
Abstract
Multiple factors are associated with human papillomavirus (HPV) infection related cervical anomalies and its progression to cervical carcinoma (CaCx), but data vary with respect to the underlying HPV genotype and with population being studied. No data are available regarding the role of immunological imbalance in HPV infected CaCx pathogenesis from Northeast India, which has an ethnically distinct population, and was aimed to be addressed through this study. The study included 76 CaCx cases, 25 cervical intraepithelial neoplasia (CIN) cases, and 50 healthy female controls. HPV screening and genotyping were performed by PCR. Differential expression of tumor necrosis factor alpha (TNF-α) was studied at serum level by enzyme-linked immunosorbent assay and tissue level by immunohistochemistry and messenger RNA (mRNA) level by real-time PCR. The data were correlated with interferon gamma (IFN-γ) and NF-κβp65 levels at protein level, as well as HPV16 E6 and E7 expression at transcript level statistically. HPV infection and HPV16 genotype were predominant in the studied cohort. TNF-α was found to be downregulated at both mRNA and protein levels in CaCx cases compared to controls; and the gradient downregulation correlated with progression of the disease from normal→CIN→CaCx. TNF-α expression correlated with insufficient modulation of both IFN-γ and NF-κβp65. The HPV16 E6 and E7 transcripts were found to be sharply upregulated in CaCx cases strongly inversely correlated with the TNF-α expression. Significant role of TNF-α downregulation associated with insufficient IFN-γ and total NF-κβp65 modulation and the resulting significant upregulation of viral transcripts E6 and E7 are key to the HPV16 infection mediated CaCx pathogenesis in northeast Indian patients.
Introduction
C
Overall HPVs associated with CaCx have been found in 6–88% of females with CaCx in India (3,5,13,51). HPV type 16 and 18 are considered to be two highest risk varieties responsible for about 70% of all CaCx cases worldwide (8,9). Molecular and epidemiologic studies have solidified the association between high risk HPV types (especially HPV16 and HPV18) and cervical squamous cell carcinoma (4).
Although HPV infection is the primary etiologic agent of CaCx, it is not sufficient; and a variety of factors contribute to the CaCx development. As per the available literature suggestion, the most specific factors responsible for CaCx development other than HPV infection are host genetic and immunological factors (15). The HPV virus is common and sexually transmitted, and the infection is often transient and self-limiting, with a majority of infected women clearing the virus through effective immune response. Rarely infection may persist and progress to high grade lesions and cancer (30). The individuals with a suboptimal or improper immune response may be at increased risk of persistent HPV infection leading to sequel of various grades of dysplasia and/or associated malignancy. There are studies that have demonstrated that the immune system can recognize and eliminate aberrant cancer cells arising within the human body (7). Thus, the interaction between cancer and the immune system plays a pivotal role in cancer development.
Several cytokines that modulate the immunologic response have been implicated in the development of CaCx (1,11,20). HPV infection normally should elicit a Th1 response, which is important for the clearance of HPV infection, and lack of such a response may be associated with persistent infection or the development of HPV-related neoplasia (50). Tumor necrosis factor alpha (TNF-α) is a multifunctional cytokine playing a key role in apoptosis and cell survival, as well as in inflammation and immunity. Hence, loss of TNF-α could lead to immune-surveillance failure and lead to HPV infection related CaCx development. Indeed the role of TNF-α in orchestrating an antitumor immune response against HPV expressing CaCx cells has been evaluated, but results involving different study population belonging to different ethnicity and different pathological types are equivocal and inconclusive (2,55,58).
Interferon gamma (IFN-γ) produced by naive Th cells contributes to the antitumor activity and has both direct toxic effect on cancer cells and antiangiogenic activity (46). TNF-α production is known to be induced by IFN-γ through promoter-dependent transcription (60).Therefore the induction of IFN-γ and resulting TNF-α levels may possibly play a critical role in the susceptibility of cervical anomalies and progression to CaCx.
Second, NF-κβ is the central key pro-inflammatory molecule of which the role in immune modulation and carcinogenicity has been established, with data being equivocal; thereby NF-κβ has been rightly stated as a two edge sword (44). NF-κβ expression has been shown to be upregulated in majority of the literatures available till date (41,45). But recently it has been established by works using in vitro systems that TNF-α induction is dependent of NF-κβ activation and expression (63). Therefore the negative or positive side of the deregulation of NF-κβ expression needs critical evaluation in the pretext of a crucial TNF-α expression regulator in the pathogenesis of HPV linked cervical carcinogenesis, with special reference to northeast Indian cases where no data are available in this context.
High-risk HPV types can be distinguished from low-risk HPV types by the structure and function of the E6 and E7 viral oncoprotein products. The most important viral oncoprotein E6 products have the ability to subvert the cell growth regulatory pathways and modify the cellular environment to facilitate viral replication (49), enhance the integration of foreign DNA into the host cell genome (25), and have antiapoptotic effect (54). In addition, HPV E6 protein may function in immune evasion by modifying an infected cell's response to such apoptotic stimuli (21). Differential expression of another critical HPV viral oncoprotein E7 had been reported to have an important role in the pathogenesis of CaCx (64). With respect to E6 and TNF-α expression, available data are equivocal with some studies reporting that E6 sensitizes cells to apoptosis induced by TNF-α (38,59), while others reporting that it protects cells from TNF-α (17,19).
The data available on HPV related CaCx in Northeast India is majorly focused on the epidemiological genotypic variations of HPV in patient cohorts in Northeast Indian CaCx patients (13,23), but the existing data largely fall short of addressing the status and role of immunological imbalance with special reference to TNF-α in the pathogenesis of cervical anomalies, which has been tried to be addressed through this present study using an ethnically distinct northeast Indian population.
Materials and Methods
Patient enrollment and sample collection
A total of 76 clinically diagnosed CaCx patients (n = 76) who were admitted in the wards of Obstetrics and Gynecology department, Gauhati Medical College hospital, were enrolled in the study with patient's informed consent and all clinical details. Tissue biopsies from the cancerous region, as well as the noncancerous adjacent control area of the cervix, and whole blood (3 mL) were collected from all the study subjects. A portion of the collected tissue samples was fixed in 4% formalin embedded in paraffin and was used for routine histopathological examination and immunohistochemistry based analysis.
Retrospectively collected paraffin embedded biopsy cases (n = 25) from patients with precancerous lesions (cervical intraepithelial neoplasia [CIN] I–III) were also included for the present study for comparative analysis. Whole blood (3 mL) was also collected from age matched voluntary females as controls (n = 50); the plasma was separated and stored at −20°C for cytokine analysis. Patients with history of previous treatment for cervical precancerous lesions or cancer and without informed consent were excluded from the study. The study was approved by the Institutional Ethics Committee of Gauhati University, Guwahati, Assam, India.
HPV screening and genotyping by PCR
Total DNA was isolated from both affected and the adjacent nonaffected areas of cervix, by treating the samples with tissue lysis buffer and proteinase K digestion, followed by standard phenol–chloroform based extraction. Presence of HPV DNA was tested by PCR amplification using the established MY09/11 primers (16). As global data and regionally available data are indicative of the presence of either HPV16 or 18 in majority of the HPV infected CaCx cases, especially in squamous cell carcinoma, screening for HPV16 and 18 genotypes was done using specific primers. The primer sequences used for HPV16 were: F: 5′TCAAAAGCCACTGTGTCCTGA3′ and R: 5′GGTGTTCTTGATGATCTGCAA3′, whereas the primer sequences used for the HPV18 screening were: F: 5′CCGAGCACGACAGGAAC GACT3′ and R:5′TCGTTTTCTTCCTCTGAGTCGCTT3′. Total DNA was extracted also from the retrospectively collected paraffin embedded biopsy samples from patients with CIN of different grades using Qiagen kits following manufacturer's protocol and was then screened for the presence of HPV16 and 18 by PCR. CIN cases (n = 25) screened positive for HPV16 were only included for further analysis.
Differential TNF-α expression and CaCx pathogenesis
The serum TNF-α level in both the cases and the controls was analyzed by MAGPIX multiplex enzyme-linked immunosorbent assay (ELISA) method using customized magnetic bead based kit and xPONENT software based analysis (Millipore). As HPV16 is significantly associated with the pathogenesis of CaCx, the cases which showed positivity for HPV were only included for the analysis.
Total RNA from affected and nonaffected comparative areas was extracted by standard TRIzol method, quantified by NanoDrop spectrophotometer, and was converted to cDNA using the high capacity cDNA Reverse Transcription Kit (Applied Biosystems), random hexamer as primer and 2 μg of the quantified total RNA as template. Differential fold change in TNF-α messenger RNA (mRNA) expression between affected and non-neoplastic control areas was studied by real-time PCR using β-actin as internal normalization control.
Differential TNF-α protein expression in non-neoplastic control sections and affected CaCx sections were studied by immunohistochemistry using TNF-α specific antibody (Abcam) and super sensitive one-step polymer-HRP detection system (BioGenex). The slides were examined and graded for TNF-α expression by a senior pathologist.
Altered IFN-γ and NF-κβp65 modulation analysis and correlation with TNF-α expression and disease pathogenesis
The serum IFN-γ level in both the cases and the controls was analyzed by MAGPIX multiplex ELISA method using customized magnetic bead based kit and xPONENT software based analysis (Millipore). The protein level fold change of IFN-γ was further correlated with the serum levels of TNF-α estimated by multiplex ELISA method, by statistical analysis using SPSS 13.0 software. The tissue based differential protein expression of NF-κβp65 was analyzed by ELISA (ab176648; Abcam). Both IFN-γ and NF-κβp65 expression were correlated with TNF-α expression.
Differential HPV16 E6 and E7 oncoprotein mRNA expression analysis
The viral oncoprotein E6 and E7 has been established to be a crucial factor in the pathogenesis of CaCx, which it mediates by influencing the regulation of different growth and immune controlling mechanisms (64). The HPV16 E6 transcript expression was compared between the affected and nonaffected area by real-time PCR using SYBR green based chemistry, and the primers used were F: 5′TAAATGTGCTCGTCATTCTG3′ and R: 5′CCAATTACCCCTTT CAACA3′. The amplification was performed at 48°C annealing temperature, using β-actin as internal control. Differential mRNA expression of HPV16 E7 transcript was also studied by real-time PCR using the primers F: 5′GCAACCAGAGACAACTGA3′ and R: 5′AACCG AAGCGTAGAGTCAC3′, the annealing temperature being 56°C. The fold change in HPV16 E6 and E7 expression in the affected area was statistically correlated with the TNF-α mRNA expression by SPSS v13.0 software.
Results
Incidence of CaCx is highest during early middle phases of life
The study included a total of 76 CaCx cases of age ranging between 30 and 70 years with a median age of 46.5 years. Cases were categorized according to their age as 31–40, 41–50, 51–60, and 61–70 years, and most of the cases (39/76, 51.32%) were found to be under the age group 41–50 years indicating a higher incidence of CaCx during the early middle phases of life. The socioeconomic status of the patients is low in almost all the cases, reflecting not a proper practice of hygiene and cleanliness. The mean gravida of patients across all the age groups was 4.59 and that of patients in age group 41–50 years was 5.2.
HPV16 is highly prevalent in CaCx cases
HPV DNA was detectable in 63/76 (82.89%) cases both in their cancerous areas and in the non-neoplastic control areas, except two cases where HPV was detectable only in the affected areas. The HPV positive cases were subjected to genotyping for HPV16 and HPV18 and were categorized into three as follows: HPV16 positive, HPV18 positive, and non-16 non-18 HPV positive cases. HPV16 was the most predominant HPV genotype (Fig. 1a) in our studied cohort (58/63, 92.06%; p = 0.025) followed by HPV18 (Fig. 1b) only in 2/76 cases (3.17%) (Table 1).
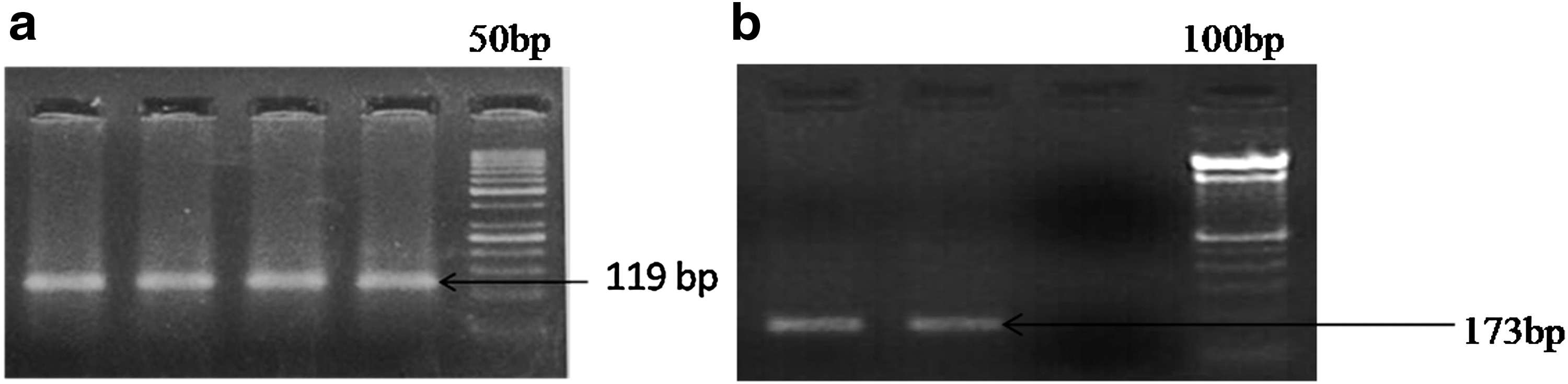
Agarose gel electrophoresis photograph depicting the
Cases represented as number (% age).
In one case HPV positivity was found only in the affected area.
HPV, human papillomavirus.
Serum TNF-α level is reduced in CaCx cases, and TNF-α expression is downregulated in cancerous regions
TNF-α level was measured only in the HPV16 positive cases, detectable with HPV infection in both the cancer affected and adjacent nonaffected areas (n = 57). When the serum TNF-α levels were compared between cases and healthy controls (n = 50), reduced serum TNF-α levels were observed in CaCx cases (5.535 ± 3.509 pg/mL) compared to controls (9.633 ± 3.607 pg/mL) (p = 0.162) (Fig. 2). The serum TNF-α levels were also found to be downregulated in HPV16 positive severe cases (with stage 3 or higher) compared to HPV16 positive CaCx cases of lower severity grade (p = 0.291).
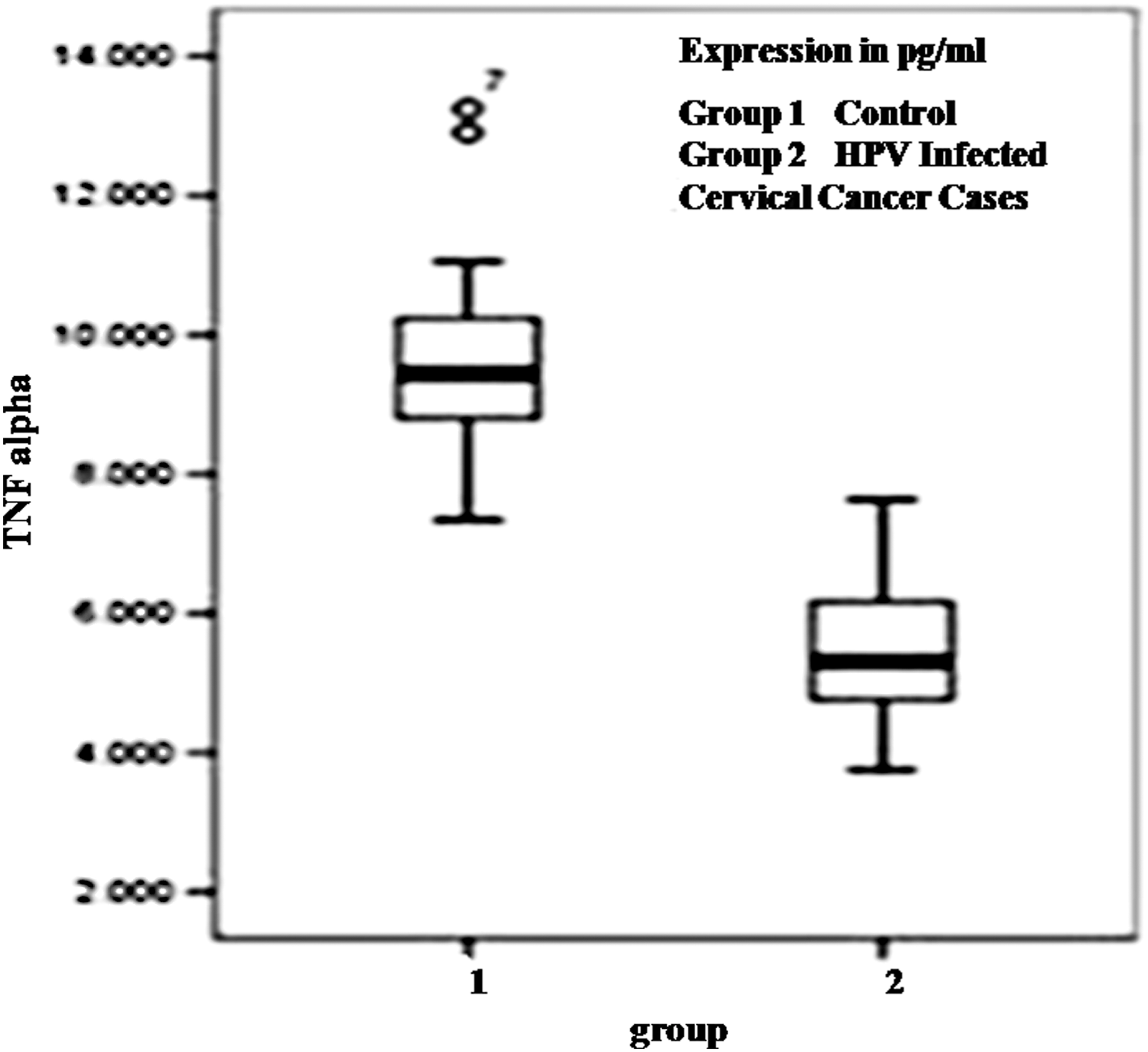
Box plot analysis for TNF-α in controls (group1) and HPV16 infected cervical cancer cases (group 2) showing significant downregulation of TNF-α is associated with the pathogenesis of cervical cancer. TNF-α, tumor necrosis factor alpha.
The real-time PCR based analysis for differential gene expression of TNF-α at mRNA level showed downregulation in TNF-α expression in affected area (0.28958 ± 0.273198 folds) compared to non-neoplastic adjacent control areas, in HPV16 positive cases. The differential mRNA expression of TNF-α was also found to be downregulated in HPV16 positive severe cases (with stage 3 or higher) (0.1622 ± 0.1234 folds) compared to HPV16 positive CaCx cases of lower severity grade (0.3556 ± 0.2112) (Fig. 3).
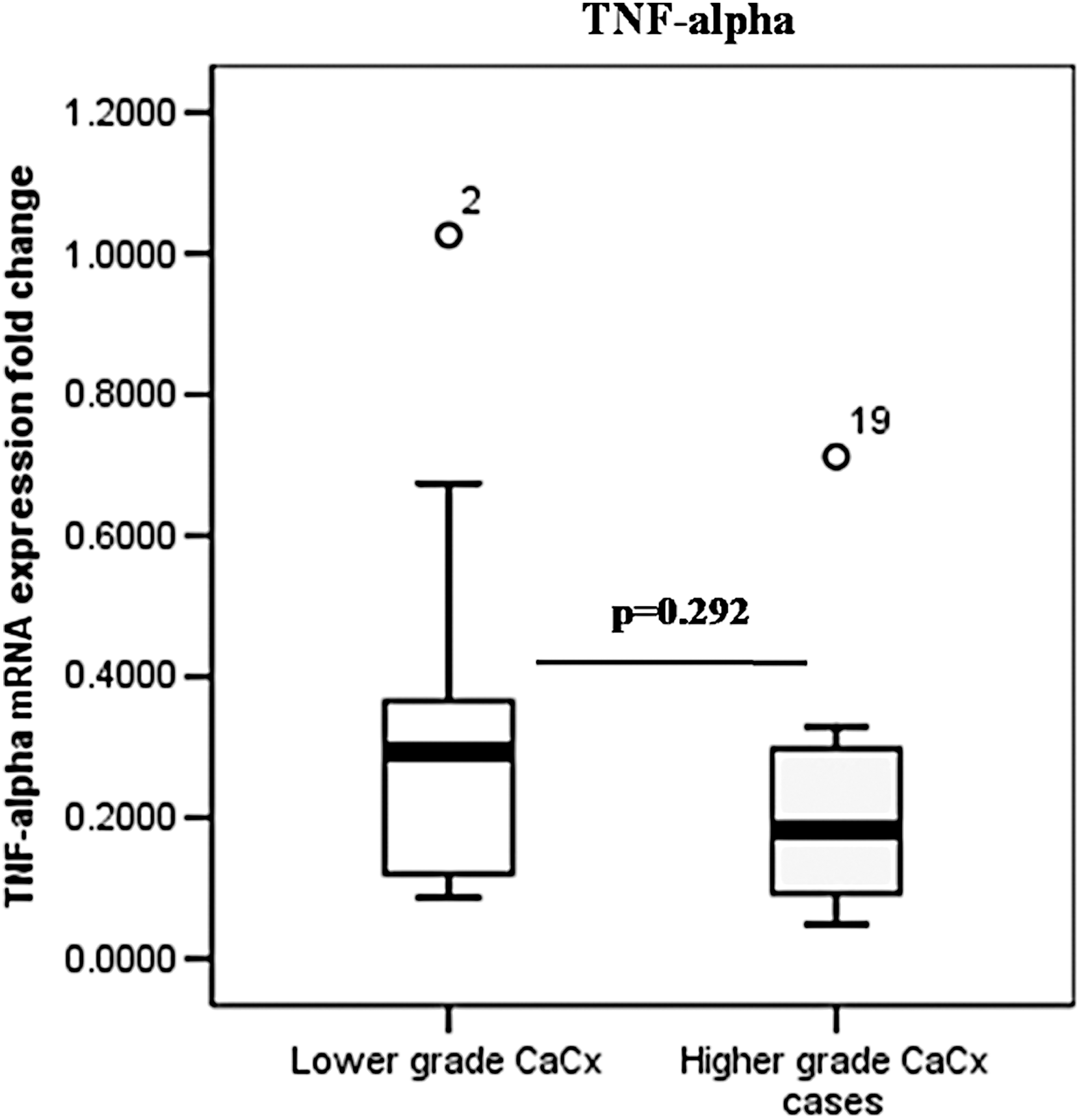
Box plot analysis for TNF-α expression in mRNA level in HPV16 infected cervical cancer cases showing downregulation of TNF-α in higher grade cervical cancer cases compared to lower grade cases. mRNA, messenger RNA.
TNF-α expression in protein level was analyzed by immunohistochemistry in randomly selected HPV16 positive cases (n = 35). Histological slides of both affected and adjacent nonaffected areas were analyzed, and it was observed that TNF-α expression was significantly lower in the affected areas compared to the non-neoplastic adjacent control regions in majority of the cases studied (26/35, 74%) (p = 0.036). Immunohistochemistry was carried out for the same parameter in CIN cases (n = 25) also. From the results obtained, it was found that the expression of TNF-α was decreasing gradually from CIN I to CIN III. Very little or no expression of TNF-α in affected areas of most of the cases was detectable indicating a negative correlation of TNF-α gene expression with the cervical carcinogenesis (Fig. 4).
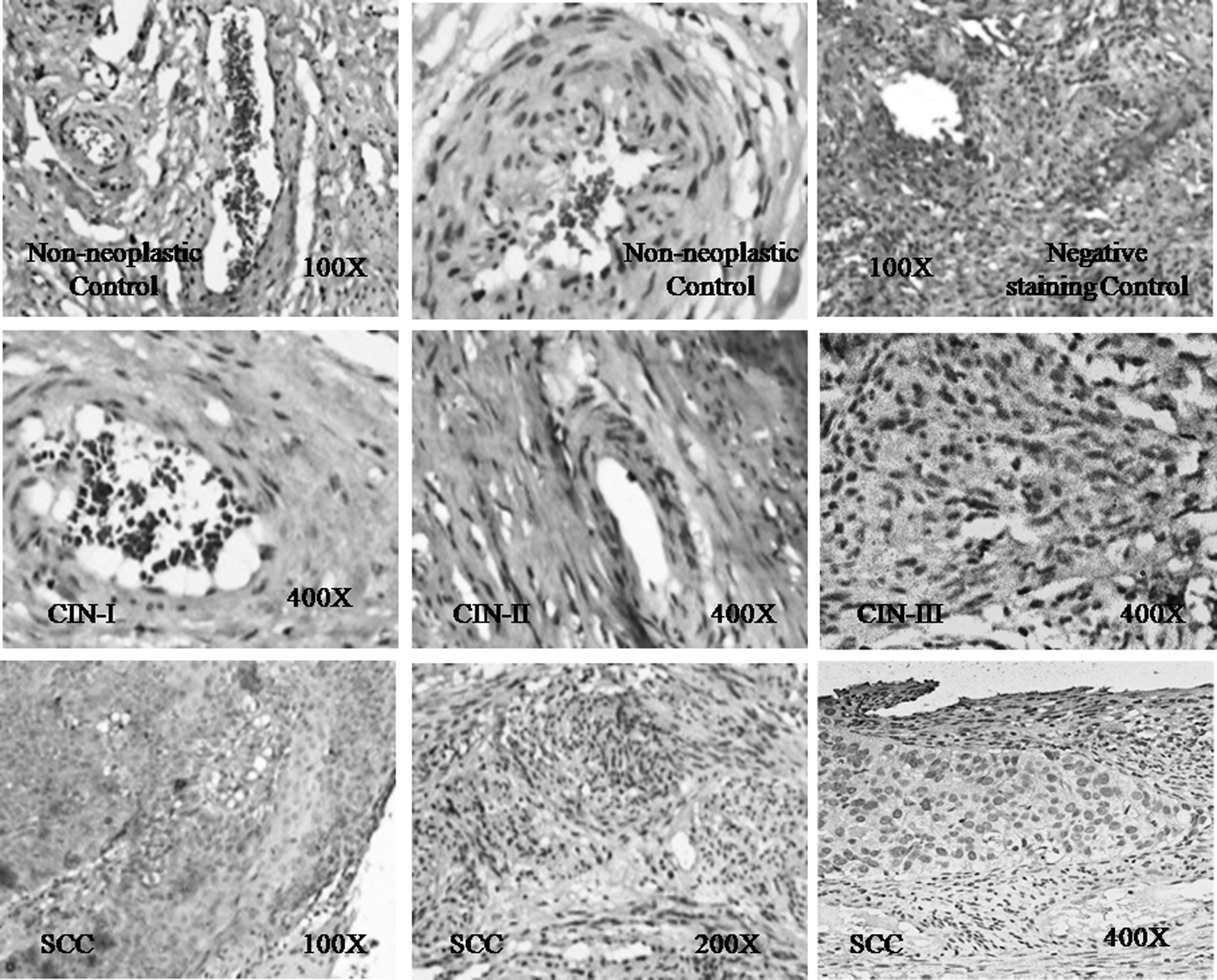
Representative panel of IHC results showing gradient downregulation of TNF-α expression during the progression to HPV related cervical cancer through the CIN stages, compared to the normal control cervix. IHC, immunohistochemistry; SCC, squamous cell carcinoma.
Serum IFN-γ levels are positively correlated with that of TNF-α which is also positively correlated with the expression of NF-κβp65
The serum IFN-γ levels evaluated by MAGPIX multiplex ELISA was found to be comparable between healthy control (8.184 ± 4.589) and HPV positive CaCx cases (9.259 ± 3.927, p = 0.734). However, a comparatively lower serum TNF-α level was observed in cases compared to controls. Statistical correlation analysis using SPSS v13.0 statistical software showed that the serum IFN-γ and TNF-α levels in HPV16 positive cancer cases correlated significantly (Table 2).
Cytokine expression levels represented as average ± standard deviation.
Correlation was significant at the 0.05 level (two-tailed).
IFN-γ, interferon gamma; TNF-α, tumor necrosis factor alpha.
The expression of NF-κβp65 at protein level was studied in tissues, and differential analysis was carried out in cancer affected and adjacent nonaffected control areas. A significantly lower expression was observed in affected area (2.585 ± 0.715 μg/mL), compared to the non-neoplastic adjacent control areas (4.065 ± 0.545 μg/mL), with a p-value, 0.002 (Fig. 5a). A significant downregulation was also observed in HPV16 positive severe cases (with stage 3 or higher) compared to HPV16 positive CaCx cases of lower severity grade (p = 0.030) (Fig. 5b). A comparative analysis of data obtained from NF-κβp65 expression in protein level in the affected areas and serum TNF-α levels in HPV16 positive cases revealed a positive correlation between the two (Pearson correlation = 0.568, p = 0.138; and Spearman's rho = 0.612, p = 0.116).
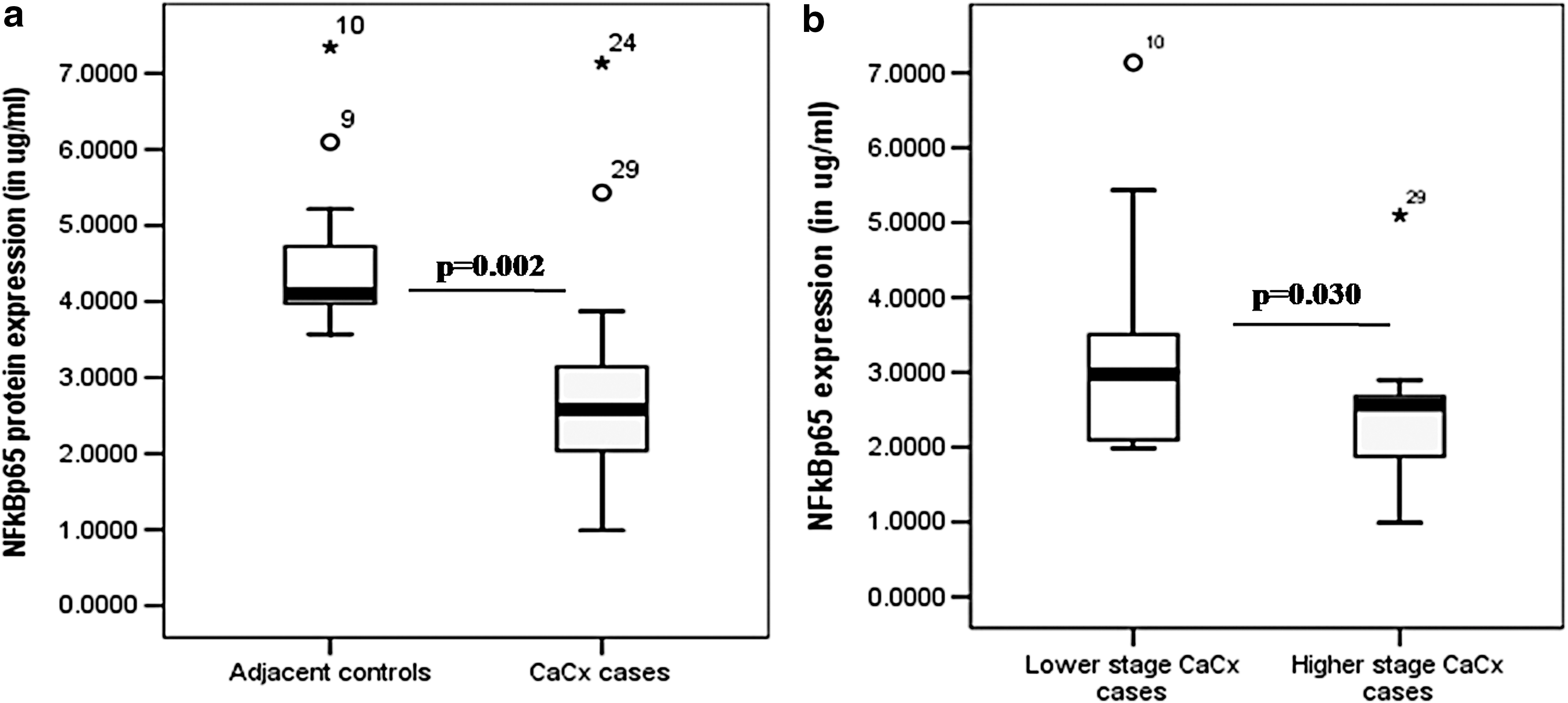
Box plot analysis for NF-κβ expression at protein level
HPV16 E6 and E7 gene expression in cancer affected regions and adjacent control regions is comparable
The oncogenes E6 and E7 of HPV16 were found to get expressed both in the affected and adjacent nonaffected control areas indicating the presence of viral oncoproteins in non-neoplastic areas of cervix adjacent to cancerous areas. The differential mRNA expression analysis of HPV16 E6 and E7 gene in cancer affected areas and the adjacent non-neoplastic areas of cervix done by real-time PCR showed that in 2 out of 57 cases, the E6 expression was comparable between the areas evaluated; however, the other cases showed that there is a sharp upregulation of E6 mRNA expression in affected area (186.9715 ± 157.3339 folds) compared to control areas. The differential mRNA expression of E6 was significantly upregulated in affected area of higher grade (197.6550 ± 164.4512 folds) CaCx cases in comparison to lower grade (98.7820 ± 72.6890 folds) CaCx cases compared with control areas (p = 0.032) (Fig. 6a).
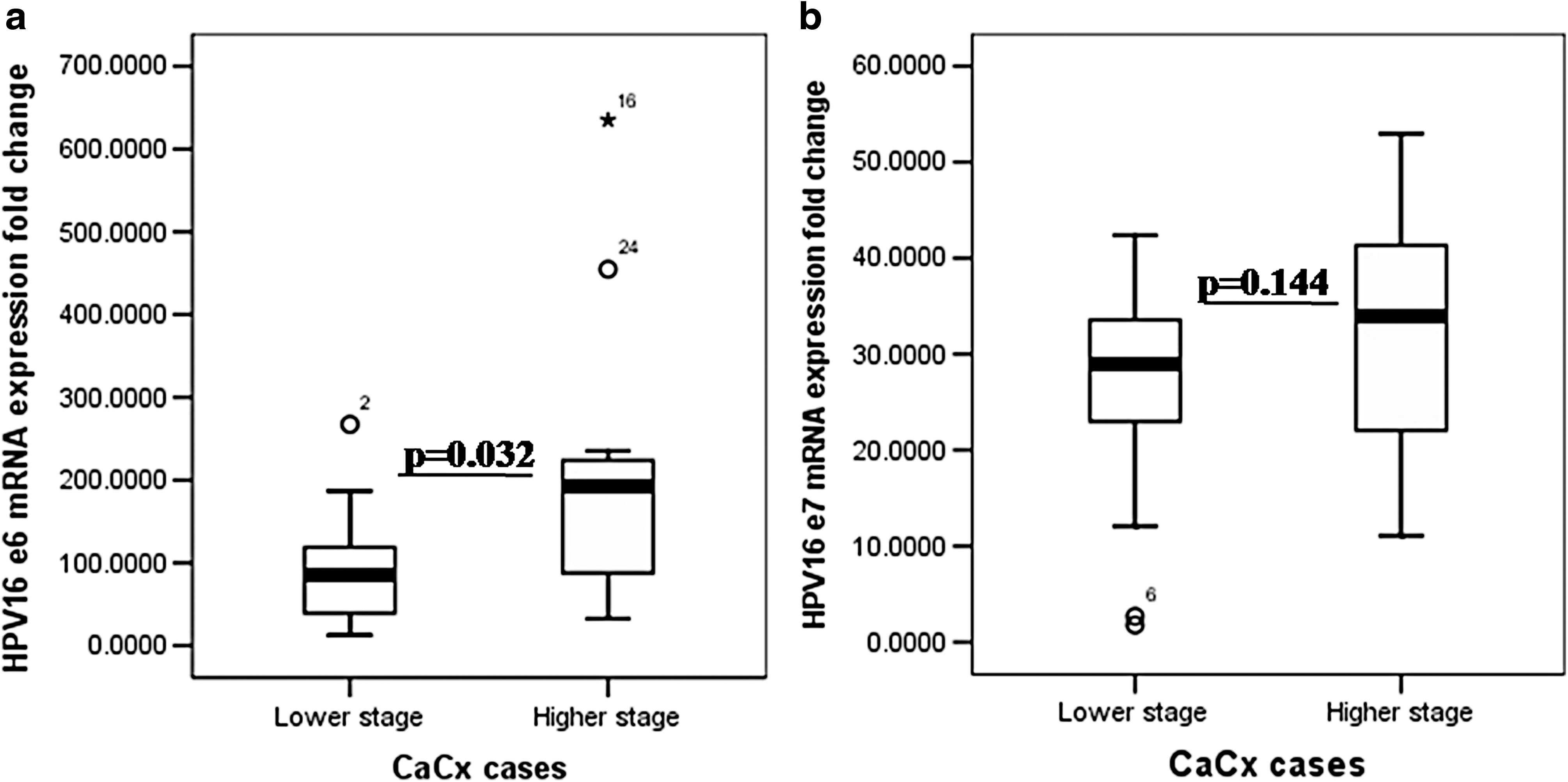
Box plot analysis for HPV16
There is a sharp upregulation of E7 mRNA expression in affected area (35.619 ± 23.107 folds) compared to control area in all the cases except three where the expression is comparable between the two areas. Furthermore, the differential mRNA expression of E7 was comparable between the higher grade CaCx cases and lower grade CaCx cases (p = 0.144) (Fig. 6b).
Expression of viral oncoproteins at mRNA level is inversely correlated with that of TNF-α
Correlation analysis showed that E6 expression is inversely correlated with TNF-α mRNA expression (Pearson correlation = −0.549, p = 0.126; and Spearman's rho = −0.603, p = 0.085) in all the cases both in cancerous and adjacent non-neoplastic areas. Similarly, a negative correlation was also found between E7 and TNF-α mRNA expression (Pearson correlation = −0.293, p = 0.524; and Spearman's rho = −0.643, p = 0.119).
Discussion
The present study involves the investigation regarding the association of HPV genotype, viral oncoproteins, and the host immunological factors with CaCx. The pathogenesis of HPV associated cervical anomalies and its progression to carcinoma has been both attributed to HPV genotype involved; viral factors as well to the immunomodulatory status of the individual patients resulting in either the susceptibility and severity to the HPV associated disease pathogenesis or viral clearance (15). There is heterogeneity in the existing data coming from different global pockets with difference in the predominance of HPV genotypes associated with CaCx pathogenesis, and henceforth, the available data are equivocal in nature.
Lacunae also exist in Northeast Indian context, which has an ethnically distinct and diverse tribal dominated population compared to other parts of India from where the majority of the available data on CaCx are about the HPV genotype diversity and not the host related factors associated with the disease pathogenesis. Present study is a conscious effort to underline the role of disturbed or altered immunological profile in the pathogenesis of CaCx with a predominant HPV16 infection. The majority of the patients are of the reproductive premenopausal age group, which indicates early HPV infection as evident from our data. As observed from the study, women under age group 41–50 are predisposed to contract CaCx the most and almost all the patients are found belonging to lower socioeconomic background indicating a poor practice of health and hygiene, a data supportive of the already existing data (47).
Immune status of an individual and its response corresponding to the pathogenesis of a disease like CaCx may vary according to the level of expression of certain genes, and therefore, identifying the immunological factors which critically influence HPV infection associated cervical anomalies in northeast India is urgently required to clinically address and devise treatment of the ever increasing load of cervical anomalies in this geographical part of the country. The present study shows the association of HPV in 82.89% of CaCx cases, which is almost concordant with the already existing data (22). Although there is a geographical variation in the genotype distribution of HPV worldwide, different studies have shown that high risk HPV16 and 18 play an associative role in most cases of CaCx (27).
In India, among all HPV types, HPV16 and HPV18 have been found to be prevalent in CaCx (4,22,51). Among the HPV positive cases in our studied cohort, HPV16 was the most prevalent genotype (92.06%) followed by HPV18 (3.17%), which is comparable with another study from Northeast population (13), but contrary to Ghosh et al., (23) who showed that HPV18 compared to HPV16 is the prevalent genotype in CaCx patients from southern Assam. Overall, HPV Genotype 16 has been shown as the most common type (59.4%) in a multicenter study in India (3). Similarly a report from Indonesia also confirms that the most prevalent genotype was HPV16 (90%) (56). It has also been reported that high-grade squamous intraepithelial lesion cytology is more frequent in women infected with HPV16 than in those infected with HPV18 and other types (18).
There is a strong basis that immune system can recognize and eliminate aberrant cancer cells arising within the human body (39,66). TNF-α, a multifunctional cytokine, is known to orchestrate cellular, inflammatory, and immunosurveillance role, and its role in HPV infection related CaCx pathogenesis has been defined but inconclusively (2,55,58). TNF-α levels were found to be lower in serum (p = 0.162) in HPV16 infected CaCx cases compared to controls. Similarly, downregulated mRNA expression of TNF-α gene (0.28-folds) in cancer affected regions of the cervix compared to the non-neoplastic adjacent control regions indicates a defensive role of this cytokine in cancer. This finding is also supportive of the downregulation of TNF-α at protein level in CaCx as observed in the present study. Tissue specific TNF-α protein expression was found to be inversely correlated with HPV16 infection associated disease severity, the lowest expression being observed in CaCx cases which underlines the prognostic and clinical significance of TNF-α.
Multiple pro-inflammatory and inflammatory cytokines influence TNF-α expression, IFN-γ and NF-κβ being the most important ones. Higher IFN-γ levels are required for antiviral responses. The absence of significant increase in IFN-γ levels in the HPV16 infected CaCx cases (p = 0.734, Table 2) is a direct indicator of insufficient antiviral response contributing to CaCx pathogenesis. The statistical analysis also showed that serum IFN-γ level in HPV infected CaCx case is significantly correlated with the serum TNF-α levels (Pearson correlation = 0.466, p = 0.011; and Spearman's rho = 0.369, p = 0.038). This finding is supportive of the fact that TNF-α production is induced by IFN-γ (60). IFN-γ expression influences TNF-α expression and vice versa. The results therefore indicate that the loss of TNF-α could be advantageous for tumor cells to escape immune clearance directly as well for the persistence of underlying HPV infection due to insufficient IFN-γ levels in the absence of sufficient TNF-α levels.
Constitutive activation of NF-κβ signaling is a key event in virus and nonvirus-induced carcinogenesis. NF-κβp65 regulates the expression of a large number of genes associated with inflammation (32,42), tissue damage and repair (36,65), cell differentiation (28,35), apoptosis (29), and tumor growth (31,52). In HPV16 positive cases in this study, the NF-κβ gene expression at protein level was significantly higher in non-neoplastic control areas compared to affected areas of the cervix (p = 0.002).
Correlation analysis between the NF-κβp65 expression in the affected area with the serum TNF-α levels in HPV16 positive cases showed a positive correlation (Pearson correlation = 0.568, p = 0.138; and Spearman's rho = 0.612, p = 0.116), suggesting a role of TNF-α and NF-κβ in regulating disease progression. These data are contrary to earlier results on the role of NF-κβ in CaCx (41,45,57), but also needs to be seen through the perspective that TNF-alpha can potentially induce both survival and apoptotic signals, and only the survival signal is mediated by the activation of NF-κβ which is a characteristic in cancer cells while the adjacent cells or tissues are subjective to apoptosis thus marginalizing and localizing tumor or abnormal cell growth as is evident from our immunohistochemistry results, which show higher expression of TNF-α in CIN cases with progressive downregulation with increasing severity.
NF-κβ activity has been reported to enhance tumor cell sensitivity to apoptosis and senescence. In addition, it has been shown that canonical NF-κβ is a Fas transcription activator, and the alternative NF-κβ is a Fas transcription repressor (37). TNF-mediated cell proliferation inhibition of primary human kerationocytes (PHKs) and HPV16-immortalized keratinocytes was earlier reported to be associated with NF-κβ activation (53). Second, inhibition of NF-κβ in human keratinocytes was reported to promote development of malignant human epidermal lesions resembling squamous cell carcinoma in mice (12), and a similar situation may also be existing in our cases since all the cases of CaCx in our studied cohort were squamous cell carcinoma. The differences in results may be also a tissue dependent context or may involve additional steps or factors that are not well understood. Moreover, the difference in the expression of different NF-κβ family members (NF-κβ1, NF-κβ2, RelA, RelB, and c-Rel) and their localization may shed more light into the matter which is a limitation of our study, but the length of the investigation required is also not in the purview of the present study.
It is also a possibility that NF-κβ expression in the adjacent control tissues is a mechanism of chronic inflammation in the neighboring cells linked to cancer progression, but this will also require a model based system to establish, which is again not in the purview of the present study. But most importantly, as TNF-α is conclusively illustrated to show an anti-tumorigenic role through our data, and since NF-κβ is to be a first responder to harmful cellular stimuli and had been reported to be induced by TNF-α, the positive correlation of NF-κβ expression may be a beneficial component.
Viral oncogenes E6 and E7 are found to be highly expressed in cancerous areas. These two HPV oncoproteins have been underlined to be ideal targets for development of a therapeutic HPV vaccine (40). It has been previously reported that TNF inhibits HPV16-immortalized but not HPV18-immortalized keratinocyte proliferation in HPV-transformed cell lines (58,61). Apart from the role of viral integration into the host genome (25) and exerting antiapoptotic effect (54,62), E6 has also been reported to be associated with the blockade of IFN (48) and to modify the cellular environment to facilitate viral replication (49).
Present investigation also shows that E6 expression has a sharp upregulation in the affected area (186.9715 ± 157.3339 folds) compared to control area, which indicates the importance of viral oncoprotein E6 in HPV related disease pathogenesis. TNF-α can downregulate HPV16 E6/E7 mRNA transcription in HPV16-immortalized human keratinocytes (34). The correlation based analysis between E6 and TNF-α mRNA expression showed that the upregulation in E6 expression is inversely correlated with TNF-α expression (Pearson correlation = −0.549, p = 0.126; and Spearman's rho = −0.603, p = 0.085), indicating the concurrence with the existing data and mechanism.
Viral oncogene E7 expression showed sharp upregulation in affected area (35.619 ± 23.107 folds) compared to control area. Acute expression of E7 oncogene from HPV16 has been shown to be sufficient to overcome TNF-α cytostatic effect on primary human keratinocytes (2,6). When the correlation based analysis was performed between E7 and TNF-α mRNA expression in all the cases statistically, it was found that the upregulation in E7 expression inversely correlated with TNF-α expression (Pearson correlation = −0.293, p = 0.524; and Spearman's rho = −0.643, p = 0.119). It has been previously demonstrated that E6/E7-transfected keratinocytes exhibit a shift of TNF receptor profile from type 1 to type 2 and weaken the apoptotic response to TNF-α stimuli (10). Existing evidence supports that E7 expression overcomes TNF effect through the upregulation of a group of genes associated with cell cycle progression (6). Higher expression of the viral oncogenes E6 and E7 in cancer affected areas and its correlation with host immunological factor TNF-α are suggestive of the immune-regulatory role of HPV factors in cancer pathogenesis. Moreover, the fact that TNF-α expression was found to be higher in control areas, even if there was expression of viral oncogenes E6/E7, is suggestive of the direct influence of TNF-α rather than the viral oncoproteins in the pathogenesis of the disease.
The data highlight the significant role of TNF-α downregulation, which is associated with insufficient IFN-γ level and total NF-κβp65 modulation, and the resulting significant upregulation of viral transcripts E6 and E7 is key to the HPV16 infection mediated CaCx pathogenesis in this studied cohort. Our data also underline the prognostic significance of both TNF-α and HPV16 viral transcript expression, as well as provide valuable insights for possibilities of efficient TNF-α agonist based therapy in combination with existing chemotherapeutics for combating HPV16 associated cervical anomalies. Moreover, the study holds importance in context to northeast India where HPV vaccine is a truly uncommon practice especially in rural parts. Our future investigations will be directed toward the identification and characterization of genetic polymorphisms or mutations in genes like TNF-α and to correlate this with cancer pathogenesis and also to evaluate the role of other immunological component, which might have some role to play in this disease pathogenesis.
Footnotes
Acknowledgments
The authors acknowledge Department of Biotechnology (DBT), Govt. of India, for financially supporting the study. The authors also acknowledge Dr. Mukherjee, New Delhi, and Dr. Anupam Sarma and Rizwana Sultana, Guwahati, Assam, for the immunohistochemistry analysis.
Author Disclosure Statement
No competing financial interests exist.