
Abstract
Evolutionarily conserved pattern recognition receptors, including Toll-like receptors (TLRs) recognize pathogen-associated molecular patterns (PAMPs) that are present in microbes. PAMPs induce several pathways downstream of TLRs that lead to induction of antiviral responses. The objective of this study was to investigate the stimulatory effect of various PAMPs (in the form of TLR ligands) in reducing Marek's disease virus (MDV) infection in chicken embryo fibroblast cells (CEFs). To this end, CEFs were pretreated with Pam3CSK4, Poly(IC), lipopolysaccharide (LPS), and CpG ODN as TLR2, TLR3, TLR4, and TLR21 ligands, respectively for 24 h followed by infection with MDV. The results indicated that pretreatment with Poly(IC) resulted in a robust reduction (by about 81%) of MDV infection in CEFs at 96 h postinfection while a moderate reduction was observed with treatment of Pam3CSK4 (35%), LPS (26%), and CpG ODN (23%) PAMPs. Transcriptional analysis of gene expression in CEFs demonstrated that all TLR ligand treatments and MDV infection significantly increased the expression of type I interferons, interleukin (IL)-1β, interferon regulatory factor 7 (IRF7), interferon induced protein with tetratricopeptide repeats 5 (IFIT5), and myxoma-resistance protein (Mx). Further studies are needed to explore the mechanism by which PAMPs, particularly the TLR3 ligands could reduce MDV infection in CEFs, which may play an important role in controlling the replication of MDV in chicken.
Introduction
M
Evolutionarily conserved pattern recognition receptors, including Toll-like receptors (TLRs) recognize pathogen-associated molecular patterns (PAMPs) that are present in microbes. Several studies including those conducted by our group have explored the use of TLR ligands (TLR-Ls) as prophylactic agents or as adjuvants to provide protective immunity against various infectious diseases in several species, including chickens (29,30). Several TLRs such as TLR1 type 1 and 2; TLR2 type 1 and 2, TLR3, TLR4, TLR5, TLR7, TLR15 and TLR21 are reported to be present in various tissues of chickens (23,37). Among them, TLR3 and TLR21 detect nucleic acid based TLR-Ls. Viral double-stranded RNA (dsRNA) and synthetic dsRNA [polyinosinic:polycytidylic acid known as Poly(IC)] are recognized by TLR3. TLR21 detects unmethylated CpG motifs present in bacterial and viral DNA. TLR2 and TLR4 detect lipid based ligands. Peptidoglycan, lipopeptide and lipoproteins of bacteria including synthetic lipoproteins such as PAM3CSK4 are recognized by TLR2, whereas bacterial lipopolysaccharide (LPS) is recognized by TLR4 and bacterial flagellin by TLR5.
In regard to MDV, similar to other herpesviruses, glycoproteins and double-stranded DNA of the virus may be recognized by TLR2 and TLR21, respectively, with the latter being the homolog of TLR9 in mammals (27). In addition, TLR4 might become activated directly through viral components of MDV or indirectly via MDV induced danger signals as it happens in other herpesviruses (39). TLR3 recognizes dsRNA, which is a transitional product in the formation of DNA viral progeny (3). Therefore, it is possible for the chicken innate immune system to recognize dsRNA by TLR3 during MDV infection. Importantly, we have previously demonstrated that the synthetic TLR3 ligand, Poly(IC), can reduce tumor formation in MDV-infected chickens (28).
The interaction of TLRs and TLR-Ls initiates downstream signaling pathways via activation of transcription factors that lead to the expression of several immune system genes, including cytokines, chemokines, adhesion molecules, and receptors. Interactions of TLR2, TLR4 and TLR5 with their ligands lead to the production of proinflammatory cytokines whereas interaction of TLR7 and TLR9 with their ligands leads to the secretion of type 1 interferons (IFNs) (24). Interactions of type 1 IFNs with their receptors result in the induction of interferon-stimulated genes (ISGs), many of which have antiviral activities, such as 2′, 5′-oligoadenylatesynthase (OAS), dsRNA-dependent protein kinase (PKR), interferon-induced protein with tetratricopeptide repeats 5 (IFIT5–also known as ISG58), and myxoma-resistance protein (Mx). These ISGs play a critical role in the inhibition of virus replication at different levels of the virus life cycle.
The aim of this study was to investigate the ability of TLR-Ls to reduce replication of MDV in cultured cells. To this end, we used a fluorescent tagged MDV in chicken embryo fibroblast cells (CEFs) and various TLR-Ls were screened to determine their efficacy in reducing virus replication. In addition, to shed light on some of the underlying mechanisms of TLR-Ls effects on MDV replication, expression of several immune system genes was examined in treated and infected CEFs.
Materials and Methods
Cells and virus
Specific pathogen-free (SPF) eggs were obtained from the Animal Disease Research Institute, Canadian Food Inspection Agency (Ottawa, Ontario, Canada). SPF eggs were incubated at the recommended temperature and humidity conditions. CEFs were prepared from 11-day-old chicken embryos as described previously (33). Enhanced green fluorescent tagged MDV, recombinant RB1B strain (vUL47-EGFP), originated in Dr. Benedikt Kaufer's lab (Freie Universität, Berlin, Germany) was obtained from Dr. Masahiro Niikura (Simon Fraser University, Burnaby, Canada) and used in this study to infect CEFs. All experiments involving chickens were approved by the Animal Care Committee, University of Guelph and complied with institutional regulations.
TLR ligands
Synthetic triacylated lipoprotein, Pam3CSK4, was purchased from InvivoGen (San Diego, CA). LPS from Escherichia. coli 0111: B4, Poly(IC) and synthetic class B CpG ODN 2007 [5′-TCGTCGTTGTCGTTTTGTCGTT-3′] with phosphorothioate backbone and non-CpG ODN (NCpG) [5′-TGCTGCTTGTGCTTTTGTGCTT-3′] (8) were purchased from Sigma–Aldrich (Oakville, ON, Canada). All ligands were resuspended in endotoxin-free sterile water.
Experimental design
First, uninfected CEFs were cultured and harvested for RNA extraction to determine the presence of TLR2, TLR3, TLR4, TLR5, TLR7, and TLR21. Next, CEFs were cultured and seeded on 96-well plate at 1 × 105/mL in RPMI-1640 medium (Invitrogen, Burlington, ON) supplemented with 10% heat-inactivated fetal bovine serum, 2% chicken serum, 200 U/mL penicillin, 80 μg/mL streptomycin, and 5% tryptose phosphate buffer. Monolayers of CEFs were stimulated with Pam3CSK4 0.1 μg/mL as a low dose and 1 μg/mL as a high dose, Poly(IC) 2.5 μg/mL as a low dose and 25 μg/mL as a high dose, LPS 0.1 μg/mL as a low dose and 1 μg/mL as a high dose, CpG ODN 0.5 μg/mL as a low dose and 5 μg/mL as a high dose; and NCpG 5 μg/mL for 24 h or left untreated. Stimulated cells were washed and infected with vUL47-EGFP MDV at MOI 0.1. CEFs were collected at 96 and 120-hour postinfection (hpi) for flow cytometry analysis to determine the MDV-infected cells. In the subsequent experiment, selected TLR-Ls were used to treat the CEFs before MDV infection and cells were collected for RNA extraction to determine gene expression at 6, 12, and 24-hour poststimulation (hps); and 24 (48 hps) and 48 (72 hps) hpi.
Flow cytometry
CEFs were harvested at 96 and 120 hpi and resuspended in FACS buffer (PBS with 1% BSA). Cells were stained with Live/dead fixable near-IR dead cell stain kit (Life technology, Eugene, OR) for exclusion of dead cells and incubated for 30 min on ice. At the end of the incubation, CEFs were washed and resuspended in FACS buffer. MDV-infected CEFs were determined by identifying EGFP MDV in FITC channel and the data were acquired using a FACSCanto flow cytometer (BD Biosciences, Mississauga, ON, Canada).
All samples within this experiment were analyzed individually using the FlowJo software (Tree Star, Ashland). Initial gating involved all live cells acquired within an SSC-A vs. FSC-A plot. Then through a two-step process involving FSC-W vs. FSC-H followed by SSC-W vs. SSC-H, all doublets were eliminated to avoid false positive stains. Final percentages were subsequently determined by gating on the cells that showed positive signals in the emission spectrum range of 530/30.
RNA extraction and cDNA synthesis
CEFs from two wells were pooled for extraction of RNA and considered as one replicate. There were six replicates. RNA was extracted using TRIzol reagent (Life Technologies, Burlington, Canada) according to manufacturer's protocol. RNA quantity and quality were determined using the NanoDrop® ND-1000 spectrophotometry (NanoDrop Technologies, Wilmington, DE). Ten micrograms of extracted RNA were treated with DNase enzyme (Ambion, Austin, TX) according to the manufacturer's protocol. Then, cDNA was synthesized from 1 μg of DNase treated RNA using Superscript II (Life Technologies). Synthesized cDNA was diluted at 1:10 ratio with nuclease-free water.
Real-time polymerase chain reaction
Real-time polymerase chain reaction (PCR) was performed using SYBR green dye in LightCycler 480 II (Roche Diagnostics, Laval, Canada) to quantify the relative expression of target genes. Chicken β actin gene was used as the reference gene to calculate the relative expression of all selected genes in this study. Primer sequences of target and reference genes are listed in Table 1. The primers were synthesized by Sigma–Aldrich.
IFIT, interferon-induced protein with tetratricopeptide repeats; IFN, interferon; IL, interleukin; IRF, interferon regulatory factor; Mx, myxoma resistance protein; TLR, Toll-like receptor.
Statistical analysis
To determine the statistically significant difference in MDV-infected CEFs, data were analyzed using one-way ANOVA followed by Dunnett's multiple comparison test to compare the pretreated group with infection only control group in GraphPad Prism version 6.04. For gene expression analysis, REST (Relative Expression Software Tool) software v2009 (Qiagen Valencia, CA) was used to calculate fold changes and standard errors. Data are presented as the mean fold change (±standard error of the mean) of the relative gene expression in the TLR-Ls treated groups and MDV infection only control group compared with PBS control group. If p value was ≤0.05, it was considered statistically significant.
Results
Expression of TLR transcripts in CEFs
The results of this study indicated that TLRs 2, 3, 4, 5, 7, and 21 were expressed by CEFs (Fig. 1). Constitutive expression of TLRs from highly expressed to minimally expressed was as follows: TLR2, TLR4, TLR21, TLR3, TLR7, and TLR5, respectively.
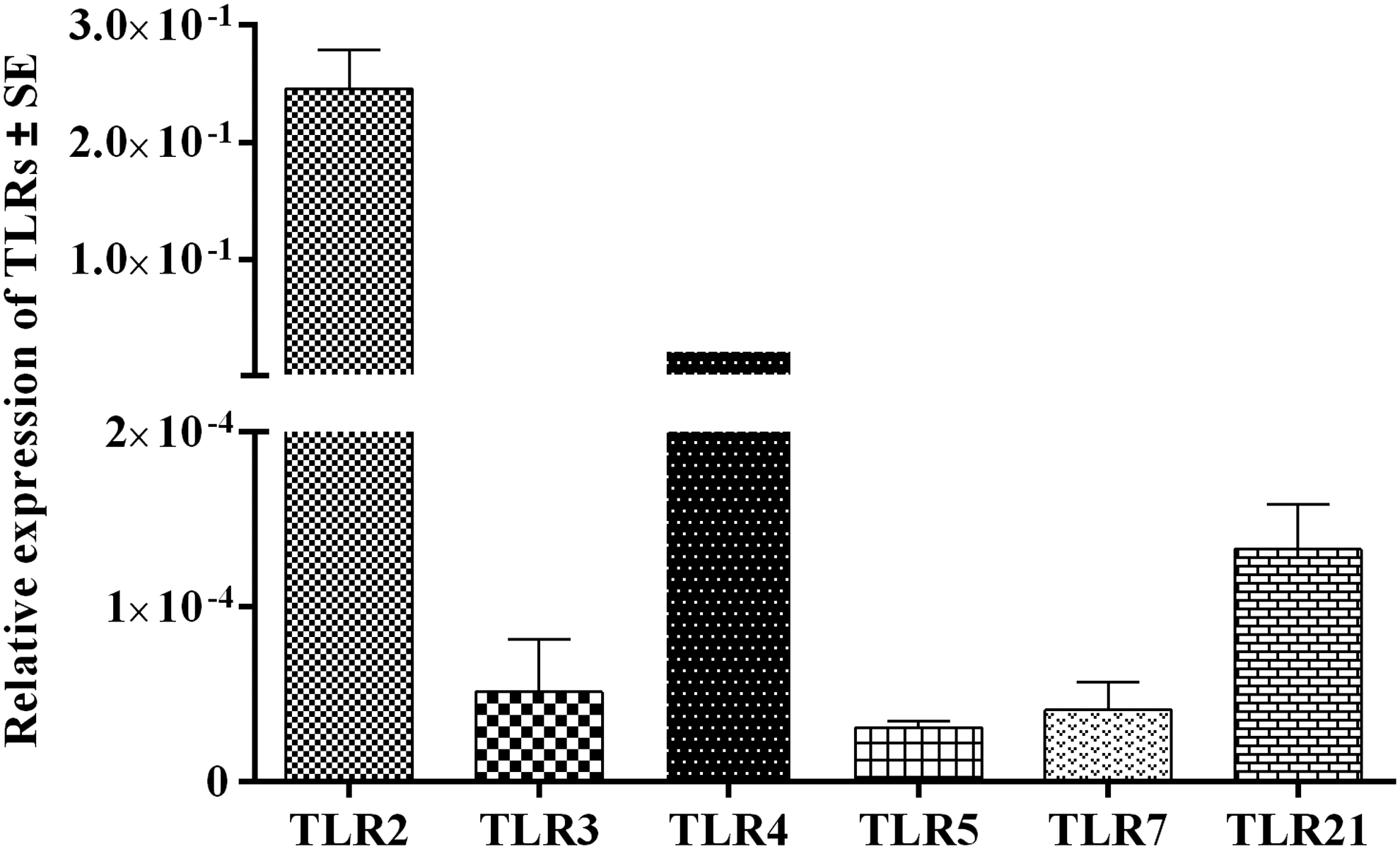
Relative gene expression of TLR transcripts in CEFs. Gene expression of TLRs 2, 3, 4, 5, 7 and 21 transcripts in CEFs relative to β-actin. CEF, chicken embryo fibroblast cell; TLR, Toll-like receptor.
Reduction of MDV infection by TLR-Ls in CEFs
TLR-Ls pretreated CEFs were infected with vUL47-EGFP MDV at 24 hps. The percentage of MDV-infected CEFs was determined by flow cytometry at 96 and 120 hpi (Fig. 2). Pretreatment with TLR-Ls significantly reduced the percentage of MDV-infected CEFs at one or both time points (Figs. 3A, B). Pretreatment with low and high doses of Poly(IC) (Figs. 2 and 3A) and low dose of Pam3CSK4 significantly reduced the percentage of MDV-infected CEFs compared with the untreated group at both 96 and 120 hpi (Fig. 3A). In addition, pretreatment with high dose of Pam3CSK4 significantly reduced the percentage of MDV-infected CEFs at 120 hpi (Fig. 3A).
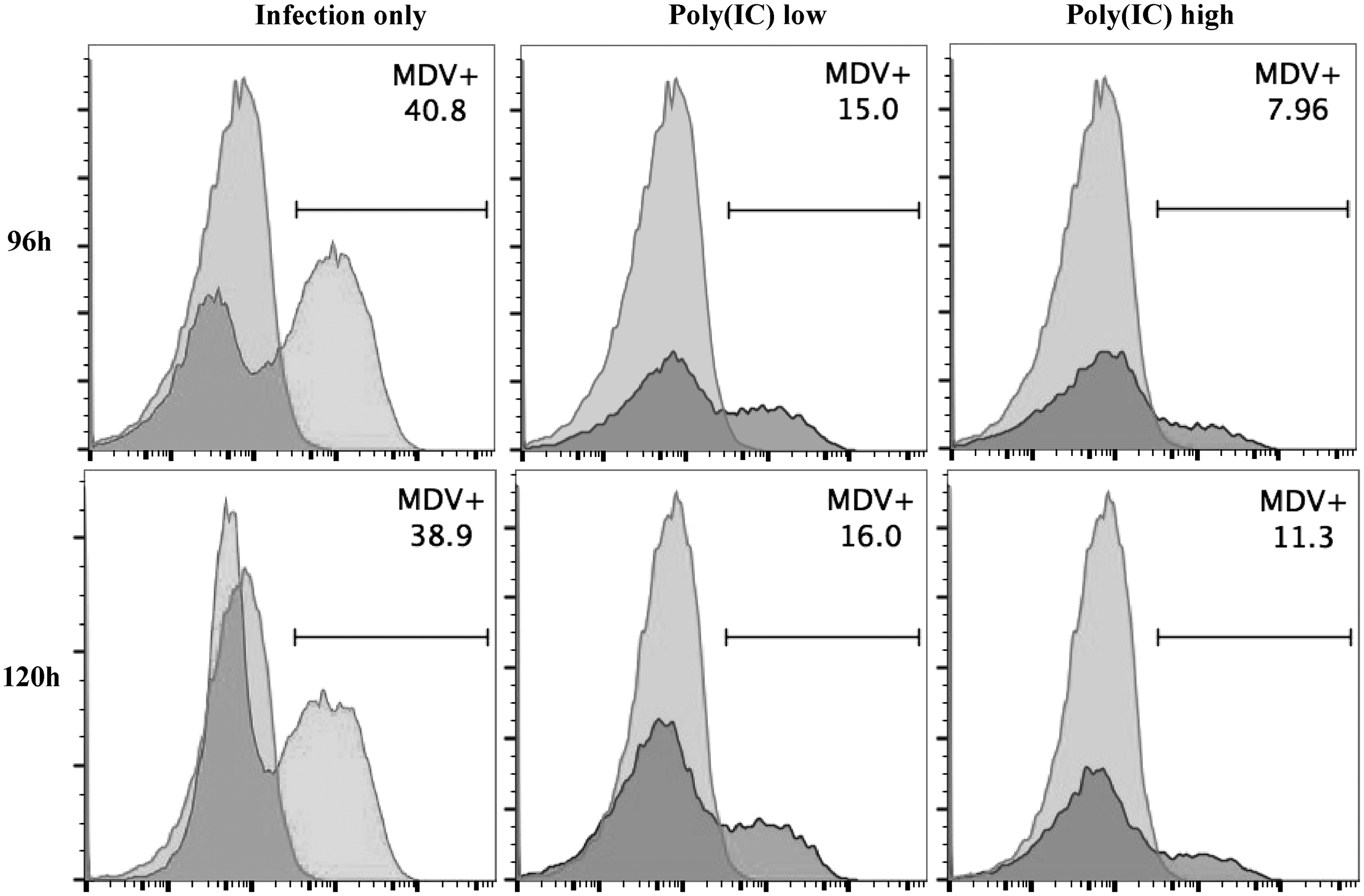
EGFP MDV-infected CEFs following pretreatment with TLR-Ls. Illustrative flow cytometry plot shows the percentage of MDV infected CEFs at 96 and 120 hpi following pretreatment with Poly(IC) for 24 h and infection with EGFP MDV along with the MDV infection only (untreated CEFs but infected) control group. EGFP positive cell population in the histograms are depicted in dark grey while the EGFP negative population is in light grey shades. EGFP, enhanced green fluorescent protein; hpi, hour postinfection; MDV, Marek's disease virus; TLR-Ls, TLR ligands.
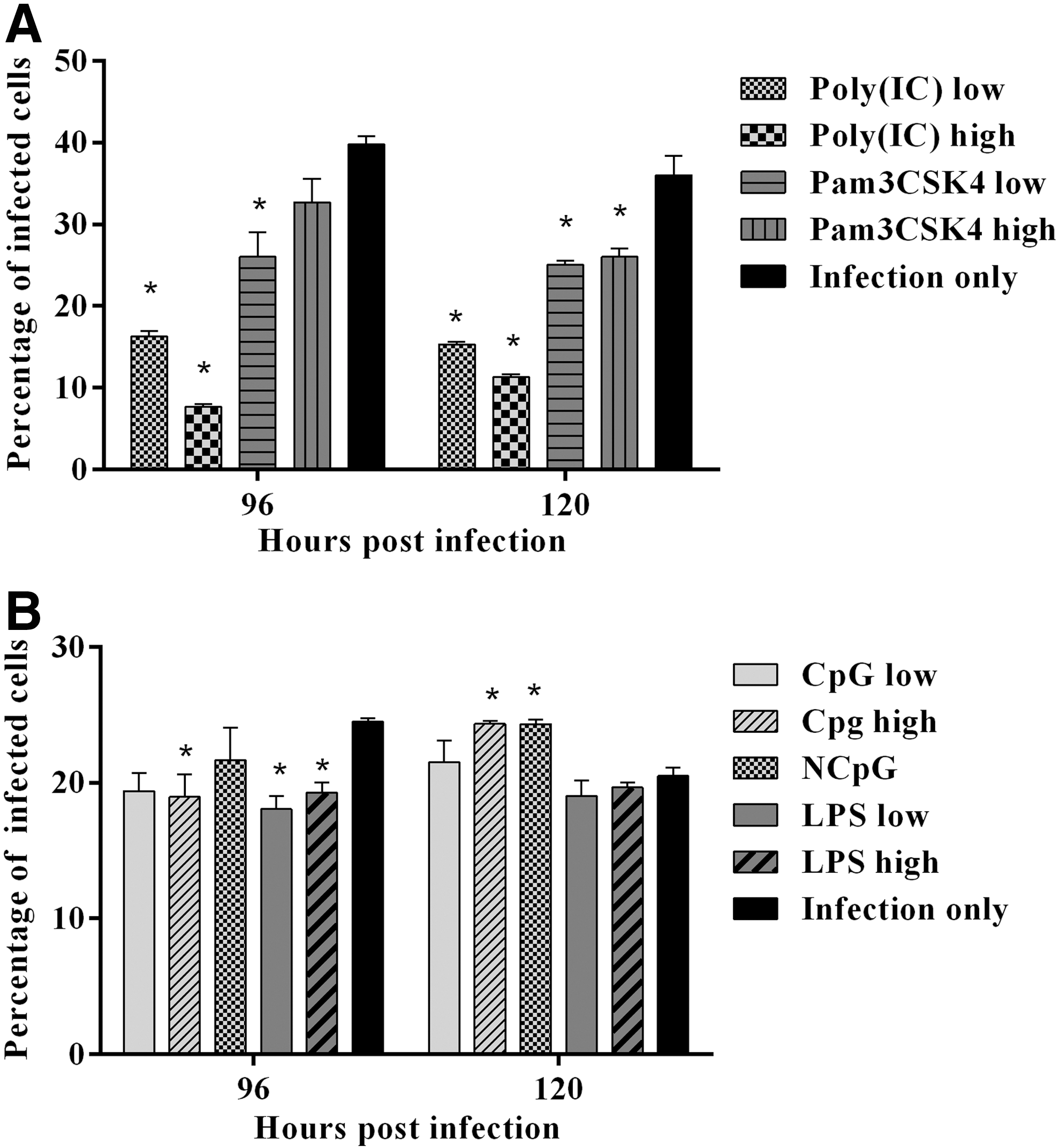
Reduction of MDV-infected CEFs by TLR-Ls pretreatment. CEFs were pretreated with
Pretreatment with high dose of CpG ODN, low and high doses of LPS significantly reduced the percentage of MDV-infected CEFs at 96 hpi compared with the untreated group (Fig. 3B). However, at 120 hpi, pretreatment with a high dose of CpG ODN and NCpG unexpectedly significantly increased MDV-infected CEFs compared with the untreated group. There was no significant difference in MDV infection in NCpG treated CEFs compared with that of CpG treated CEFs.
Based on the significant percentage of reduction of MDV-infected CEFs (Table 2), high dose of Poly(IC) (81% and 69%), low dose of Pam3CSK4 (35% and 31%) and low dose of LPS (26%) were selected for further experiment to investigate the involvement of related cytokine genes and ISGs.
hpi, hour postinfection; LPS, lipopolysaccharide.
Induction of cytokine genes and ISGs expression
CEFs were pretreated with high dose of Poly(IC), low dose of Pam3CSK4 and low dose of LPS for 24 h and infected with vUL47-EGFP MDV. Expression of cytokine genes and ISGs was analyzed at 6, 12, 24, 48 (24 hpi), and 72 hps (48 hpi) with TLR-Ls. Stimulation of CEFs with Poly(IC) significantly upregulated the expression of IFN-α gene at all time points except at 6 hps (Fig. 4A). In addition, Pam3CSK4 and LPS significantly upregulated the expression of IFN-α gene at 48 and 72 hps (Fig. 4A). Further, MDV infection alone significantly upregulated IFN-α gene expression at 48 hps (Fig. 4A). The expression of IFN-β gene was significantly upregulated by Poly(IC) pretreatment at all time points except 24 hps (Fig. 4B). All TLR-L pretreatments and MDV infection only control significantly upregulated IFN-β gene expression at 48 and 72 hps (Fig. 4B). In addition, Pam3CSK4 pretreatment significantly upregulated IFN-β gene expression at 12 hps (Fig. 4B).
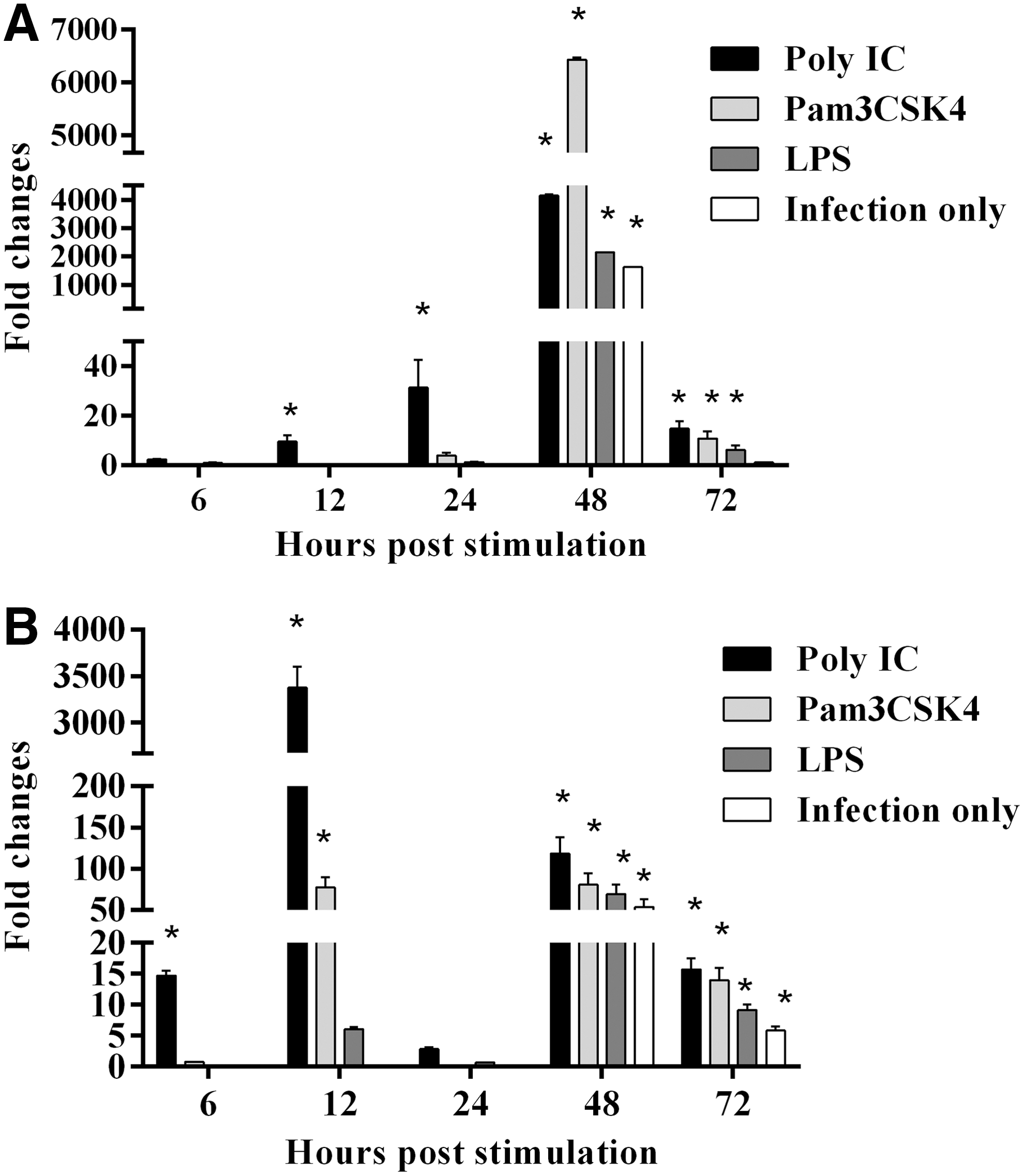
Relative expression of IFN-α and IFN-β in TLR-Ls pretreated, MDV-infected CEFs. CEFs were pretreated with Pam3CSK4, Poly(IC) and LPS, and infected with EGFP MDV. There were MDV infection only (Untreated CEFs but infected) and medium control groups.
Pretreatment with TRL-Ls significantly upregulated the expression of interleukin (IL)-1β gene at all time points except 24 hps at which LPS pretreatment only upregulated IL-1β gene (Fig. 5A). In addition, MDV infection also significantly upregulated IL-1β gene expression at 48 and 72 hps. Poly(IC) pretreatment significantly upregulated interferon regulatory factor 7 (IRF7) gene expression in all time points except 72 hps (Fig. 5B). Further, all TLR-Ls pretreatment and MDV infection only control significantly upregulated IRF7 gene expression at 48 hps (Fig. 5B).
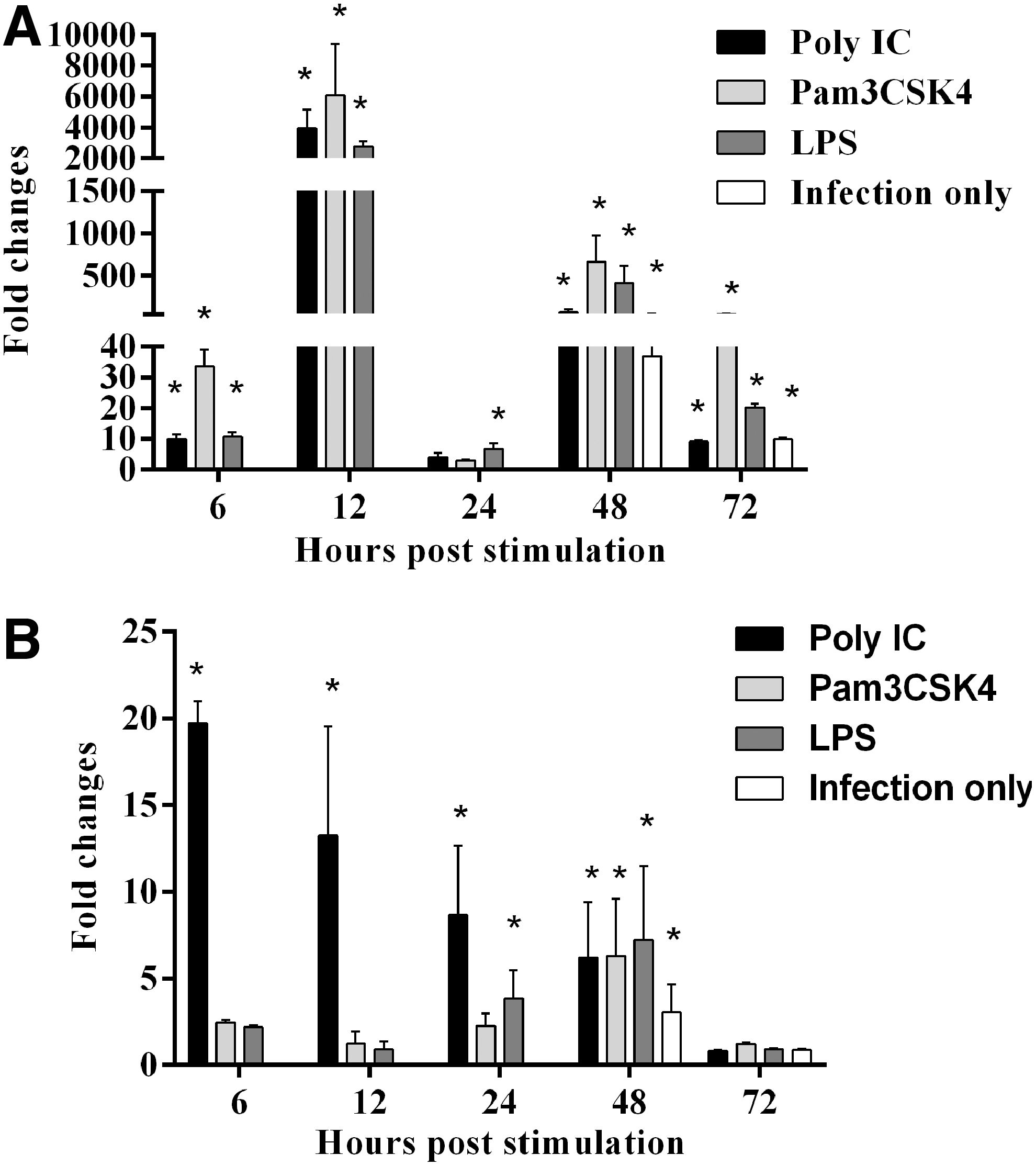
Relative expression of IL-1β and IRF7 in TLR-Ls pretreated, MDV-infected CEFs. CEFs were pretreated with Pam3CSK4, Poly(IC), and LPS, and infected with EGFP MDV. There were MDV infection only (Untreated CEFs but infected) and medium control groups.
Poly(IC) pretreatment significantly upregulated IFIT5 gene expression at all time points (Fig. 6A). Pam3CSK4 pretreatment significantly upregulated IFIT5 gene at 6, 24, and 48 hps (Fig. 6A). LPS pretreatment and MDV infection only control significantly upregulated IFIT5 gene expression at 48 and 72 hps (Fig. 6A). Mx gene expression was significantly upregulated by Poly(IC) pretreatment at all time points except 6 hps (Fig. 6B). Pam3CSK4 and LPS pretreatment significantly upregulated Mx gene expression at 48 and 72 hps (Fig. 6B). Furthermore, Mx gene expression was significantly upregulated by MDV infection at 48 hps (Fig. 6B).
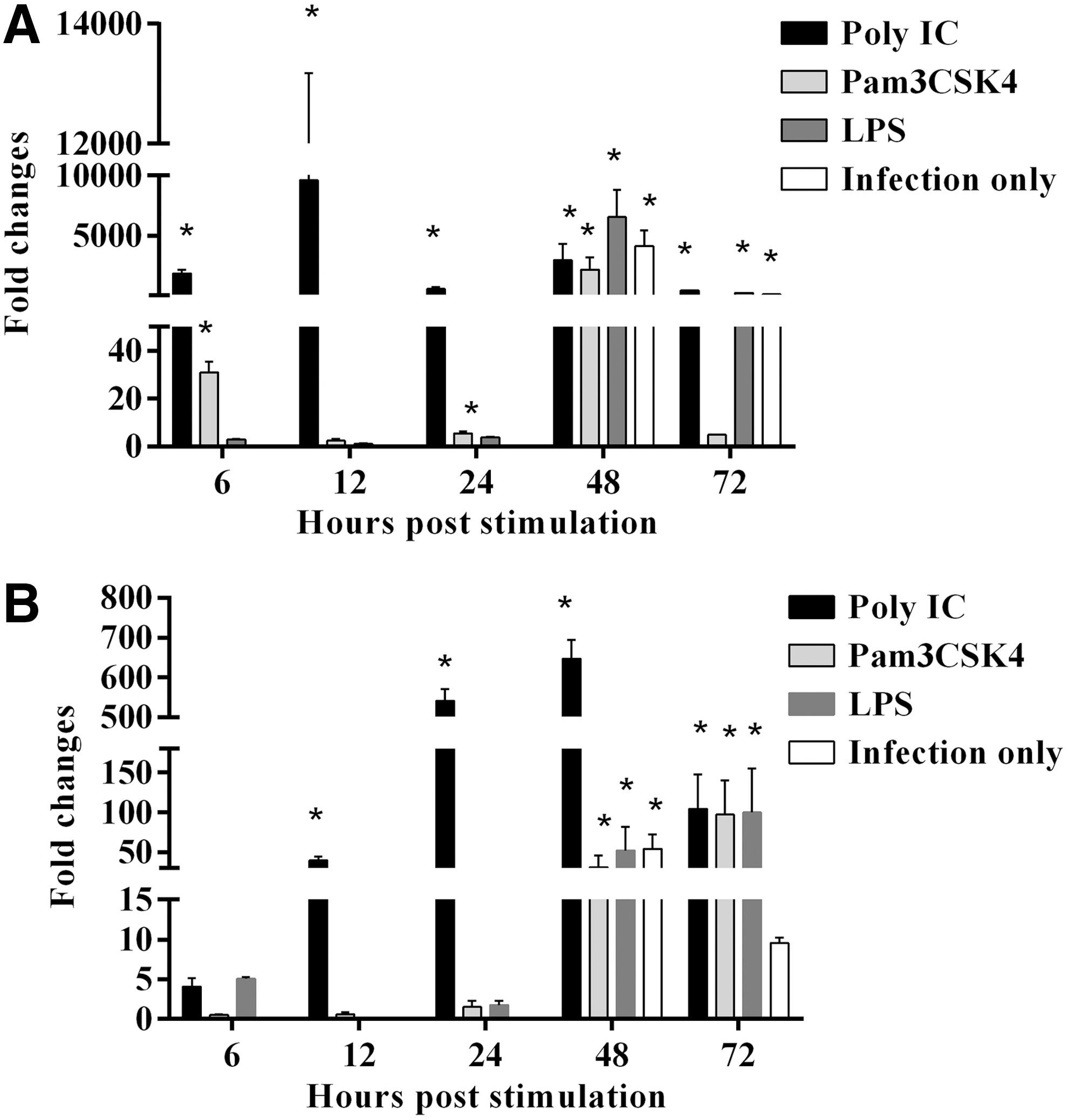
Relative expression of IFIT5 and Mx in TLR-Ls pretreated, MDV-infected CEFs. CEFs were pretreated with Pam3CSK4, Poly(IC), and LPS, and infected with EGFP MDV. There were MDV infection only (Untreated CEFs but infected) and medium control groups.
Discussion
In this study, initially the constitutive expression of TLRs, such as TLR 2, 3, 4, 5, 7, and 21 was investigated in CEFs. Next, various TLR-Ls, including Poly(IC), Pam3CSK4, LPS, and CpG ODN, were evaluated for their ability to inhibit MDV infection in CEFs. Following that, TLR-L treatments causing significant reduction of MDV infection were selected for further study, in which the expression of selected cytokine genes and ISGs was investigated.
We demonstrated that all TLRs examined in this study were expressed in CEFs. The relative expression of these TLRs in CEFs, from the highest to the lowest, was TLR2, TLR4, TLR21, TLR3, TLR7, and TLR5. In comparison to this observation, another study reported that high expression of TLR3 was followed by TLR7 and TLR4 in CEFs from MD-resistant and -susceptible chicken lines (15). In addition, the expression of TLR3, TLR4, and TLR7 in uninfected CEFs was higher in MD-resistant chicken lines than that of susceptible chicken lines (15). This difference suggests that expression pattern of TLRs in CEFs might vary depending on the genotype of chickens.
This study illustrated that it is possible to reduce MDV infection by pretreatment of cells with TLR-Ls. Although pretreatment of cells with all TLR-Ls caused significant reduction of MDV-infected CEFs, pretreatment with Poly(IC) had a more potent effect compared with other TLR-Ls. Similarly, in chickens, Poly(IC) treatment significantly reduced the oropharyngeal and cloacal shedding of avian influenza virus (30). MDV and herpes simplex viruses (HSV) are categorized under the subfamily Alphaherpesvirinae because of their structural similarities. In addition, MDV has some physiological properties comparable to HSV. Recent reports have also shown that Poly(IC) pretreatment could effectively reduce HSV type 1 and 2 (HSV1 and HSV2) infection in vitro (5,12). Additionally, strong antiviral effect of Poly(IC) against HSV2 was demonstrated when vaginal delivery of Poly(IC) completely inhibited the viral replication in mice (13).
The potent effect of Poly(IC) pretreatment was observed even with low level expression of TLR3 in CEFs. In this case, Poly(IC) might have induced the expression of additional TLR3 in CEFs leading to more effective recognition of this dsRNA mimic to trigger the downstream signaling pathway. Changes in the expression of TLR3 due to TLR-L treatment in CEFs were not examined in this study, however, other researchers have demonstrated the upregulation of TLR3 following Poly(IC) treatment (19,32) where the upregulation of TLR3 occurred due to the secretion of type 1 IFN in cell lines of chickens and mammals (19,38). It is well known that Poly(IC) recognition by TLR3 leads to the induction of type 1 IFN. Therefore, it is plausible that the positive feedback mechanism of type 1 IFN leads to the increased expression of TLR3 by Poly(IC) pretreatment, which might have contributed to the strong antiviral response against MDV infection in this study. Moreover, recognition of Poly(IC) is not solely dependent on TLR3. Other cytoplasmic receptor-like melanoma differentiation-associated gene 5 (MDA5) can act as a receptor for Poly(IC) (20,21). Since MDA5 is present in most cells including DF-1 cells (16), it is likely that MDA5 might have compensated for the low constitutive expression of TLR3 by CEFs.
The assessment of transcriptional profiles of cytokines and ISGs in this study revealed that all target genes were upregulated after MDV infection in all experimental groups. This raises the possibility that induction of these genes by TLR-Ls was responsible for the reduction in MDV replication in TLR-L pretreated cells. The significant upregulation of both IFN-β and IFN-α genes by Poly(IC) pretreatment occurred at early time points. In contrast, a study by Gill and colleagues (13) indicated that IFN-β, but not IFN-α, stimulated by Poly(IC) played a major role in protection against HSV2 infection in mice. In addition to type 1 IFN, IRF7, IFT5, and Mx genes were significantly upregulated by Poly(IC) pretreatment at early time points in this study. Type 1 IFN, induced by the TLR3 signaling cascade, can act on cells in an autocrine and/or paracrine fashion and induces the subsequent signaling events to activate ISGs.
The ISGs studied here, such as IFIT5 and Mx, act at different steps of viral replication and halt the production of new generation of viruses (35). Further, IRF7, which is an IFN regulator, can directly trigger the expression of certain ISGs like IFIT5, independent of IFN secretion (34,35). All these events might be attributed to the potent antiviral response conferred by Poly(IC) pretreatment.
In general, two scenarios for the reduction of MDV-infected cells can be envisaged. First, this result could be due to the interference in replication of MDV in infected CEFs. This could have happened due to the presence of several ISGs including IRF7 and IFIT5, which generate a hostile environment for virus replication. Second, even after successful replication in CEFs, reduction could be due to the ineffective spreading of MDV to adjacent cells, possibly due to the activation of an antiviral state in bystander cells by Poly(IC). Type 1 IFN secreted by infected cells may have acted on neighboring cells and induced the activation of ISGs to prevent viral entry. The mechanism of MDV entry into the cells has yet to be elucidated, however, similar to other viruses, Mx expression in CEFs might have contributed to resistance of the cells to virus infection (35). Hence, the antiviral state induced by Poly(IC) pretreatment before MDV infection may be the reason for the effective reduction in the MDV-infected CEFs by Poly(IC).
Pam3CSK4 and LPS failed to induce a functional antiviral state equal to that triggered by Poly(IC). Poly(IC) significantly upregulated all genes examined in this study, however, Pam3CSK4 significantly upregulated only IFN-β, IL-1β, and IFIT5 genes while LPS significantly upregulated only IL-1β and IRF7. This insufficient production of antiviral components for the preparation of CEFs by Pam3CSK4 and LPS might be the reason for the lower reduction of MDV-infected CEFs compared with Poly(IC) pretreatment.
Surprisingly, high dose of CpG and NCpG pretreatment significantly increased MDV-infected CEFs at 120 hps when compared to MDV infection only control. Similar to this observation, proviral activity of CpG was reported with human cytomegalovirus (HCMV) infection in neonatal human dermal fibroblasts when CpG was used as postviral treatment (17). The reason for this proviral activity is assumed to be the involvement of the PI3K/Akt pathway, which is activated in a TLR9-dependent manner (36) and HCMV uses the same pathway for its replication (18). However, since cellular activation and signaling pathways induced by MDV are not well studied, it is difficult to speculate the possible mechanism of enhancement of MDV infection in CpG treated cells.
Further, enhanced viral replication is not uncommon in cells treated with CpG. CpG pretreatment increased the replication of Friend retrovirus and led to erythroleukemia in resistant mice (25). In another study, splenocytes from human immunodeficiency virus (HIV) transgenic mice were stimulated ex vivo with CpG and enhanced HIV replication was observed (11). Furthermore, There have been reports of immune stimulatory activities of NCpG similar to CpG (22,40). The mechanisms of action of NCpG have not been well studied in chickens, but in mammals, it appears that NCpG can operate through TLR9 signaling or perhaps other unknown pathways.
In conclusion, TLR2, TLR3, and TLR4 ligands (Pam3CSK4, Poly(IC) and LPS respectively), significantly limited MDV infection in CEFs. This effect was mediated by upregulation of cytokines related to antiviral responses and ISGs. Additional studies are needed to explore the mechanisms by which PAMPs reduce MDV infection in CEFs with the aim of developing methods to control MDV replication in chicken. Moreover, further investigation of the effect of Poly(IC) on reducing MDV infection in chicken, particularly in feather follicular epithelial cells is necessary. This might reveal the possibility of interrupting transmission of MDV to susceptible chickens.
Footnotes
Acknowledgments
We would like to thank Ayumi Matsuyama for her help in preparing MDV stocks. We would like to extend our gratitude to Dr. Benedikt Kaufer (Freie Universität) for his permission to use the fluorescently tagged MDV. We also thank the Central Instrumentation facility at the Pathobiology Department of University of Guelph for the use of FACS-Canto II flow cytometer. Jegarubee Bavananthasivam is a recipient of an Ontario Trillium Scholarship. This work was funded by the Natural Sciences and Engineering Research Council of Canada, and the Ontario Ministry of Agriculture, Food and Rural Affairs.
Authors' Contributions
Conceived and designed the experiments: J.B., R.K., S.S. Performed the experiments: J.B., L.R. Analyzed the data: J.B., R.K. Contributed to the writing of the article: J.B., R.K., L.R., S.S.
Author Disclosure Statement
Authors declare no conflicts of interest.