
Abstract
The antiviral and immunomodulatory role of vitamin D has been shown in various viral infections. However, there is scanty literature available about the effect of vitamin D supplementation in herpes simplex virus-1 (HSV-1) infection. Therefore, the present study aimed to evaluate the role of two different forms of vitamin D: 25-hydroxyvitamin D3 (25D3) and 1,25-dihydroxyvitamin D3 (1,25D3) against HSV-1 in HeLa cells. The HeLa cells were supplemented with either 25D3 or 1,25D3 before HSV-1 infection and were studied after 6, 12, and 24 h postinfection (p.i.). The mRNA levels of toll-like receptors (TLRs), (2, 3, 4, 7, and 9), vitamin D signaling genes, and HSV-1 were studied using real-time PCR. The HSV-1 DNA load was estimated in culture supernatant. The supplementation of 25D3 and 1,25D3 significantly downregulated the mRNA levels of TLR2 (p < 0.0001) at 12 h p.i. The mRNA levels of TLR9 were found to be significantly downregulated in 1,25D3-supplemented cells at 12 h p.i. Furthermore, the significant downregulation was observed in HSV-1 titer in both 25D3- and 1,25D3-supplemented cells at 24 h p.i.(p < 0.0001). However, the effect of 25D3 supplementation persisted till 24 h p.i. with significant downregulation of TLR2 (p < 0.05) mRNA levels. The supplementation of both 25D3 and 1,25D3 before HSV-1 infection was found to downregulate the viral titer and TLR2 mRNA during the intial phase of infection. However, the effect of 25D3 supplementation was found to last for a longer duration compared with 1,25D3.
Introduction
H
Recently, the antiviral and immunomodulatory role of vitamin D3 has been shown in respiratory viruses, hepatitis C virus (HCV), and dengue virus (2,7,14,15,21). Vitamin D3 can be obtained from 7-dehydrocholestrol in the skin and also from the diet. The vitamin D3 gets hydroxylated to 25-hydroxyvitamin D3 (25D3) by 25-hydroxylase (cytochrome P450 family 2 subfamily R member 1[CYP2R1]). Further hydroxylation by 1,α-hydroxylase (cytochrome P450 family 27 subfamily B member 1[CYP27B1]) converts 25D3 to 1,25-dihydroxyvitamin D3 (1,25D3). Both 25D3 and 1,25D3 are the agonists of vitamin D receptor (VDR) and could independently regulate the expression of various genes (12). Previous study on HSV-1-induced Behcet's disease has shown the downregulation of TLR2 and TLR4 on macrophage supplementation with 1,25D3 (3). However, the direct effect of vitamin D supplementation against HSV-1 infection in cell lines has not been studied earlier. Hence, the present study aimed to evaluate the role of 1,25D3 and 25D3 against HSV-1 infection in the in vitro system.
Materials and Methods
The HeLa cells were maintained in the department using Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS). The HSV-1 (KOS) was obtained from the National Institute of Virology, Pune, and propagated in Vero cells. The cytotoxicity of 25D3 (10 nM to 100 μM) and 1,25D3 (1 nM to 10 μM) was checked in HeLa cells, cultured in 96-well plate, using MTT (Sigma).
The in vitro testing of 25D3 (Enzo) and 1,25D3 (Sigma) using 0.1% absolute ethanol (Merk) as vehicle was performed in HSV-1-infected HeLa cells. The HeLa cells (105 cells/mL) were cultured for 24 h in 12-well culture plates (Corning, Sigma). The supplementation of 25D3 and 1,25D3 till 10 μM and 1 μM concentration to HeLa cells did not affect the viability of the respective cells (Supplementary Fig. S1; Supplementary Data are available online at
The RNA was isolated from cells at 6, 12, and 24 h postinfection (p.i.) and harvested using 1 mL of TRIzol (Ambion). The RNA was converted to cDNA using the High Capacity cDNA Synthesis Kit (Applied Biosystems) as per the manufacturer's instructions. The real-time PCRs were performed for TLRs (2,3,4,7,9), VDR, CYP27B1, cytochrome P450 family 24 subfamily A member 1 (CYP24A1), and HSV-1 (Supplementary Table S1) using 1X Maxima SYBR Green master mix (Thermo Fisher) and 250 nM of primers. The GAPDH was used as an endogenous control. The HSV-1 mRNA expression was calculated w.r.t. (with respect to) nonsupplemented HSV-1-infected cells (control or calibrator). The relative fold change expression was calculated using the 2−ΔΔCt method. The viral load was calculated from culture supernatants at 24 h p.i. because the lytic life cycle of HSV-1 generally takes around 18 h (8). The in-house TaqMan real time was performed targeting the glycoprotein D of the HSV-1 as described previously (9).
The results are expressed as mean ± SEM. The significance between groups was checked by using one-way analysis of variance (ANOVA) with Tukey's test using GraphPad Prism 6 (GraphPad Software). The p value <0.05 was taken as significant.
Results
The HeLa cells were supplemented with 25D3 and 1,25D3 before HSV-1 inoculation and the relative expression of VDR, CYP27B1, CYP24A1 was studied w.r.t. control cells at 6, 12, and 24 h HSV-1 p.i. (Supplemantary Fig. S2). The CYP24A1 was taken as the positive control for vitamin D signaling. The 25D3 and 1,25D3 supplementation significantly upregulated mRNA levels of CYP24A1 in controls and in HSV-1-infected cells.
The HSV-1 infection resulted in the significant upregulation of TLR2 throughout the 24-h period, while the mRNA levels of TLR 4 and 9 were significantly upregulated only during the initial 12 h p.i. The 25D3 supplementation resulted in significant downregulation of TLR2 mRNA levels in HSV-1-infected cells at 12 h p.i. 71.7 ± 11.5 vs. 16.2 ± 2.2 (p < 0.0001). The downregulation of TLR2 mRNA levels in HSV-1-infected 25D3 supplementation cells was seen to persist even at 24 h p.i. (11.6 ± 2.02 vs. 4.7 ± 0.81, p < 0.05). The mRNA levels of TLR2 were also found to be downregulated in HSV-1-infected 1,25D3 supplemented cells at 12 h p.i. 71.7 ± 11.5 vs. 17.9 ± 2.9 (p < 0.0001). Furthermore, the 1,25D3 supplementation also showed significant downregulation of TLR9 mRNA levels in HSV-1-infected cells at 12 h p.i. (3.5 ± 0.47 vs. 1.6 ± 0.12, p < 0.01) (Fig. 1).
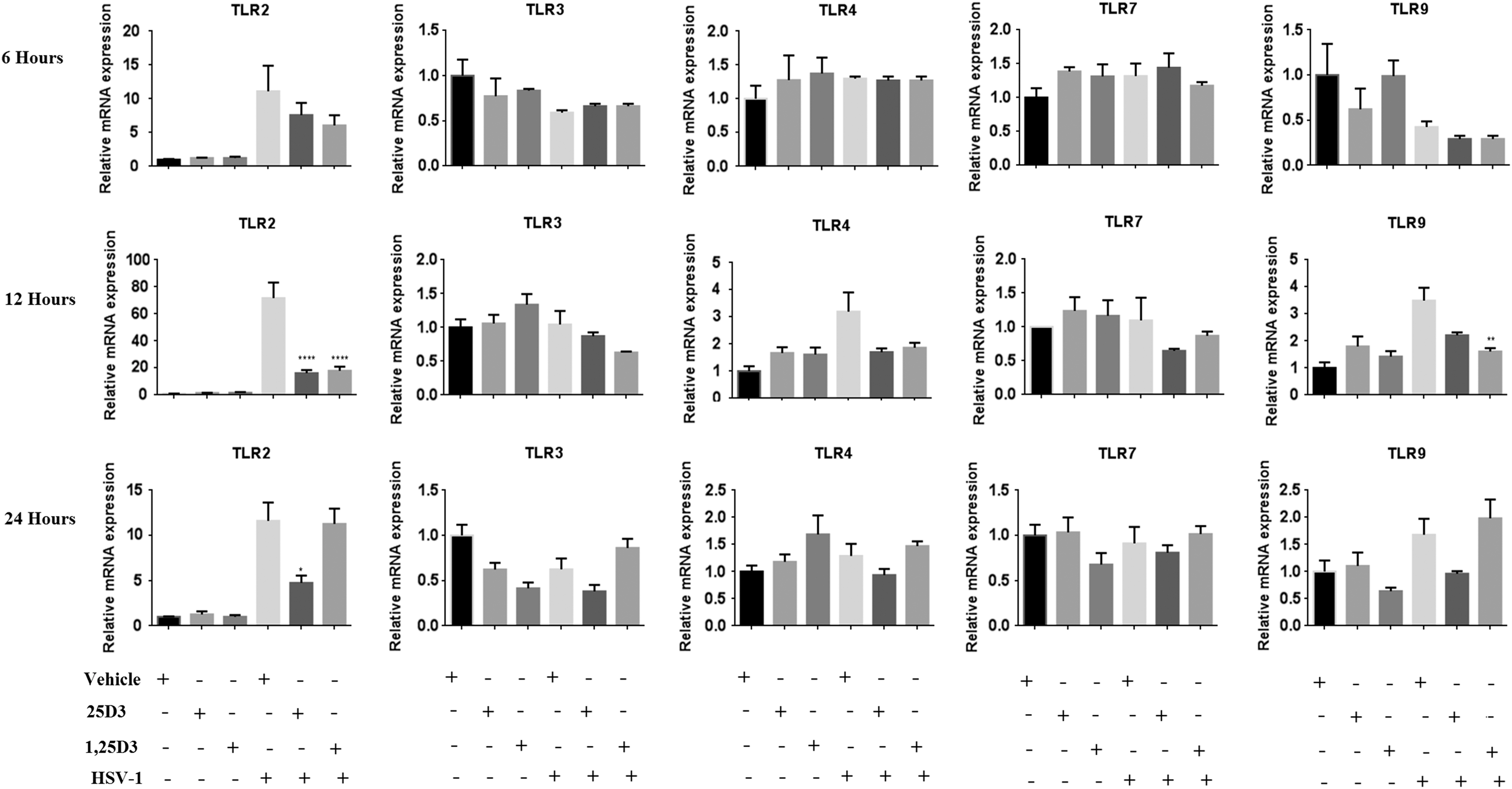
Relative mRNA expression of TLRs (2, 3, 4, 7, and 9) w.r.t. controls at 6, 12, and 24 h after HSV-1 infection in nonsupplemented and vitamin D-supplemented (25D3 and 1,25D3) HeLa cells. Data represented as mean ± SEM (*p < 0.05, **p < 0.01, ****p < 0.0001). HSV, herpes simplex virus; SEM, standard error of mean; TLRs, toll-like receptors; w.r.t, with respect to.
The 25D3 and 1,25D3 supplementation in HSV-1-infected HeLa cells was found to significantly regulate the viral titer and HSV-1 mRNA levels (Fig. 2). The HSV-1DNA titer was found to significantly decrease in both 25D3 (678,338.3 ± 30,996.3 vs. 190,400.1 ± 15,771.5, p < 0.0001)- and 1,25D3 (678,338.3 ± 30,996.3 vs. 211,634.1 ± 4,786.1, p < 0.0001)-supplemented HeLa cells after 24 h p.i. Furthermore, the HSV-1 mRNA levels were found to be significantly downregulated in the 25D3-supplemented group (p < 0.01).
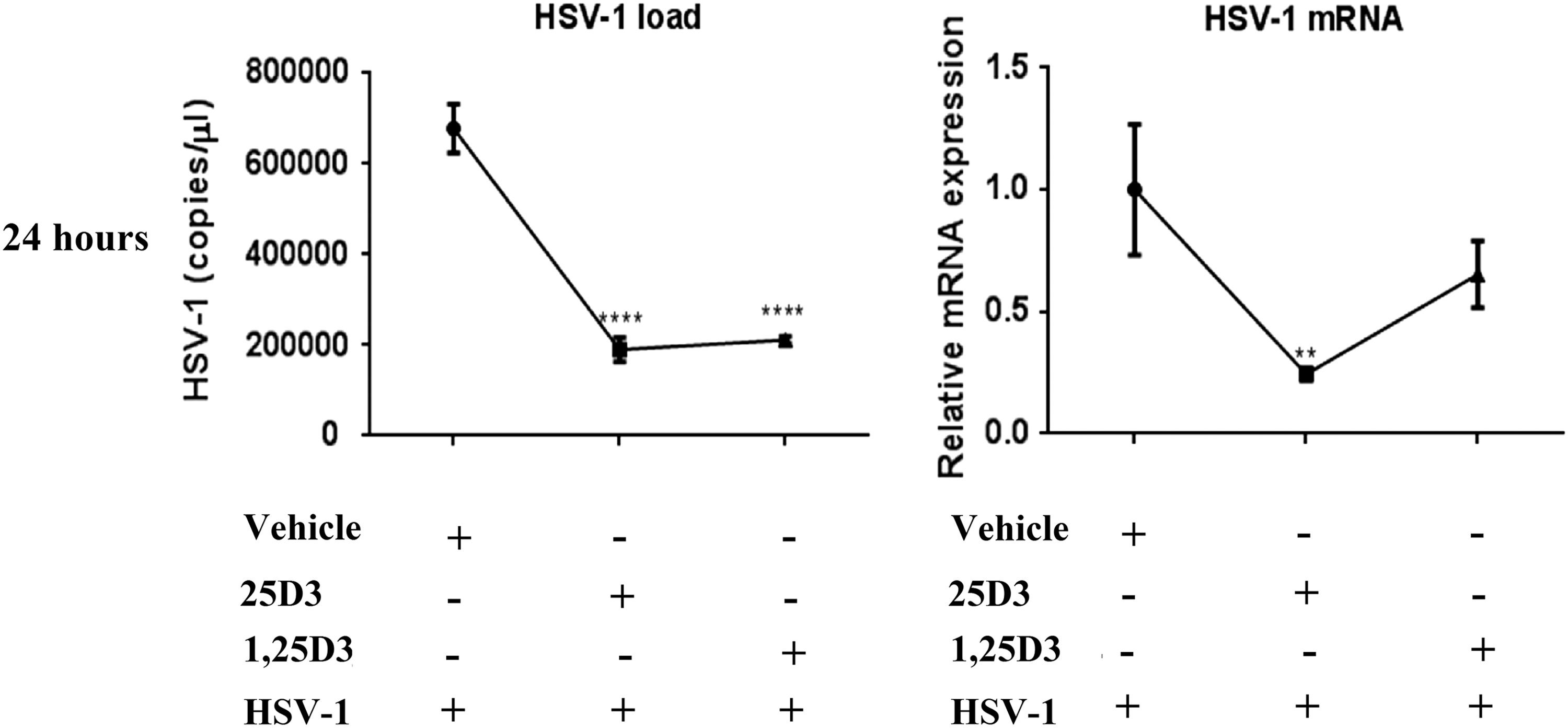
HSV-1 load in culture supernatant and mRNA levels in HSV-1 HeLa cells at 24 h after HSV-1 infection in nonsupplemented and vitamin D supplemented (25D3 and 1,25D3). Data represented as mean ± SEM (**p < 0.01, ****p < 0.0001).
Discussion
The HSV-1 infection to host cells results in the activation of TLRs and thereby initiates an immune response against viral infection either by regulating NFκB or interferon regulatory factors (IRFs) (13). In the present study, a significant increase in the expression of TLR2, with minimal expression of TLR4 and TLR9, was observed in a time-dependent manner, which may be attributed to various factors such as cell tropism, synergy between the TLRs, and viral titer (10). However, the vitamin D supplementation to the HeLa cells before the HSV-1 infection significantly downregulated the TLR2 as well as HSV-1 mRNA expression.
In the present study after HSV-1 infection of HeLa cell line, the mRNA level of TLR2 was found to be elevated throughout the study. Various studies have shown the role of TLR2 in inducing the expression of proinflammatory cytokines against HSV-1 infection (1,19). On the contrary, a few studies have shown that the upregulation of TLRs may have a beneficial role for the host. Sørensen et al. have shown a synergistic effect of TLR2 along with TLR9 in controlling HSV-2 infection (16). However, Lima et al. have shown that the TLR9 plays a more important role in increasing mice survival compared with TLR2 in HSV-1-infected mice (11). In the present study, both 1,25D3 and 25D3 significantly downregulated the mRNA levels of TLR2 in HeLa cells at 12 h p.i. Also at 12 h p.i., downregulation of TLR9 was observed in 1,25D3-supplemented cells. Similar results have been reported by Dickie et al. (2010) in human monocytes by TLR9 agonists (4). However, the effect of 25D3 supplementation on TLR2 downregulation lasted for 24 h HSV-1 p.i. Choi et al. have shown that the supplementation of 1,25D3 to the macrophages isolated from HSV-1-induced BD-like mice led to a significant reduction in the expression of TLR2 and TLR4 (3). In another study, 1,25D3 supplementation in dengue-infected U937 and Huh-7 cells was found to downregulate the expression of TLR2 and TLR4 (15). In the present study, low expression level of TLR4 was observed, which did not significantly change with vitamin D supplementation.
The antiviral effect of vitamin D3 has been shown in various viral infections such as HCV, rhinovirus, and dengue virus (2,14,15,18). In the present study, significant lower levels of HSV-1 mRNA and DNA were observed in 25D3-supplemented cells. However, the HSV-1 DNA levels were significantly lower in 1,25D3-supplemented cells, but HSV-1 mRNA levels were nonsignificant.
In the present study, the effect of 25D3 persisted for a longer period, that is, 24 h p.i. compared with 1,25D3. The difference in these results may be attributed to the following: (a) the use of different concentrations of both forms of vitamin D in the present study. The studies on rhinovirus have found that 1,25D3 is effective in regulating proinflammatory cytokines at 10 μM and 100 nM concentrations, while the downregulation of rhinovirus mRNA levels was observed only at 100 nM of 1,25D3 (2). In the present study, 1,25D3 at 100 nM and 25D3 at 1 μM were shown to have both immunomodulatory and antiviral activities in a time-dependent manner. (b) It is well known that 1,25D3 is a direct agonist of VDR and regulates the expression of various genes. Recently, it has been shown that 25D3 can also directly interact with VDR without being converted to 1,25D3 (12). Furthermore, the different binding affinities of 25D3 and 1,25D3 could result in different findings (12). Matsumura et al. have also shown that the inhibition of HCV was observed by using 1 μM concentration of 25D3, while no inhibition of HCV was observed by using either vitamin D3 or 1,25D3 (14). The findings of the present study are in agreement with the previous authors who have suggested the involvement of different signaling mechanisms of 25D3 independent of 1,25D3 (14).
The supplementation of 25D3 and 1,25D3 before HSV-1 infection was found to be effective in regulating TLR2 and HSV-1 titer and thereby may be helpful in regulating the virus-induced pathogenesis. Moreover, the 25D3 supplementation was found to be more effective and lasted for a longer time compared with 1,25D3. Hence, based on the current findings, the supplementation of 25D3 may be effective in modulating HSV-1-induced inflammatory pathways.
Footnotes
Acknowledgment
We thank the Indian Counsil of Medical Research, New Delhi, for providing Senior Research Fellowship to A.K.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.