0.0447).
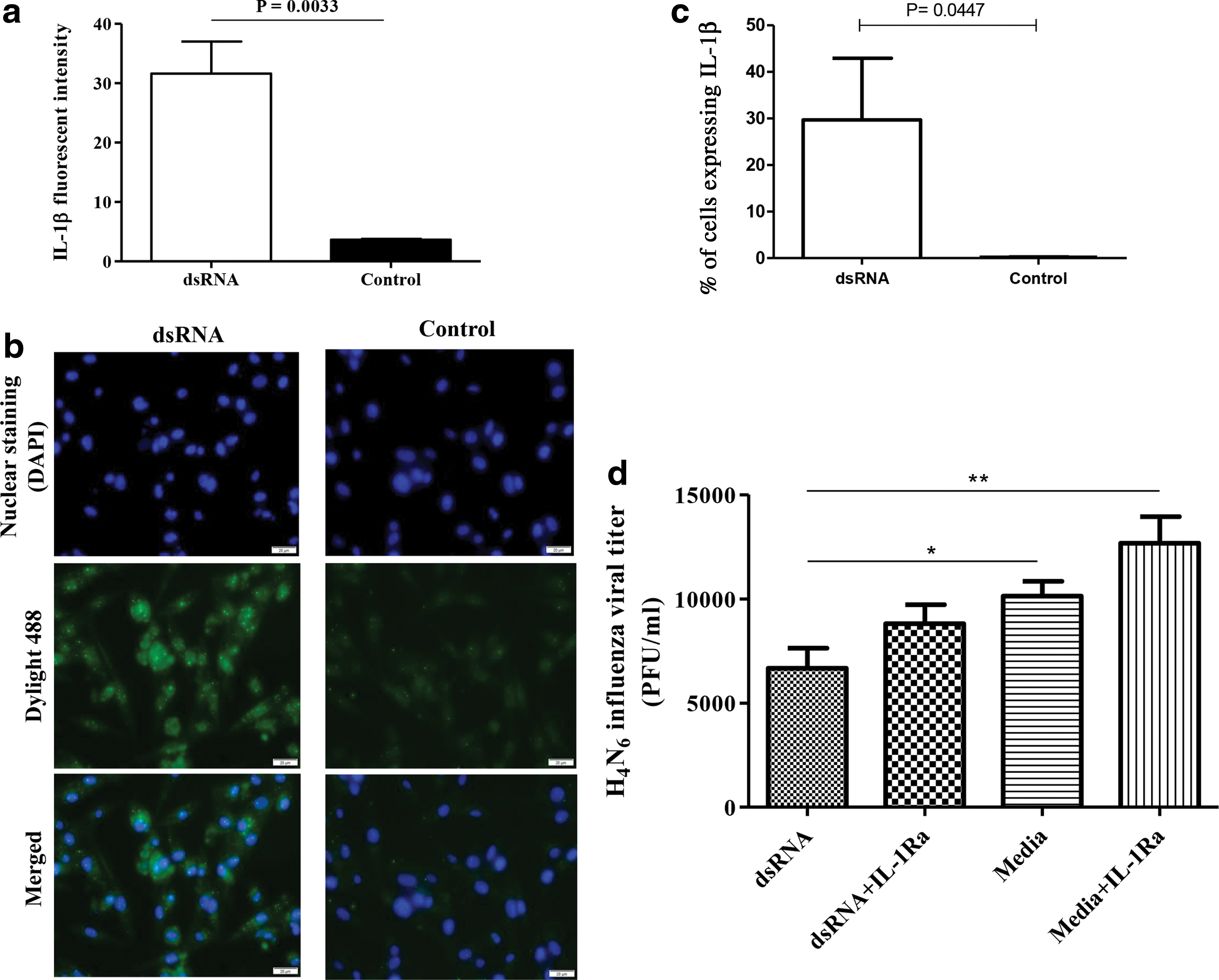
TLR3 ligand, dsRNA induces IL-1β expression, yet dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts is not attributable to IL-1β expression. The DF-1 cells were cultured on coverslips in 12-well plates with 0.2 × 106 cells/well and protein transport inhibitor (2 μL/mL) was added to culture medium after 6 h. After 24 h of culture, the cells were stimulated with dsRNA at a concentration of 50 μg/mL in Opti-MEM I reduced serum medium or only with the growth medium as a control (nine replicates per treatment). Following 24 h of treatment, immunofluorescent staining was performed after fixation with 4% paraformaldehyde (three wells per treatment). In immunofluorescent staining, unlabeled rabbit polyclonal antibody specific for chicken IL-1β was used as a primary antibody in 1:10 dilutions and biotinylated goat anti-rabbit IgG(H+L) was used as the secondary antibody in 1:400 dilution.
Since we observed that avian fibroblasts produce IL-1β in response to dsRNA treatment, we then investigated to see whether stimulation of avian fibroblasts with dsRNA inhibits H4N6 LPAIV replication in an IL-1β-dependent manner. In this study, we found that culture supernatants derived from avian fibroblasts following stimulation with dsRNA were able to inhibit H4N6 LPAIV replication compared to the medium controls (Fig. 5d, p < 0.05) and blocking IL-1β signaling using IL-1Ra did not abrogate the antiviral response elicited against H4N6 LPAIV (Fig. 5d, p > 0.05).
Discussion
The work described in the article uncovers a number of mechanisms relevant to dsRNA-mediated antiviral response elicited against LPAIV replication in avian species. First, we found that dsRNA could elicit antiviral response, reducing LPAIV replication in avian fibroblasts. Our observation of dsRNA-mediated antiviral response is in agreement with findings of the previous studies where dsRNA has been used to inhibit MDV replication in CEFs (37). Parenteral administration of dsRNA has also been shown to reduce LPAIV shedding in chickens (76).
Second, we showed that dsRNA-mediated antiviral response correlates with the expression of TLR3 in fibroblasts. Although this is the first study that showed that dsRNA-mediated antiviral response is correlating with TLR3 signaling, the study by Karpala and his colleagues (42) recorded, using RNA interference targeting TLR3 in chicken fibroblasts, dsRNA-mediated mRNA expression of IFN-α is dependent on TLR3 mRNA expression.
Third, we observed that the dsRNA-mediated antiviral response against H4N6 LPAIV is not attributable to the expression of type 1 IFNs, although IFN-β expression was higher in dsRNA-treated fibroblasts during H4N6 LPAIV infection. Although there are no comparable data in avian species that indicate dsRNA-mediated antiviral response is attributable to type 1 IFNs, some studies have shown that dsRNA is capable of increasing mRNA expression of type 1 IFNs in various avian cells (22,42,76,80). Finally, we explored the potential involvement of proinflammatory cytokines such as IL-1β in the dsRNA-mediated antiviral response against H4N6 LPAIV infection and found that, although dsRNA is capable of inducing IL-1β, the antiviral response mediated by dsRNA against H4N6 LPAIV infection is not dependent on IL-1β alone.
The ligand, dsRNA, has been shown to signal through TLR3 (3), RIG I, or MDA5 (44). In chickens, RIG I gene is absent (7) and, thus, dsRNA-mediated signaling in chickens is relying on either TLR3 (42) or MDA5 (43) binding. Although we did not investigate whether dsRNA was signaling through TLR3 or MDA5, our observation and the data of others indicate that dsRNA would have recognized by TLR3.
First, in this study, we observed that dsRNA increased the expression of TLR3 and due to the lack of required reagents, we were unable to observe the expression of MDA5 in the context of dsRNA-mediated antiviral response. In agreement with our data, in mammalian cells, dsRNA treatment has been shown to modulate the expression of TLR3 (46,61,74). This is also in agreement with other avian studies that showed increased mRNA expression of TLR3 gene in response to dsRNA treatment of CEFs (32,41).
Second, it has been observed that MDA5 preferentially acts as a receptor for short dsRNA strands (33) and, in our studies, we used long dsRNA strands as ligands for the induction of TLR3 signaling. Third, it has been shown that chicken MDA5 expression is not a critical factor that influences the antiviral response against AIV infection (33,43).
The results of in vivo studies in mammalian hosts suggest that activation of TLR3 signaling pathway by its ligand, dsRNA, provides protection against a number of viral infections. TLR3 ligand, dsRNA-mediated antiviral response against herpes simplex virus (HSV) has been shown in a mouse model (6). Furthermore, intranasal delivery of dsRNA to mice has also been shown to provide protection against lethal challenge with an HPAIV strain, H5N1, and against lethal seasonal influenza A/PR/8/34 [H1N1] and A/Aichi/2 [H3N2] viral strains (84). In vitro, in mammals, TLR3 has been shown to elicit antiviral response against rhinovirus infection (34) as well as against HSV infection (59).
In chickens, the mRNA expression of TLR3 has been shown to be associated with H5N1 AIV infection in brain (42) and intramuscular delivery of dsRNA has been shown to decrease shedding of LPAIV (76). In agreement with these observations, in a different context, our data show that dsRNA is capable of reducing LPAIV replication in avian fibroblasts. We could not find any literature to compare our data on dsRNA-mediated antiviral response against LPAIV replication in avian fibroblasts.
However, it has been also shown that dsRNA does not reduce LPAI infection in avian macrophages (8). The discrepancy of our results with that recorded by the later study may be due to the difference in cells that were stimulated with dsRNA: avian fibroblasts versus avian macrophages. Previously, it has been shown that, depending on the length, dsRNA elicits stronger response in fibroblasts when compared to macrophages (56). The dsRNA we used in our experiments as well as by Barjesteh and colleagues (8) were 1.5 kb to 8 kbp in size and considered long-strand dsRNA. The long dsRNA stretches are poorly taken up by macrophages when compared to fibroblasts as such antiviral responses elicited by macrophages in response to long dsRNA strands are insignificant (56).
We observed that activation of TLR3 signaling leads to IFN-α and IFN-β production in avian fibroblast. In mammals, it is well known that TLR3 signaling leads to the production of IFN-α and IFN-β (36). Although there is no comparable work performed in chickens, which demonstrated TLR3 signaling leads to IFN-α and IFN-β production, transcription of IFN-β gene following dsRNA treatment of avian fibroblast cells has been shown (42,43).
It was also evident that dsRNA treatment has improved the recognition of H4N6 LPAIV by TLR3, leading to higher IFN-β production when the dsRNA-treated cells were infected the H4N6 LPAIV. It has been shown previously that influenza virus can be recognized by TLR3 in human primary epithelial cells (48). When we used TBK1 inhibitor to block type 1 IFNs, type 1 IFN bioassay (VSV-GFP assay) indicated that type 1 IFNs were significantly reduced both in the dsRNA-treated and control fibroblast cells, although it was not reflected in the inhibition of IFN-β production. We understand the limitation of not measuring IFN-α in our type 1 IFN blocking experiments. Previously, it has been shown that IFN-α is ∼100-fold potent when compared to IFN-β against AIV infection in avian fibroblasts (64).
We observed that antiviral response against H4N6 LPAIV is not attributable to type 1 IFNs, although type 1 IFNs are effective in reducing avian viruses such as NCDV, infectious bursal disease virus, infectious bronchitis virus, MDV, and AIV subtypes (H9N2, H1N1, and H5N9) in vivo and in vitro (29,38,39,55,57,62,64). There can be few potential explanations, why we did not see that antiviral response against H4N6 LPAIV was not dependent on type 1 IFNs, although type 1 IFNs, particularly IFN-β, were produced downstream of dsRNA activation and infection of fibroblasts. First, it is possible that blocking of type 1 IFNs by TBK1 inhibitor in our experiment may not have been successful since we did not observe significant reduction in IFN-β expression following use of TBK1 inhibitor, although type 1 IFN assay indicated the reduction in type 1 IFN activity by TBK1 inhibitor.
It is also possible that some other mediators such as nitric oxide (NO) may have been involved in the dsRNA-mediated antiviral response against H4N6 LPAIV in avian fibroblasts since it has been shown that, in mouse fibroblasts, dsRNA elicits antiviral response against HSV attributable to NO (54). We evaluated two additional potential proinflammatory mediators, including NO, and found dsRNA does not induce NO production in avian fibroblasts (data not shown), although IL-1β expression was significantly expressed in fibroblasts following dsRNA treatment. However, blocking of IL-1β receptor engagement did not abrogate the dsRNA-mediated antiviral response against H4N6 LPAIV infection in fibroblasts.
We conducted blocking experiments targeting type 1 IFNs and IL-1β to investigate the links between dsRNA-mediated activation of these cytokines and antiviral response against H4N6 LPAIV infection in fibroblasts. When we blocked the functions of these cytokines, we did not observe significant abrogation of the dsRNA-mediated antiviral response against H4N6 LPAIV infection in fibroblasts. One potential explanation for observing a lack of antiviral response abrogation in our experiments may be that the functions of most of the cytokines are redundant and it is possible that compensatory mechanisms may be activated maintaining the antiviral response (66).
Second, it is also possible that, we did not target the most relevant cytokine with antiviral activity that will be induced following dsRNA treatment for avian fibroblasts and this may require further studies with a view of exploring other potential mediators that will be activated by dsRNA in avian fibroblasts. Third, it is possible that combination of type 1 IFNs and IL-1β, rather than individual cytokines, elicits antiviral response against H4N6 LPAIV infection in avian fibroblasts. Previously, it has been shown that combination of type 1 IFNs with other cytokines such as IL-29 or IFN-γ result in augmented antiviral responses against hepatitis C virus and HSV infections (60,68). Whether the dsRNA-mediated antiviral response against H4N6 LPAIV infection in avian fibroblasts is dependent on the combination of type 1 IFNs and IL-1β requires further studies.
We are puzzled by the fact that the use of TBK1 inhibitor in type 1 IFN assay resulted in significantly higher abrogation of antiviral activity against VSV-GFP (Fig. 5a) and significantly lower abrogation of antiviral activity against H4N6 LPAIV (Fig. 5c) in untreated controls. The sensitivity of VSV and AIV to type 1 IFNs in avian cells is not different (73). However, influenza viral nonstructural protein 1 (NS1) can counteract type 1 IFNs (31). In an AIV population or a viral stock, 40–60% of individual viral particles can express higher NS1 due to packaging of multiple copies of segment 8 of the genome (73) and, consequently, these viruses with higher NS1 expression can be resistant to type 1 IFN activity. We believe that this feature of AIV may potentially have influenced the type 1 IFN-mediated antiviral response against H4N6 LPAIV when compared to VSV infection of avian fibroblasts.
Assessing the control groups that are infected with H4N6 LPAIV (Figs. 1a, 4c and 5d), it is apparent that there is a variability in H4N6 LPAIV titers, although experimental conditions were similar. The MDCK cell line used for titration of H4N6 LPAIV is known to consist of heterogeneous cell populations and as a result, plaque assay may yield different influenza viral titers depending on the infected cell type within the MDCK cell line (53), and we believe this may be a potential explanation for the variation in H4N6 LPAIV titers between experiments.
LPAIV infection in chickens impacts not only poultry industry but also other animal species, including other livestock species and public health, since poultry can be a major source of virus for transmission to these animal species. Due to the limitations in avian influenza control measures, novel control measures are necessary and that should be based on understanding of the mechanisms of host responses elicited against this virus. Although our findings of dsRNA-mediated antiviral response against LPAIV need further investigations, dsRNA may provide a basis for developing control measures against LPAIV infections in chickens.
In conclusion, we discovered that the TLR3 ligand, dsRNA, increases the expressions of TLR3, type I IFNs, and IL-1β in avian fibroblasts. However, the antiviral response elicited by dsRNA against H4N6 LPAIV is not attributable to the production of type 1 IFNs or IL-1β, and the potential role of other innate immune mediators or combination of these mediators in the antiviral response mediated by dsRNA needs to be elucidated.
Ethical Statement
The Health Science Animal Care Committee (HSACC) approved the use of specific pathogen-free (SPF) eggs and embryos used in all our experimental procedures.
Footnotes
Acknowledgments
This study was funded by the Canadian Poultry Research Council, Alberta Livestock and Meat Agency, and Agriculture and Agri-Food Canada. H.A.-H. received a scholarship from the Egyptian government to visit the University of Calgary and participate in the research.
Author Disclosure Statement
No competing financial interests exist.