
Abstract
Follicular helper T cells (TFH) and follicular regulatory T cells (TFR), subsets of T cells, co-regulate the reactions of germinal center (GC) B cells. TFH provide help to the GC response, while the TFR suppress those processes. Nevertheless, the role of circulating TFR (cTFR) and circulating TFH (cTFH) in chronic hepatitis B virus (HBV) infection remains limited. Twenty healthy controls (HCs), 24 patients with chronic hepatitis B (CHB), 23 with HBV-related liver cirrhosis (LC), and 18 with HBV-related hepatocellular carcinoma were enrolled in our study between October 2015 and September 2016. We detected the frequencies of cTFR-like cells and cTFH-like cells, the percentage of programmed cell death-1 (PD-1), inducible co-stimulator (ICOS), and interleukin-21 (IL-21) expressed on circulating CD4+CXCR5+ T cells by flow cytometry. Compared to the HC group, the percentage of cTFR-like cells and ratio of cTFR-like/cTFH-like were significantly increased in patients with HBV infection. A raised percentage of PD-1 on circulating CD4+CXCR5+ cells and decreased IL-21-producing circulating CD4+CXCR5+ cells were observed in CHB and LC. The production of IL-21 by circulating CD4+CXCR5+ cells was significantly higher in HBeAg negative group than the positive one. Patients with high levels of alanine aminotransferase and HBV-DNA accompanied increased CXCR5+PD-1+CD4+ T cells. In addition, the frequency of cTFR-like cells and the ratio of cTFR-like/cTFH-like were positively correlated with FIB-4 and APRI. Increased cTFR-like cells and impairment of circulating CD4+CXCR5+ T cells might participate in HBV chronic infection and HBV-related diseases.
Introduction
H
T cells play a critical role in resistance to viral infection. CD8+ T cells directly eliminate virus-infection cells through cytolytic and noncytolytic mechanisms, while CD4+ T cells play a key regulatory role (28). Successful HBV clearance is due to vigorous, broad, and polyclonal T cell response. On one hand, this helps the clearance of virus; on the other hand, production of antibodies by B cells against antigens is also dependent on CD4+ T cells (25).
Follicular helper T cells (TFH) and follicular regulatory T cells (TFR) have been identified as novel subsets of CD4+ T cells located in follicles, first reported in 2001 and 2013, respectively. Both types of cells express chemokine (C-X-C motif) receptor 5 (CXCR5), which allows them to migrate into B cell follicles in response to CXCL13. The two cells have different phenotypes and functions. TFR and TFH both express programmed cell death-1 (PD-1) and inducible co-stimulator (ICOS), but expression of FoxP3 in TFR is distinct with TFH.
TFH interact with B cells by secreting the cytokines interleukin-4 (IL-4) and interleukin-21 (IL-21) and express surface molecules (CD40L, ICOS), which contribute to B cell proliferation, somatic hypermutation, and class-switch recombination. TFH-mediated B cell activation and antibody production are inhibited by TFR. Although the mechanism of TFR in downregulating germinal center (GC) response is still unclear, such cells dynamically regulating in vitro and in vivo reactions of the GC have been reported (7,8,32).
Peripheral blood CD4+CXCR5+ T cells are counterparts of TFH and were reportedly increased in patients undergoing HBsAg seroconversion or autoimmune hepatitis (22,29,35,39). In addition, researchers pointed out that the circulating CD4+CXCR5+ T cells are closely related to TFH of lymphoid organs; those cells might be helpful to assess the situation of TFH in HBV infected patient (20). Moreover, circulating CD4+CXCR5+ T cells could be divided into TFR-like cells (CD4+CXCR5+FOXP3+) and TFH-like cells (CD4+CXCR5+FOXP3−) according to the expression of FOXP3 or not. The imbalance ratio of TFH-like cells and TFR-like cells was found in myasthenia gravis patients (42). However, little is known about the roles of circulating TFR (cTFR) and circulating TFH (cTFH) in the HBV-related disease progression.
Our previous studies demonstrated that host genetic factors, virus genotypes, and molecules expressed on T cells were correlated with HBV clearance or clinical outcome (3,21,44). In this study, we analyzed the frequencies of cTFH-like T cells and cTFR-like T cells and the phenotype of circulating CD4+CXCR5+ T cells in HBV infected patients. The aim of our study was to determine whether circulating follicular T cells involved in the inadequate humoral immune response result in persistent HBV infection and disease progression.
Patients and Methods
Patients
Twenty healthy controls (HCs), 24 patients with CHB, 23 patients with HBV-associated liver cirrhosis (LC), and 18 patients with HBV-related hepatocellular carcinoma (HCC) were enrolled in our study between October 2015 and September 2016. Among 65 patients with persistent HBV infection, 29 patients were HBeAg positive. According to Asian-Pacific clinical practice guidelines on HBV and American Association for the Study of Liver Diseases guidelines (33,37), CHB is defined as HBsAg seropositive status at 6 months or beyond and no evidence of obvious LC and HCC. LC is defined by clinical criteria and confirmed by imaging technologies (magnetic resonance imaging, ultrasonography, or computed tomography) and no evidence for HCC. HCC met the criteria from The American Association for the Study of Liver Diseases guidelines (2) and was confirmed by computed tomography and magnetic resonance imaging. Before our study, none of the patients had received anticancer therapy.
All patients were classified into subgroups according to alanine aminotransferase (ALT), HBV-DNA, and HBeAg status. Based on the level of ALT, HBV-infected patients were divided into two groups: ALT normal group (ALT ≤30 IU/L for men; ALT ≤19 IU/L for women) and ALT abnormal group (ALT >30 IU/L for men; ALT >19 IU/L for women). In addition, the levels of HBV-DNA were used to classify patients into HBV-DNA >103 copies/mL and HBV-DNA <103 copies/mL. HC was enrolled with HBsAg, HBeAg, HBeAb, and HBcAb negative, normal clinical parameters.
Exclusion criteria for our study were as follows:(1) coinfection with hepatitis A virus, hepatitis C virus, hepatitis D virus, hepatitis E virus, human immunodeficiency virus, or syphilis; (2) LC and HCC not caused by HBV; (3) patients with nonalcoholic steatohepatitis, alcoholic liver disease, autoimmune disease, primary biliary cirrhosis, or primary HCC; (4) patients with evidence of pregnancy, hematological disease, diabetes mellitus, or other disease that could influence our results; and (5) age younger than 18.
Immunophenotyping and flow cytometry
For T cell phenotypic analysis, the following antibodies were used: CD3-PerCp (BD), CD4-FITC (BD), CXCR5-APC (BD), IL-21-PE (BD), PD-1-PE (eBioscience), ICOS-PE (eBioscience), and FOXP3-PE (eBioscience).
Fifty microliter whole blood were incubated with CD3,CD4,CXCR5, and ICOS/PD-1 at 4°C for 30 min for analyzing frequency of circulating CD4+CXCR5+ T cells and percentage of PD-1, ICOS on those cells, then lysed erythrocytes with lysis solution at room temperature for 10 min, and washed with PBS twice.
For cTFR-like T cell and cTFH-like T cell analysis, the antibodies of CD4, CXCR5, Foxp3, and FoxP3 fix/perm kit were used according to the product manual.
Intracellular cytokine staining
Expression of IL-21 on cTFH was stimulated in vitro and added 500 μL whole blood with phorbol myristate acetate (PMA, 50 ng/mL) plus ionomycin (1 μg mL-1) at 37°C under 5% CO2 environment for 5 h. Fifty microliter stimulated blood was incubated with CD3, CD4, and CXCR5 at 4°C for 30 min, after erythrocyte lysis, fixated, and permeabilized at 4°C for 45 min. This was followed by wash with permeabilization buffer, incubated with antibodies of IL-21 at 4°C for 30 min, washed again, and flow cytometry staining buffer added for analysis.
The stained cells were analyzed by BD FACSCanto II flow cytometer (BD, Biosciences), and data were analyzed by FlowJo software (version 10.0.7).
Serological marker assays and HBV-DNA assays
Serological markers (HBsAg, anti-HBs, HBeAg, anti-HBe, and anti-HBc) were measured using commercially available kits (Roche, Germany). HBV-DNA was measured by real-time polymerase chain reaction (Roche). Serum levels of ALT, aspartate aminotransferase, total protein, albumin, γ -glutamyl transferase, and alkaline phosphatase were determined by Roche P800 (Roche). Serum level of alpha fetoprotein was determined by Roche E170 (Roche).
Statistical analyses
Data are presented as mean ± SD or median with interquartile (25th and 75th) according to data distribution. Student's t test and ANOVA were used for normal distribution data. Nonparametric tests (Kruskal–Wallis H, Mann–Whitney's U test) were used for non-normal distribution. Correlation analysis was evaluated by the Spearman rank correlation test.
All data were analyzed by SPSS software (version 23.0); two-sided p < 0.05 was considered statistical significance.
Results
Baseline and clinical characteristics of subjects
Baseline characteristics of patients in our study are presented in Table 1. Sixty-five patients with HBV infection and 20 HCs were enrolled. Infected patients were divided into CHB group (n = 24), LC group (n = 23), and HCC group (n = 18), according to the clinical information and imaging technologies. Compared with HCs, patients with HBV infection displayed expected differences in clinical parameters, including liver function, platelets (PLT), and HBV DNA (Table 1).
Mean ± SD or median with interquartile (25th and 75th) as appropriate.
Analysis of variance (ANOVA) or nonparametric test (Kruskal–Wallis H); p < 0.05 was considered statistically significant.
ALB, albumin; ALT, alanine aminotransferase; AST, aspartate aminotransferase; CHB, chronic hepatitis B; GGT, gamma-glut amyl trans peptidase; HBcAb, HBV core antibody; HBeAb, HBV e antibody; HBeAg, HBV e antigen; HBsAb, HBV surface antibody; HBsAg, HBV surface antigen; HBV, hepatitis B virus; HC, healthy control; HCC, hepatocellular carcinoma; LC, liver cirrhosis; NA, not available; PLT, platelets; TP, total protein; WBC, white blood cell.
Frequency of cTFR-like cells and the ratio of cTFR-like/cTFH-like cells were increased in patients with persistent HBV infection
To determine whether those cells were related to HBV persistent infection, flow cytometry was performed to analyze those cells in peripheral blood. We analyzed the circulating CD4+CXCR5+ T cells among HC group, CHB group, LC group, and HCC group. There was no difference of circulating CD4+CXCR5+ T cells among four groups (16.11 [12.3–18.5] for HC, 15.8 [12.1–20.2] for CHB, 18.9 [14.2–25.7] for LC, and 17.0 [14.5–21.3] for HCC, respectively, p = 0.300).
Gating strategies of cTFR-like cells (CD4+CXCR5+FOXP3+) and cTFH-like cells (CD4+CXCR5+FOXP3−) were based on expression characteristics (Fig. 1A)
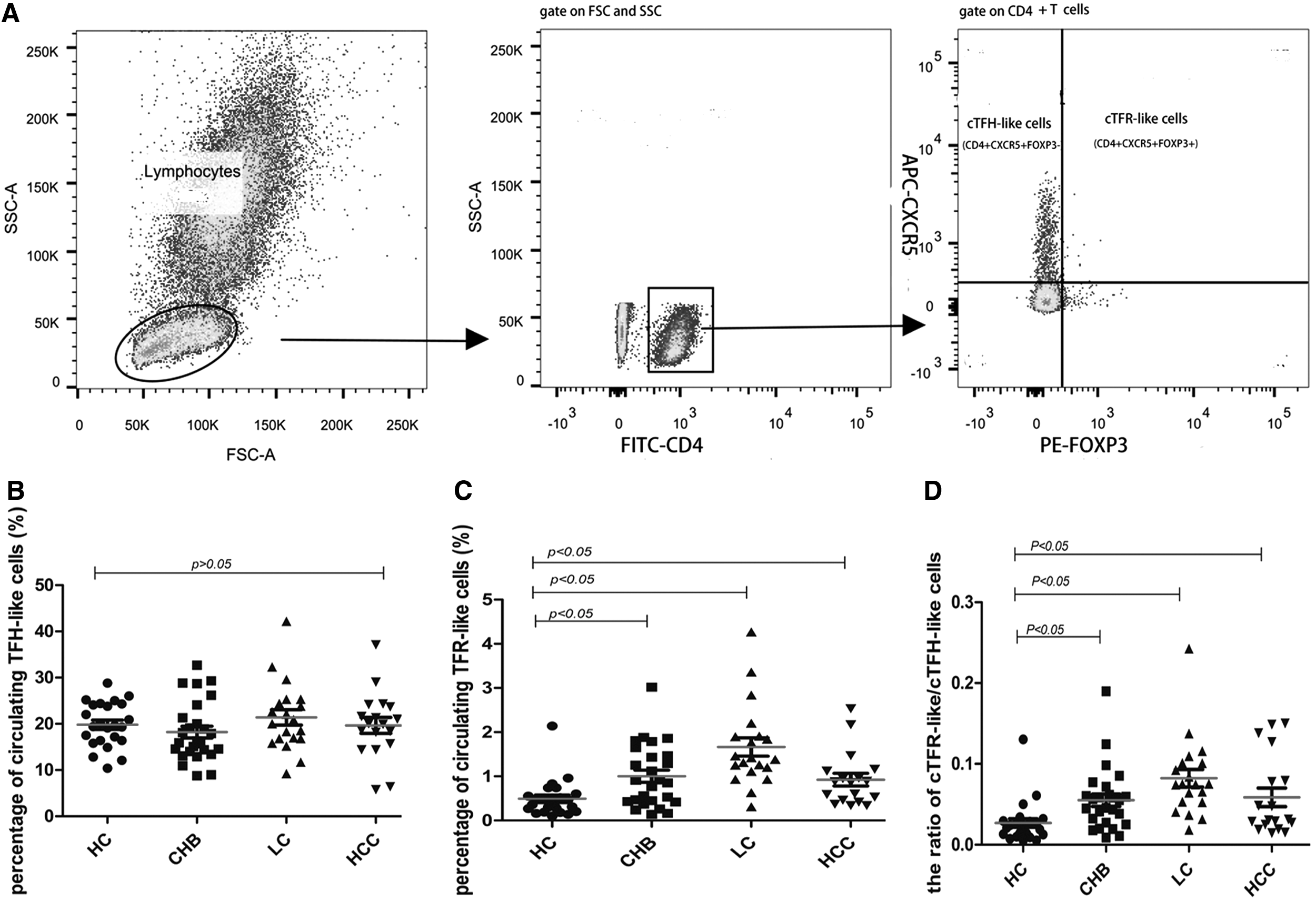
The frequencies of cTFH-like T cells and cTFR-like T cells in different groups.
There was no significant difference of cTFH-like cells among four groups (Fig. 1B). Frequency of cTFR-like cells was significantly increased in patients with HBV infection compared to HC. A significantly higher frequency of cTFR-like cells was observed in CHB group relative to the HC group (0.41 [0.16–0.68%] vs. 0.66 [0.48–1.09%], p = 0.002). In the LC group, cTFR-like cells were increased almost threefold more than the HC group (0.41 [0.16–0.68%] vs. 1.16 [0.54–2.2%], p < 0.001) (Fig. 1C). Moreover, correlation analysis showed that the frequency of circulating CD4+CXCR5+ T cells was strongly associated with cTFH-like cells (rs = 0.897, p < 0.000), but not with cTFR-like cells (rs = 0.177, p = 0.099).
The balance ratio of cTFR-like cells/cTFH-like cells in patients with HBV infection was significantly higher compared with the HC group: 0.046 [0.028–0.069] for the CHB group, 0.077 [0.027–0.115] for LC group, 0.054 [0.019–0.102] for HCC group, and 0.027 [0.010–0.038] for HC group (Fig. 1D).
Phenotype and function of circulating CD4+CXCR5+ cells in patients with chronic infection
To determine if the phenotypes and functions of circulating CD4+CXCR5+ cells were changed in patients with HBV infection, we tested the expression of inhibitory receptors PD-1 and costimulate receptors ICOS on circulating CD4+CXCR5+ cells and analyzed the percentage of IL-21-producing CD4+CXCR5+ cells after stimulation with PMA/ionomycin. The gating strategies for expression of PD-1, ICOS, and IL-21 on circulating CD4+CXCR5+ T cells are showed in Figure 2A.
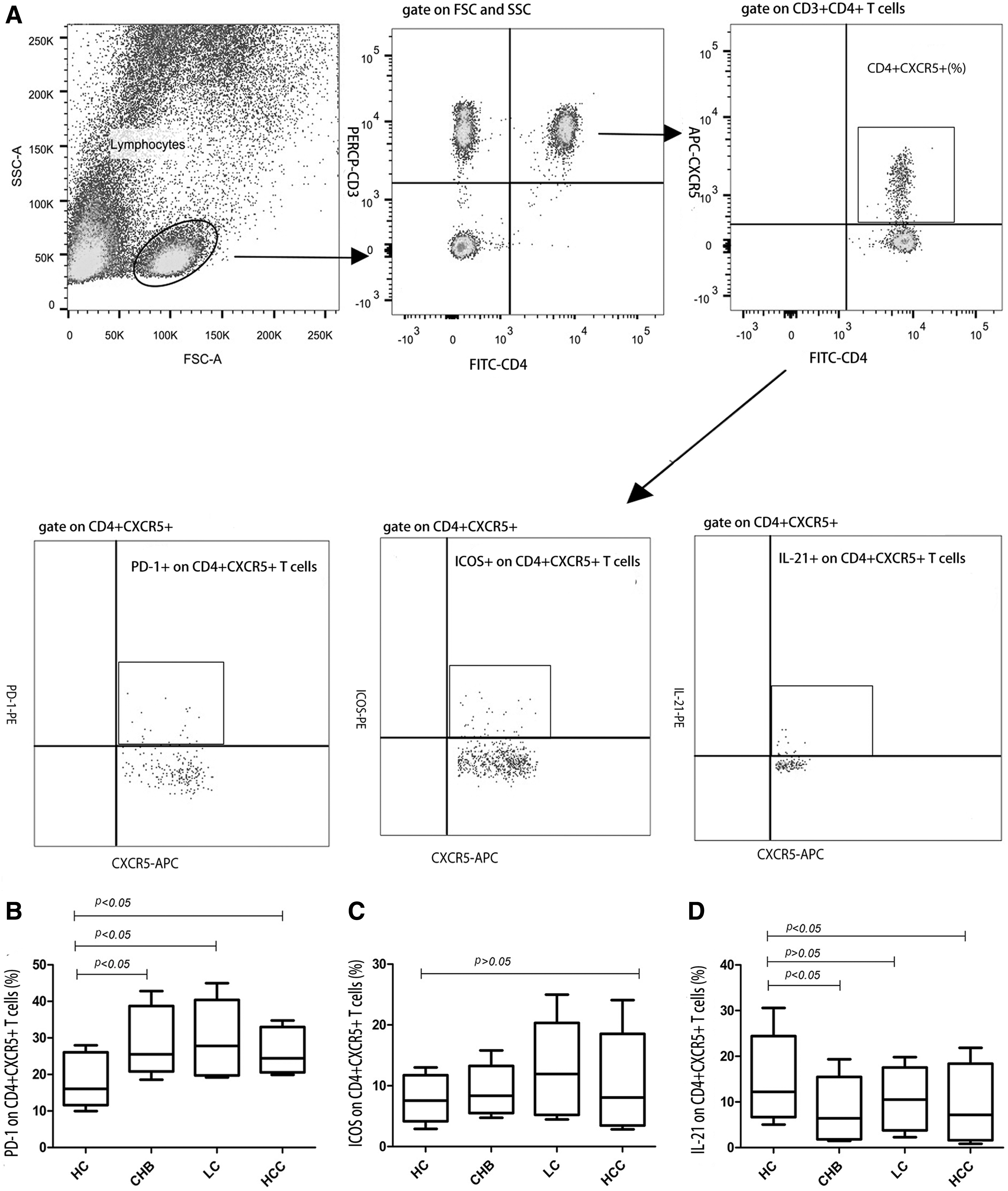
Both phenotype and function of circulating CD4+CXCR5+ T cells were changed in patients with chronic HBV infection.
The percentage of PD-1 on circulating CD4+CXCR5+ cells was much higher in the CHB group (25.5 [23.1–34.6%], p < 0.001), LC group (27.8 [20.2–35.8%], p < 0.001), and HCC group (24.5 [21.1–31.2%]; p = 0.002) than HC group (15.5 [13.1–24.2%], p < 0.001) (Fig. 2B). However, no differences of the ICOS on circulating CD4+CXCR5+ cells were observed among the four groups (7.6 [5.4–10.8%] for HC, 8.3 [6.3–10.7%] for CHB, 11.9 [6.0–15.7%] for LC, and 8.1 [4.1–13.0%] for HCC, p > 0.05) (Fig. 2C).
On the contrary, as seen in Figure 2D, a significantly decreased frequency of IL-21 on circulating CD4+CXCR5+ cells was found in the CHB group and HCC group compared with the HC group (12.8% ± 6.9% for HC group, 7.3% ± 4.1% for CHB group, and 9.1% ± 7.3% for HCC group, p < 0.001).
Spearman analysis showed that the percentage of PD-1 on circulating CD4+CXCR5+ cells was negatively correlated with the percentage of IL-21 on circulating CD4+CXCR5+ cells (rs = −0.240, p = 0.036).
Relationship of cTFH-like T cells and cTFR-like T cells with liver inflammation, HBV-DNA, and HBeAg seroconversion
To determine whether the balance of cTFR-like/cTFH-like T cells and the phenotype of circulating CD4+CXCR5+ T cells were involved in liver inflammation, levels of HBV-DNA, and HBeAg seroconversion, subgroup analysis was performed based on clinical information. The data are presented in Table 2.
ALT normal group (ALT ≤30 IU/L for men; ALT ≤19 IU/L for women). ALT abnormal groups (ALT >30 IU/L for men; ALT >19 IU/L for women). PD-1+/CD4+CXCR5+: percentage of PD-1 on circulating CD4+CXCR5+ T cells. ICOS+/cTFH: percentage of ICOS on circulating CD4+CXCR5+ T cells. IL-21+/cTFH: percentage of IL-21 on circulating CD4+CXCR5+ T cells. cTFH-like T cells: CD4+CXCR5+FOXP3−. cTFR-like T cells: CD4+CXCR5+FOXP3+.
Median with interquartile (25th and 75th); Mann–Whitney's U test; p < 0.05 was considered statistical significance.
ICOS, inducible co-stimulator; IL, interleukin; PD-1, programmed cell death-1; TFH, follicular helper T cell; TFR, follicular regulatory T cell.
First, we divided HBV infected patients into HBV-DNA <103 copies/mL and HBV-DNA ≥103 copies/mL groups. The frequencies of cTFH-like T cells and cTFR-like T cells were similar in two groups, while the frequency of CXCR5+PD-1+CD4+ cells was significantly increased in patients with HBV-DNA ≥103 copies/mL (2.8 [2.3–5.2%] vs. 5.3 [3.2–6.6%], p = 0.007) (Table 2).
Second, patients with HBV infection were divided into normal level (ALT ≤30 IU/L for men; ALT ≤19 IU/L for women) and abnormal level (ALT >30 IU/L for men; ALT >19 IU/L for women). Significantly increased cTFH-like T cells and CXCR5+PD-1+CD4+ T cells were observed in patients with elevated ALT. In addition, Spearman analysis revealed that the percentage of cTFR-like T cells and the ratio of cTFR-like/cTFH-like T cells were positively correlated with APRI (rs = 0.461, p < 0.001 for cTFR-like T cells and rs = 0.404, p < 0.001 for cTFR-like/cTFH-like) (Fig. 3B, D) and FIB-4 (rs = 0.483, p < 0.001 for cTFR-like T cells and rs = 0.390, p < 0.001 for cTFR-like/cTFH-like) (Fig. 3A, C).
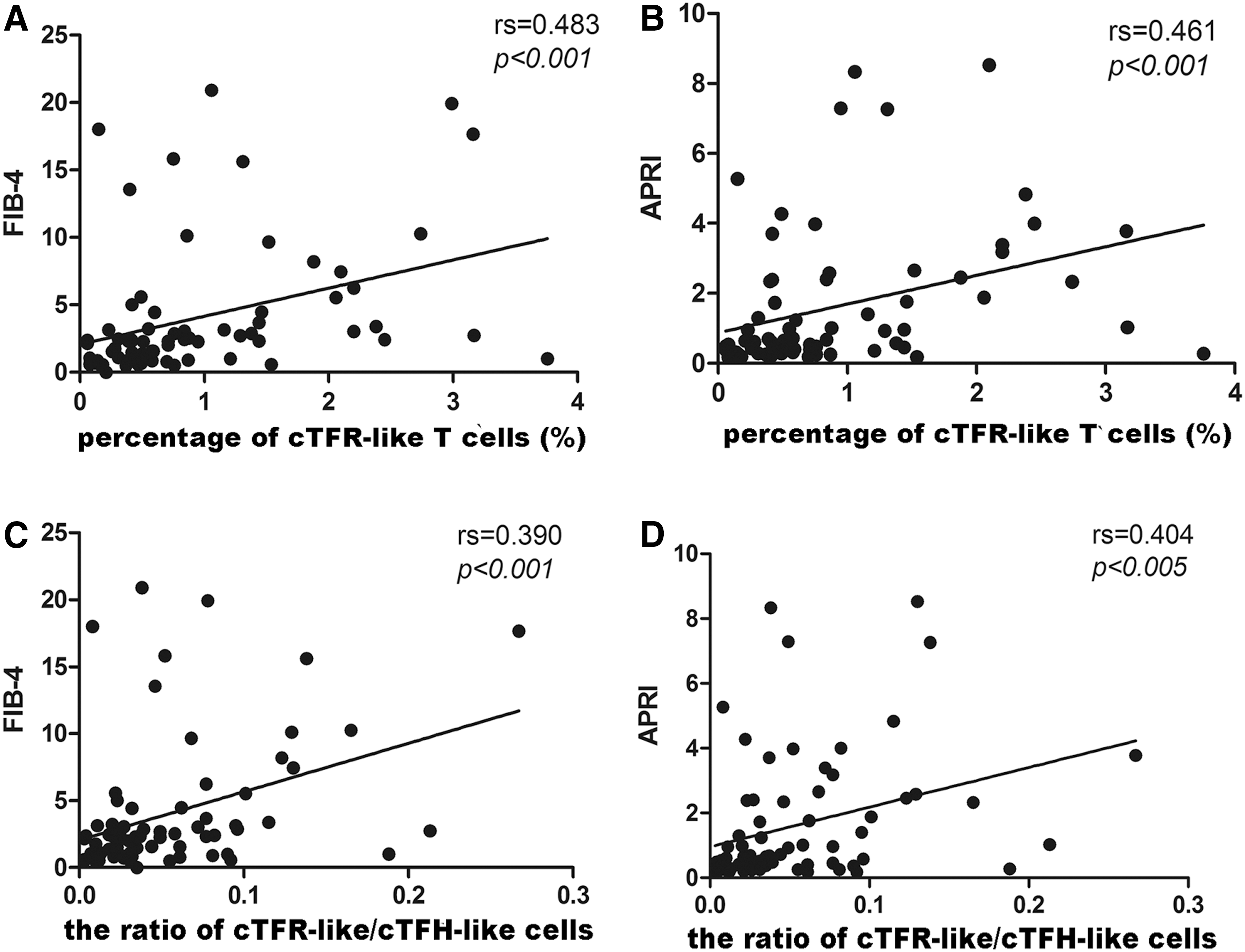
Correlation between cTFR-like T cells and clinic parameters in patients with persistent HBV infection.
Third, subgroup analysis was performed on the HBeAg status of patients. High frequencies of IL-21 on circulating CD4+CXCR5+ T cells and CXCR5+IL-21+CD4+ T cells were observed in the HbeAg negative group (Table 2).
Discussion
Effective control of HBV infections requires cooperation between cellular and humoral immune response. T cells play a major role in the immune pathogenesis. Defective T cell function results in viral persistence and chronic liver inflammation (1,34). Multiple subsets of T cells contribute to virus clearance and liver pathogenesis.
T cells participate in the humoral immune response when TFH and TFR are present in follicles. TFH has a role in virus infection and several types of cancers (12). The ratio of TFR/TFH is considered to be the potential mechanism in control of GC reactions as well (6,18,23,30). However, understanding of TFH and TFR effects on HBV chronic infection and disease progression remains poor.
We first demonstrated that frequency of cTFR-like T cells and ratio of cTFR-like/cTFH-like increased significantly in patients with chronic HBV infection. Wang's study showed that cTFR was higher in the CHB group than the HC group (40). Moreover, the frequency of cTFR-like T cells was positively correlated with FIB-4 and APRI, indicating that increased cTFR-like T cells might be correlated with severe liver fibrosis. We suggest that increased cTFR skews the balance toward immune tolerance during HBV persistent infection.
All results suggested that increased cTFR-like T cells and imbalanced cTFR-like/cTFH-like ratio might contribute to virus chronic infection and chronic liver damage. In chronic HIV infection, TFR expansion was benefited by regulatory DCs, TGF-β signaling, expression of CXCR5+ in Treg, and resistance to apoptosis (26). Defective function of pDC, higher concentration of TGF-β in sera, and dramatically increased Treg were observed in HBV infected patients, which might promote upmodulation of the cTFR cells (24,27,36). However, the potential mechanisms of the expansion of cTFR in HBV infection were still unknown.
Second, our study indicated that functional impairment of circulating CD4+CXCR5+ T cells was related to chronic infection and development of HBV-related diseases. Correlation analysis showed that the circulating CD4+CXCR5+ T was strongly associated with cTFH-like T cells. Studies pointed out that analyzing the circulating CD4+CXCR5+ T cells might be helpful for assessing the situation of TFH (17,20,35,39). Patients with chronic HBV infection exhibited a higher percentage of PD-1 on circulating CD4+CXCR5+ T cells and functional deficiency in producing IL-21. Expansion of TFR might lead to a functional suppression of TFH in HCV infection (5).
Researchers concluded that the possible functions of TFR were as follows: TFR could secrete suppressive cytokines and disrupt interactions between TFH and B cells. Any direct suppressive effect on GC B cells would have an indirect effect on TFH, and vice versa (31). However, whether dysfunction of cTFH was caused by increased TFR needs further verification in HBV infection.
In addition, the expression of PD-1 on circulating CD4+CXCR5+ T cells was weakly negatively correlated with IL-21 on circulating CD4+CXCR5+ T cells. We speculated that increased expression of PD-1 on circulating CD4+CXCR5+ T cells might limit the proliferation and activation of circulating CD4+CXCR5+ T cells during persistent exposure in virus. Other studies have shown that the PD-1 signal in TFH might have a role in its differentiation and GC reaction. PD-1 deficiency in TFH occurs in antibody-mediated autoimmune diseases. Shinmei's study demonstrated that excess TFH was found in PD-1 deficiency mice (15). PD-1: PD-L1/PD-L2 pathway regulates T cell activation and tolerance, PD-1 dephosphorylating of TCR signaling and reducing CD28-mediated PI3K activation. Expression of PD-1 and connection with its ligands regulate the threshold for T cell activation (16). However, lack of PD-1 signaling might result in less expression of IL-4 and IL-21 (11). Future studies on the relationship between the expression of PD-1, IL-21, and IL-4 and dysfunctional TFH are needed.
More important, IL-21 plays a critical role in regulating immunoglobulin production. Decreased frequencies of IL-21-producing TFH were observed in HCV and HIV chronically infected patients (10,35). However, the role of IL-21-producing TFH in HBV remains controversial. IL-21-production in CXCR5+CD4+ T cells increased in CHB patients, but decreased in patients with HCC (13,20,45). In our study, reduced IL-21-producing circulating CD4+CXCR5+ T cells were observed in CHB group and HCC group. Such discrepancy might be due to the age of patient, treatment for HBV, and potential gender factors. Other researchers have noted that the combination of ICOS signal and lower PD-L1 in GC B cells could increase IL-21 secretion (9,38). An HBV-related HCC study reported that reduced IL-21 in TFH was caused by upregulated hepatic PD-L1 (45).
The percentage of ICOS on circulating CD4+CXCR5+ T cells between HCs and patients with chronic infection was similar in our study; combined with enrichment PD-1 in circulating CD4+CXCR5+ T cells, IL-21 might decrease during infection. In addition, TFR might interact with TFH, which could be the mechanism of TFH dysfunction (5,26,43). Interaction between TFR and plasma cells could modulate the GC reaction and IL-21 in TFH through an undefined mechanism. Increasing the ratio of TFR led to decreased production of IL-4 and IL-21 on TFH (26). It may be reasonably concluded that increased cTFR and imbalance of cTFR/cTFH might involve disability of cTFH in producing of IL-21 in chronic HBV infection.
Increased action of IL-21 on circulating CD4+CXCR5+ T cells might contribute to HbeAg seroconversion in both Hou's and our studies. T cell-derived IL-21 maintained transcription factor AP4 expression, which promoted selection and sustained expansion of GC B cells, and expressed IL-21 in TFH which may be closer to the dark zone in mouse model (4,41). IL-21 signaling profoundly affects the GC reaction and influences the affinity maturation of antibody (38,46). Recently, IL-21 was found to have a reinforcing effect on humoral immunity by restricting TFR cell proliferation (14). Further study is needed to explore the role of TFH-derived IL-21 in HBV infection and disease progression.
There were many limitations in our study. First, considering the complication of liver biopsy, our study analyzed the cells of peripheral blood instead of liver or follicles. Although few studies have shown the strong association between TFH and TFR in blood and in tissue, the cell culture experiment and mouse mold study still need to verify our results. Second, the numbers of patients in different groups were not enough to divide patients into subgroups for more detailed research. Third, more experiments are still required to explain and verify our results, the mechanisms of how cTFR exert their suppressive effect and why aberrant expression of cTFR in CHB is still unknown. Fourth, how the subgroups of circulating CD4+CXCR5+ T cells participated in HBV chronic infection and disease progression still need to be clarified.
Taken together, increased cTFR-like T cells and an imbalance ratio of cTFR-like/cTFH-like cells were observed in patients with HBV infection. In addition, impairment of circulating CD4+CXCR5+ T cells might play a key role in weakening immune response to HBV and contributing to the progress of disease. Knowing the mechanisms of those immune cells may help us to understand the immune pathogenesis of HBV chronic infection. Further functional studies may provide a potential target for HBV treatment.
Ethical Approval
All procedures performed in the study involve human participants and were in accordance with the ethical standards of the institutional research board and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Footnotes
Acknowledgments
The authors thank Dr. Qi Zhang for the guidance of their experiment. The authors thank all the patients who participated in their study. The authors also thank the hospital staff for giving them useful advice about their experiments. This work was supported by the National Natural Science Foundation of China (No. 81273256 and No. 81571561).
Author Disclosure Statement
No competing financial interests exist.