
Abstract
Emerging evidences showed the promoting role of spleen in the development of cirrhosis and hepatocellular carcinoma, in the context of portal hypertension and hypersplenism due to hepatitis B virus-related cirrhosis. In this study, we tried to explore the precise mechanism of how spleen regulates this process from the serum cytokines profile level. Compared with the 16 healthy subjects, the RayBio Human Cytokine Antibody Array identified 136 cytokines differentially expressed in 36 cirrhotic patients. Splenectomy resulted in significant changes in 28 cytokines. Differentially expressed cytokines were mainly involved in cellular processes, responses to stimuli, immune processes, binding, extracellular regions, and extracellular matrix. These cytokines were mainly enriched in cytokine–cytokine receptor interactions, Jak-STAT and MAPK signaling pathways, and pathways in cancer. These results may provide new clues to the function of the spleen, and modulation of cytokine expression maybe a potential alternative therapeutic strategy for these patients.
Introduction
W
Hepatitis B virus (HBV) infection is the main cause of liver cirrhosis, and both of them are major risks of hepatocellular carcinoma (HCC), especially in China. Portal hypertension (PH) is a progressive complication of cirrhosis that can cause splenomegaly, hypersplenism, ascites, gastroesophageal varices, and variceal hemorrhaging. The contribution of the spleen in liver regeneration is important because these two organs are anatomically closely associated by the portal circulation. Splenectomy plus pericardial devascularization is a conventional surgical procedure for the treatment of PH and hypersplenism, which can reverse pancytopenia, improve liver function, and reduce portal pressure and the risk of variceal hemorrhaging. The improved liver fibrosis following splenectomy has been reported in basic and clinical research (10,12). Furthermore, splenectomy may cause beneficial immunological changes in cirrhosis patients (9,12,23). Splenectomy in combination with liver resection has been reported to reduce tumor recurrence and improve prognoses in cirrhosis patients with HCC (5,33). The spleen stiffness also even can be used to evaluate both liver disease severity and the development of cirrhosis complications (8). However, the role of the spleen in PH and HBV-related cirrhosis remains unclear. The precise mechanism of how the spleen regulates the liver regeneration has also not yet been determined. Exploration of this mechanism would be helpful in finding new strategy to improve liver regeneration.
Regarding the underlying mechanisms, splenic-derived cell and cytokine may play a pivotal role through liver–spleen crosstalk (1,14,29). To date, only few cytokines have been linked to the effect of splenectomy in liver cirrhosis, including transforming growth factor (TGF)-β1. Whether the splenectomy can induce changes in serum cytokines pattern has not been well investigated.
In this study, we focus on changes in the serum cytokines expression pattern between normal control and cirrhotic patients before and after splenectomy and evaluate the potential effect of splenectomy on HBV cirrhosis patients with PH and hypersplenism by in silico analysis.
Patients and Methods
Patients
From December 2012 to June 2014, a total of 36 consecutive HBV-related cirrhosis patients with PH and hypersplenism, who underwent splenectomy at the Second Affiliated Hospital of Xi'an Jiaotong University, were enrolled in this prospective study. The clinical characteristics of the cirrhotic patients and healthy controls are shown in Table 1. All the patients suffered varying degrees of splenomegaly, and the majority of them had moderate–severe esophagogastric varices as revealed by upper gastrointestinal radiography or endoscopy examinations. The indications for splenectomy included endoscopic treatment-resistant esophagogastric varices with or without variceal hemorrhage, a tendency to bleeding and infection due to hypersplenism (platelet count <50 × 109/L and/or leukocyte count <3.0 × 109/L), and upper abdominal discomfort due to an enlarged spleen. Patients were excluded from the study if any of the following confounding or complicating factors was present: coinfection with hepatitis C virus or HIV, cancer, autoimmune disorder, previous heart disease, chronic kidney disease, chronic respiratory disease, diabetes or hyperlipidemia, intravenous drug addiction, alcohol abuse, or interferon/nucleoside antiviral therapy 6 months before splenectomy. The control group populations were composed of 16 sex- and age-matched healthy individuals who underwent routine health checkups in our hospital and were without liver disease and the exclusion criteria listed above. This group included 11 males and 5 females, with a mean age of 49.6 ± 10.2 years (Table 1). Six of the 36 patients and four of the 16 healthy controls were used for microarray screening, as described below. Validation studies were performed using all subjects.
Data are expressed as n (%) or mean ± standard deviation.
HBeAg, hepatitis B e antigen; PLT, platelet count; WBC, white blood cell count; ALT, alanine transaminase; PT, prothrombin time.
All patients were informed about the benefits and risks of the surgery. Written and signed consent was obtained from all subjects. The study was approved by the Institutional Ethics Committee of the Second Affiliated Hospital of Xi'an Jiaotong University.
Serum collection
Peripheral venous blood was collected into procoagulation tubes before and 2 weeks after splenectomy. The tubes were left undisturbed at room temperature for 30 min and then centrifuged at 2,000 rpm for 10 min at 4°C. The supernatants (sera) were divided into 200 μL aliquots and stored at −80°C. One aliquot was used for each assay to avoid multiple freeze/thaw cycles.
Cytokine array hybridization
The protein antibody array, AAH-CYT-G4000, which was designed and manufactured by RayBiotech, Inc. (Norcross, GA), contains antibodies for 274 different serum cytokines. Each of the antibodies has two replicates, and each array contains positive and negative control antibodies. The experiment was performed by Wayne Biotechnology (Shanghai, China) according to their established protocol. The serum samples of six patients, before and 2 weeks after splenectomy, and four healthy controls were analyzed. In brief, four-fold diluted serum was incubated on the slide overnight at 4°C, and then, the hybridized factors were detected with biotin-conjugated secondary antibodies and Cy3-conjugated streptavidin. All slides were scanned using an Axon GenePix 4000B Microarray Scanner (Molecular Devices) and analyzed using GenePix Pro 6.0 software (Molecular Devices). The F532 median - B532 score was used. The results were then normalized using the positive controls.
Validation assays
The serum samples of 36 patients, collected before and 2 weeks after splenectomy, and 16 healthy controls, including those enrolled in array study, were used to verify the reliability of the cytokine arrays. The levels of brain-derived neurotrophic factor (BDNF), platelet-derived growth factor BB (PDGF-BB), matrix metalloproteinase 1 (MMP-1), beta-2-microglobulin (β2-M), vascular cell adhesion molecule-1 (VCAM-1), and epidermal growth factor (EGF) were measured using ELISA kits from Neobioscience (Beijing, China).
Bioinformatics analysis of cytokines
To visualize the differentially expressed cytokines, a heat map was generated using Cluster 3.0 and Tree View. Gene ontology (GO) analysis of the cytokines was performed using the PANTHER classification system (
Statistical analysis
The differential expression of cytokines was considered statistically significant when the ratio between samples was >1.5 or <0.65 and the p value was <0.05. Pearson correlations were used to evaluate the association between differentially expressed cytokines and clinical characteristics as appropriate. Other analyses were performed using GraphPad Prism 5.0 (GraphPad software, La Jolla, CA). Data are presented as the mean ± standard deviation. Differences in mean serum cytokine levels between healthy subjects and patients before and 2 weeks after splenectomy were tested by Student's t-test, with p values <0.05 considered statistically significant.
Results
Identification of differentially expressed cytokines
In the antibody-array data analysis, a total of 136 cytokines were identified to be differentially expressed (upregulated >1.5 time or downregulated <0.65 with p < 0.05) in the presplenectomy patient group compared with the control group (Supplementary Table S1; Supplementary Data are available online at
Compared with the presplenectomy sera, the postsplenectomy sera contained 28 differentially expressed cytokines (upregulated >1.5 time or downregulated <0.65 with p < 0.05), including 17 that were upregulated and 11 that were downregulated (Table 2). Some cytokines associated with cell proliferation, bone marrow hematopoiesis, and thymus activation such as BDNF, PDGF, vascular endothelial growth factor (VEGF), and thymus and activation-regulated chemokine (TARC) were significantly upregulated. Some cytokines associated with genesis, invasion, and metastasis of tumors such as VCAM-1, NCAM-1, and β2-M were significantly downregulated after splenectomy.
The hierarchical cluster analysis of these differentially expressed cytokines showed that the expression patterns are significantly changed in patients, compared with the normal control, but getting close to normal after splenectomy (Fig. 1).
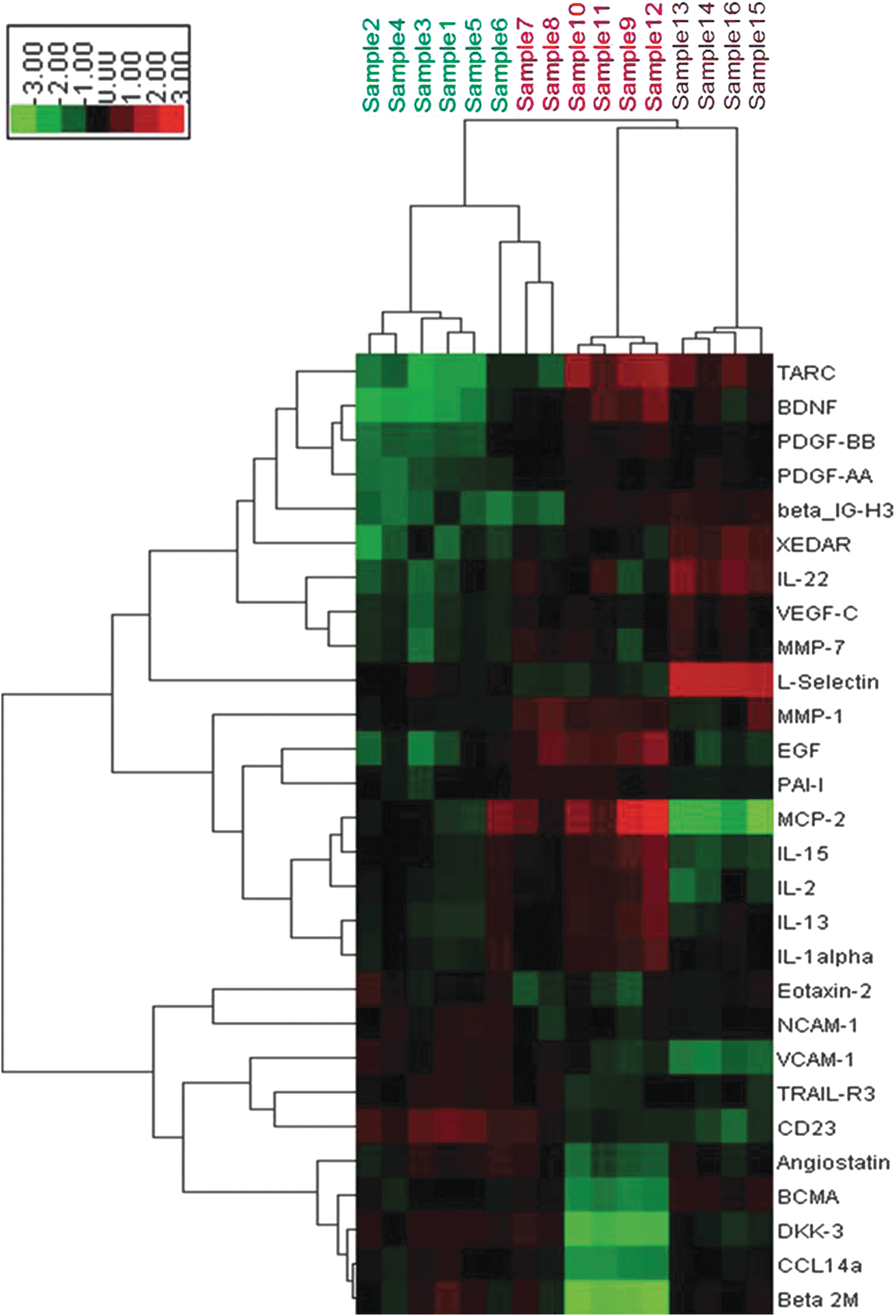
Hierarchical cluster analysis of the 28 cytokines differentially expressed in HBV-related cirrhosis patients before and 2 weeks after splenectomy. Red color: high expression; green color: low expression; black color: moderate expression. Samples 1–6 (green): presplenectomy; samples 7–12 (red): postsplenectomy; samples 13–16 (brown): healthy controls.
Validation by ELISA assay
To assess the reliability of the cytokine arrays, we selected six cytokines according to signal intensity and fold-change after splenectomy. They also involved in angiogenesis and tissue remodeling. Compared with presplenectomy samples, postsplenectomy samples contained significantly increased levels of BDNF (3.04 ± 1.82 vs. 13.97 ± 6.63 ng/mL, p < 0.001), EGF (10.10 ± 8.11 vs. 34.39 ± 14.23 pg/mL, p < 0.001), PDGF-BB (458.21 ± 154.11 vs. 4755.27 ± 1723.85 pg/mL, p < 0.001), and MMP-1 (957.99 ± 799.01 vs. 7744.87 ± 4598.47 pg/mL, p < 0.001) (Fig. 2A) and significantly decreased levels of β2-M (86.39 ± 24.51 vs. 50.15 ± 14.79 μg/mL, p < 0.001) and VCAM-1 (1557.91 ± 744.88 vs. 697.97 ± 169.24 ng/mL, p < 0.001) (Fig. 2B). These ELISA results were consistent with those from the cytokine arrays.
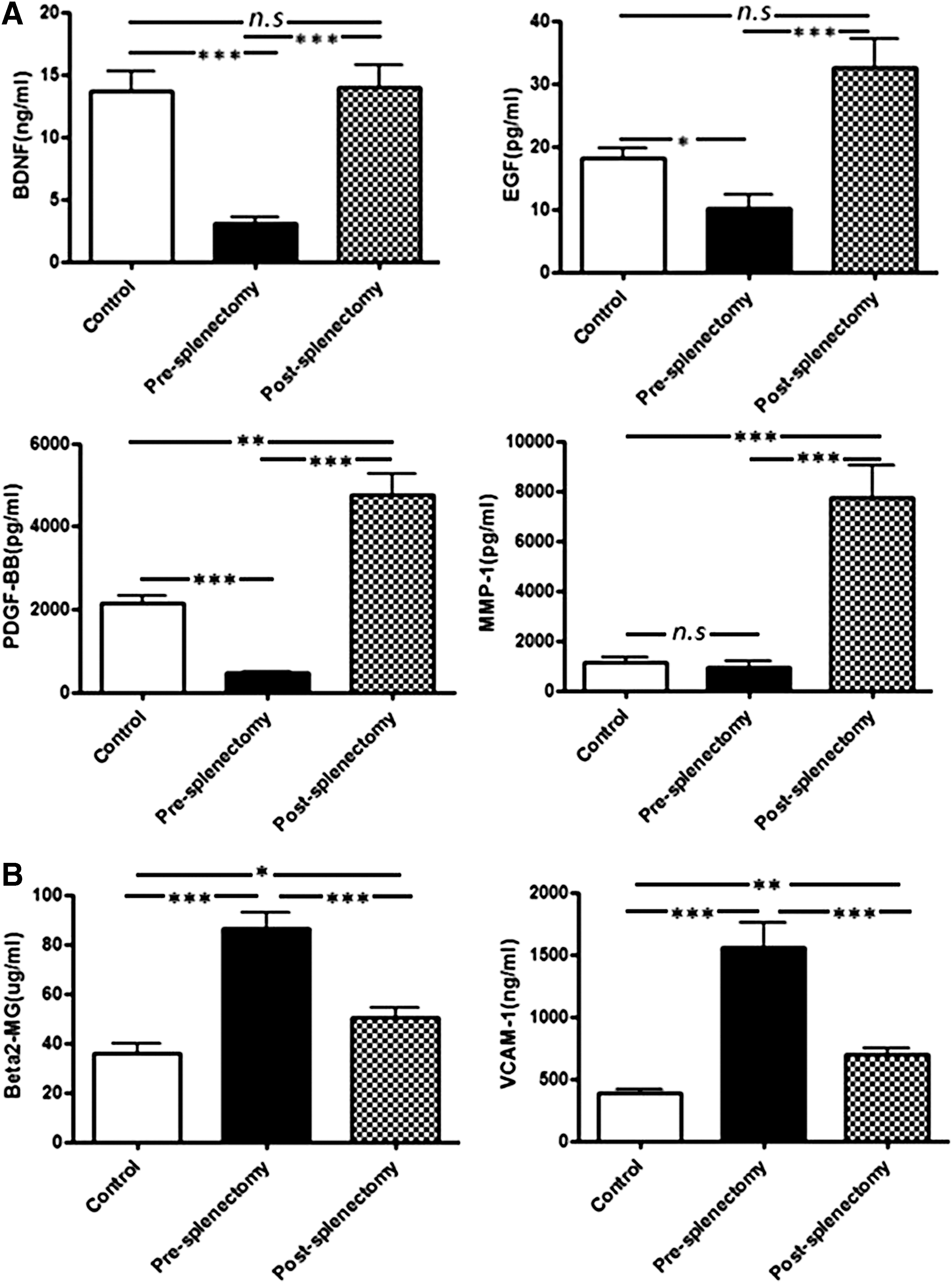
Verification of the cytokine arrays by ELISA.
Correlations and Bioinformatics analysis of differentially expressed cytokines
To understand the biological function of the differentially expressed cytokines, we first did the correlation analysis between the differentially expressed cytokines and the clinical characteristics of the patients. Among the 28 differentially expressed cytokines, the serum level of soluble VCAM-1 was negatively correlated with albumin (r = −0.752, p < 0.001), white cell count (r = −0.496, p = 0.002), and platelet count (r = −0.436, p = 0.008) and positively correlated with total bilirubin (r = 0.634, p < 0.001) in patients before splenectomy, while not in patients after splenectomy (Supplementary Fig. S2).
We further used the online tool, PANTHER, to classify the cytokines according to biological process, molecular function, and cellular component.
The 136 differentially expressed cytokines in the presplenectomy group (compared with the control group) were classified into 14 biological process groups, the most common of which were cellular process (GO: 0009987; 23.1%) and response to stimulus (GO: 0050896; 17.6%). The cytokines were classified into six molecular function groups, the most common of which was binding (GO: 0005488; 60%). The cytokines were classified into five cellular component groups, the most common of which was extracellular region (GO: 0005576; 58.5%) (Supplementary Fig. S3).
The 28 differentially expressed cytokines in the postsplenectomy group (compared with the presplenectomy group) were classified into 12 biological process groups, the most common of which were cellular process (GO: 0009987; 20.4%) and response to stimulus (GO: 0050896; 17.3%). In addition, many of the 28 cytokines were involved in immune system processes (GO: 0002376; 11.2%) and biological regulation (GO: 0065007; 11.2%). The cytokines were classified into six molecular function groups, the most common of which was binding (GO: 0005488; 44.1%). The cytokines were classified into five cellular component groups, the most common of which were extracellular region (GO: 0005576; 56.2%) and extracellular matrix (GO: 0031012; 25%) (Fig. 3).
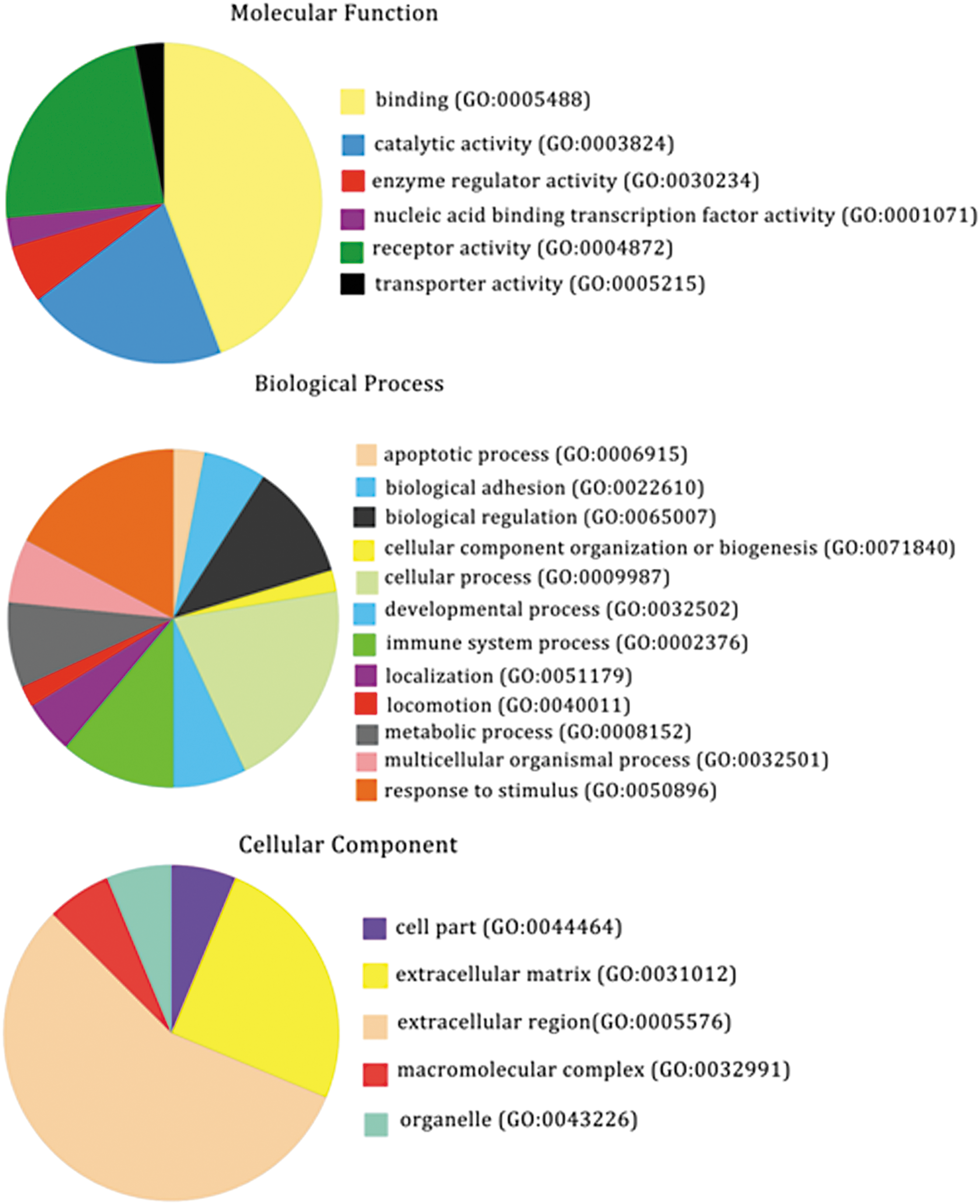
Functional distribution of the 28 cytokines differentially expressed in HBV-related cirrhosis patients before and 2 weeks after splenectomy. Cytokines were classified by GO analysis according to their biological process, molecular function, and cellular component. GO, gene ontology.
The KEGG pathways of the two sets of differentially expressed cytokines are shown in Table 3 and Supplementary Table S2. The differentially expressed cytokines were mainly enriched in cytokine–receptor interaction pathways, the Jak-STAT signaling pathway, the MAPK signaling pathway, and pathways in cancer.
KEGG, Kyoto Encyclopedia of Genes and Genomes.
Discussion
Cytokines are a diverse group of small soluble proteins composed of interleukins, chemokines, growth factors, and interferons, which play important roles in the physiological and pathological processes of chronic viral hepatitis (13). In addition to the cytokines, the array used in this study also included other soluble proteins, such as angiogenesis factors, proteases, apoptotic factors, soluble receptors, adhesion molecules, and adipokines.
Studies showed that cytokines in the spleen may be involved in the progression of liver cirrhosis through liver-spleen crosstalk (1,14,29). TGF-β is often considered the predominant profibrotic signal, and TGF-β1 is the major fibrogenic cytokine in liver fibrosis (14,21). Spleen-derived TGF-β1 has already been indicated as inhibitory factor in liver regeneration (30). Among the differentially expressed 136 cytokines identified in the presplenectomy patient group, the serum levels of TGF-β1, TGF-β2, and TGF-α were significantly increased. Meanwhile, matrix metalloprotease (MMP)-9 and MMP-7 expression were significantly decreased in cirrhotic patients (Supplementary Table S1). MMP-9 expression is negatively regulated by TGF-β at the transcriptional level, and TGF-β has an inhibitory effect on the expression of MMP-9 (24). Studies showed that TGF-β signaling also can activate ligand-independent downstream PDGFRα signaling, and TGF-β may be also induced by PDGFRα (31). Thus, there may be a reciprocal regulation between TGF-β and PDGF signaling in hepatic fibrosis. In our study, PDGFRα has the most obvious change with a 21-fold upregulated in cirrhotic patients compared with the healthy subjects.
In addition to the TGF-β, leptin and leptin-R also play pivotal role in profibrogenic responses in the liver through lipid metabolism (11,31). We also observed the significantly increased expression in cirrhosis patients. Other cytokines upregulated in cirrhosis patients, including FGF-6, IL-9, and SDF-1, also have been linked to the development of fibrosis or cirrhosis (2,7,16,26). These can partially explain why the hypertensive spleen is related to the progression of liver cirrhosis.
Upregulated VCAM-1 expression in chronic liver disease has also been reported, and it has been correlated to the hyperdynamic circulation in liver cirrhosis (19). In this study, we found the serum level of soluble VCAM-1 negatively correlated with albumin, white cell, and platelet count and positively correlated with total bilirubin in cirrhotic patients. These supported a potential role of sVCAM-1 as a marker of severity of liver disease.
Splenectomy increases peripheral platelet counts, and that platelets and platelet-derived serotonin reduce liver fibrosis and promote liver regeneration by upregulating HGF and its receptor c-Met (15,22,32). Our antibody-array results also showed that EGF and HGF were upregulated 6.57- and 1.23-fold change after splenectomy, respectively. MMP-1 and MMP-7 were also significantly upregulated after splenectomy. Leptin receptor levels showed a trend to decrease after splenectomy. Changes in these cytokines maybe favorable to reduce liver fibrosis, enhance hepatocyte proliferation, and prevent the progression of liver cirrhosis. Some of the other cytokines upregulated after splenectomy were related to cell proliferation, hematopoiesis, and thymus activation such as BDNF, PDGF, VEGF, and TARC; these may be associated with other immune organs and may promote bone marrow hematopoiesis, wound healing, and tissue reconstruction after surgery.
Splenectomy had been reported as an independent protective factor for HCC development in cirrhosis patients (20). Our unpublished data show a low cumulative incidence of HCC in 1.1%, 7.4%, and 13.0% after 1, 3, and 5 years after surgery, respectively. In this study, we also found that splenectomy could reverse the abnormal expression pattern of cytokines in some extent. Furthermore, these changes of cytokines that belonged to EGF, ERK/MAPK, PI3K/AKT, and TGF-β signaling pathways may play critical regulatory roles for early HCC transformation (4,6,17,18,27). Our results showed that the differentially expressed cytokines were mainly involved in cytokine–receptor interaction pathways and pathways in cancer-like Jak-STAT and MAPK (Table 3 and Supplementary Table S2). The changes in these cytokines can maintain at least 6 months after splenectomy (data no shown). These alterations may play a protective effect in the development of HCC.
This study has several limitations. First, the follow-up period after splenectomy: as the cytokine expression pattern is a highly dynamic changed process, it can start as early as 6 h after splenectomy in rats. In patients with HCC, TGF-β1 levels slightly increased in PB samples 1 month after splenectomy and then decreased and subsequently returned to the level measured before splenectomy in 1 year (23). So, we pick up 2 weeks as our observation time point and further validate some cytokines at 6 month after splenectomy. These follow-up periods might be optimized with large scale study in the future to elucidate the corresponding mechanism. Next, this study only focused on cytokine alteration of the effects of splenectomy, whereas splenectomy also induces cellular component alterations. Combination of both aspects would provide a comprehensive picture of the effect of splenectomy on liver disease.
In conclusion, the present study unveiled serum cytokine profiles in HBV-related cirrhosis patients with PH and hypersplenism, indicating a potential role of the hypertensive spleen in the progression of liver disease. Furthermore, the changes in cytokine levels following splenectomy maybe potential advantageous to reduce liver fibrosis and accelerate liver regeneration as well as reduce the risk of HCC. Modulation of cytokine expression pattern in spleen maybe a potential alternative therapeutic strategy for these patients.
Footnotes
Acknowledgments
We thank for the technical supports from Shanghai H Wayne Biotechnologies in the protein antibody array.
This work was supported by the National Natural Science Foundation of China (No.30901268, 91442122, 81470131, 81270504).
Author Disclosure Statement
The authors have no conflict of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.