
Abstract
Accumulating evidence demonstrates that CD8+CD28− regulatory T cells increase in chronic viral infection as well as tumorigenesis. However, it is still not clear about their characteristics in hepatitis B virus (HBV) infection. In addition, it is not understood whether this regulatory immune subset is distinct from CD4+CD25high regulatory T cells in the aspect of impact on or relationship to the progression of HBV infection. Hence, we investigated their dynamics and compared their correlations with clinical parameters in the chronic and advanced phases of HBV infection. The data showed that compared with healthy controls, the frequencies of CD28+CD8− and CD4+CD25high T cells increased in both chronic and advanced phases, while there is no significant difference between the two case groups. Interestingly, we found that in chronic phase, the frequency of CD8+CD28− subset was negatively correlated with the levels of alanine aminotransaminase (ALT) and aspartate aminotransferase (AST), respectively, and did not present association with HBV DNA load, whereas that of CD4+CD25high T cells was positively correlated with HBV DNA load and the levels of ALT and AST, respectively. Amazingly, in advanced phase, the frequency of CD4+CD25high T cells was negatively correlated with HBV DNA load and the levels of ALT, respectively, while there is no significant correlation between the frequency of CD8+CD28− subset and those clinical parameters. Thereby, our findings demonstrated that CD28+CD8− and CD4+CD25high regulatory T cells might exert distinct effect on modulating antiviral immune responses and mitigate immunomediated liver damage in different phases of HBV infection, which represent potential prognostic markers and therapeutic targets for HBV-infected patients based on further exploration of detailed mechanism.
Introduction
C
HBV-specific T lymphocytes play a critical role in eliminating viruses (9,13,23). However, they are usually suppressed with a status of functional exhaustion by multiple aspects of inhibitory factors (10,30,33). For example, immune checkpoints, such as programmed cell death 1 (PD-1), lymphocyte-activation gene-3 (LAG-3), and T cell immunoglobulin mucin-3 (Tim-3), when upregulated on T cells and engaged with their ligands, inhibit T cell antiviral function and contribute to progression of HBV infection (14,18 –20). At the same time, upregulation of immune checkpoints on T cells is associated with liver injury, in addition to HBV amplification. For instance, the increase of PD-1 expression on peripheral T cells and PD-L1 expression on CD19+ B cells in CHB patients is in consistence with liver function injury as well as HBV amplification (11,32).
Moreover, regulatory immunocytes such as CD4+CD25high regulatory T cells play a vital important role in suppressing T lymphocyte function in HBV infection and relate to liver injury in CHB patients (6,21,31). In our and another previous study, CD4+CD25high regulatory T cells were revealed to increase in HBV infection and correlate with the elevation of alanine aminotransaminase (ALT) and aspartate aminotransferase (AST) levels (7). Nevertheless, the detailed dynamics of CD4+CD25high regulatory T cells is still not fully understood during HBV infection from chronic to advanced stages.
CD8+CD28− T cells were initially considered an age-related increased population (26). Recently, CD8+CD28− T cells have been described as a subset of suppressor cells within the CD8+ T cell population (5). CD8+CD28− T cells can directly inhibit T lymphocyte activation and proliferation, downregulate effector cytokines secreted from activated T cells, and induce T cell apoptosis (5,16). In addition, CD8+CD28− T cells can also exert their extraordinary effect on regulating T cell responses by secreting soluble factors and expressing membrane ligands, such as TGF-β, IL-10, PD-L1, and FasL (3).
Although CD8+CD28− T cells have a protective role in preventing experimental inflammatory bowel disease and alleviating acute allograft rejection (26,28,29), accumulating evidence demonstrates that an increase of this subset is associated with tumor progression and chronic viral infections such as hepatitis C virus (HCV) or human immunodeficiency virus (HIV) (8,26). However, it is not clarified how the frequency of CD8+CD28− subset varies and whether it is correlated with disease progression in HBV infection. Furthermore, it is not unknown whether CD8+CD28− subset is distinct from CD4+CD25high regulatory T cells in aspects of their association with the progression of HBV infection.
Hence, in this study, we aim to explore the dynamics and clinical significance of CD8+CD28− and CD4+CD25high regulatory T cells since their correlations with HBV infection from chronic to advanced phases have not been or have not been fully described. At the same time, we sought to clarify whether there is a difference between these two kinds of regulatory T cells associated with the progression of HBV infection.
Materials and Methods
Study participants
The enrolled samples in this study included healthy controls (HC) (n = 21) and patients with HBV infection in chronic phase (CHB, n = 34) and advanced phase (n = 24, including LC and HCC patients). HC were negative with serum HBV antigens and antibodies, and did not show infection of HCV, hepatitis D virus (HDV), and HIV-1 and other autoimmune diseases. CHB patients presented positive HBsAg and persistent elevation of serum ALT levels for 6 months. At the same time, CHB patients were identified with detectable HBV DNA. The diagnosis of LC was based on computed tomography (CT), acquiring images with an evidence of cirrhosis. LC patients were also serum HBsAg positive carrying HBV DNA. HCC patients with positive serum HBsAg and elevated levels of alpha-fetoprotein had multiple arterial vascular nodules by CT imaging examination.
All the patients were negative with antibodies to HCV, HDV, and HIV-1, and not combined with tumor and other symptoms of liver damage such as autoimmune disease, alcoholic liver disease, and drug-induced hepatitis. None of the patients received antiviral therapy and/or immune therapy before they were enrolled (clinical baseline were listed in Table 1). All the patients and HC were Chinese. The study was conducted at the Affiliated Infectious Hospital of Soochow University (The Fifth Hospital of Suzhou City) and approved by the ethics committee of the hospital (No. 2016–003).
Data are presented as number or mean ± standard error of the mean.
ALB, albumin; ALT, alanine aminotransaminase; AST, aspartate aminotransferase; HBeAg, hepatitis B virus e antigen; HBV-DNA, hepatitis B virus-DNA; HC, healthy controls; NA, not applicable; TBIL, total bilirubin.
Serological assays
HBsAg, HBeAg, HBsAb, HBcAb, and HBeAb were detected by enzyme-linked immunosorbent assay with commercially available kits (Abbott Ireland, Sligo, Ireland). Serum HBV-DNA level was measured by fluorescent quantitative PCR with commercially available kits (Shanghai Shenyou Technology Co. Ltd., Shanghai, China). Positive samples with HBV had a viral load >5 × 102 copies/mL. Biochemical analysis was applied to measure ALT, AST, and other liver function.
Flow cytometry
Heparin-anticoagulated blood was collected from healthy donors and patients with HBV infection. All antibodies were purchased from BD Pharmingen (San Diego, CA). Staining for the surface markers was performed according to the manufacturers' instructions as follows: briefly, 100 μL blood from study samples for each test was incubated with anti human CD3-APC, CD4-PerCP5.5, and CD25-FITC (or Mouse IgG-FITC), or CD3-APC, CD8-PerCP5.5, and CD28-PE for 15 min. Lysed blood was prepared by red blood cell (RBC) lysis buffer and washed twice in phosphate-buffered saline. Eventually, cells were suspended and determined immediately.
For determining FoxP3 expression in CD4+CD25+ regulatory T cells, a Human Regulatory T Cell Whole Blood Staining Kit purchased from eBioscience (San Diego, CA) was adopted according to the standard procedure. Briefly, for each test, 100 μL of fresh heparinized whole blood from patients or healthy donors was incubated with mouse antihuman CD4-FITC, CD25-PE/Cy7, for 15 min, then lysed with RBC lysis buffer, and subsequently dealt with fixation/permeabilization solution after washing and centrifuging. Furthermore, mouse anti human Foxp3-Alexa Flour 647 or mouse IgG1-Alexa Flour 647 was added and incubated at 4°C for 30 min in the dark. Eventually, cells were suspended and determined immediately. All samples were determined by BD FACSAria (San Jose, CA) supported by DB FACSDiva software (San Jose, CA). Data were analyzed using FlowJo (San Carlos, CA). Then the samples were measured by using the instrument fluorescence-activated cell sorting (FACS). FACS data were analyzed using FlowJo software (San Carlos, CA). Monoclonal antibodies were purchased from BD PharMingen (BD Biosciences, San Jose, CA).
Statistical analysis
Statistical analyses were performed using GraphPad Prism 6.0 software showing mean ± standard error of the mean. Unpaired t test was used to assess the difference between different groups. The Spearman correlation coefficient was calculated to assess the frequencies of CD4+CD25high and CD28+CD8− subsets and HBV viral load, ALT, or AST level. A two-tailed p < 0.05 was considered statistically significant.
Results
Investigation of the frequency of CD8+CD28− regulatory T subsets from patients with HBV infection
The participants recruited in this study contain HC (n = 21) and patients with HBV infection in chronic phase (n = 34) and advanced phase (n = 24). The clinical parameters of these study subjects are provided in Table 1, including age, gender, Child-Pugh score, BCLC stage, HBeAg sera status, serum HBV DNA concentrations, serum ALT and AST levels, albumin, and total bilirubin.
Since it is still not clear whether the frequency of CD8+CD28− regulatory T cells varies in and correlates with clinical parameters during the progression of HBV infection from chronic to advanced phases, we examined this subset within CD8+ T cell population by using FACS. The gating strategies for determination and analysis of this subset are shown in Figure 1A. Briefly, CD8+ T cells was first gated from CD3+ population and then the frequency of CD8+CD28− subset occupied in CD8+ population was figured out. The data showed that compared with the HC group, the frequency of CD8+ T cell increased in groups with HBV infection (p < 0.05) (Fig. 1B), but did not present a change between the two groups, respectively, in chronic and advanced phases of HBV infection (Fig. 1B).
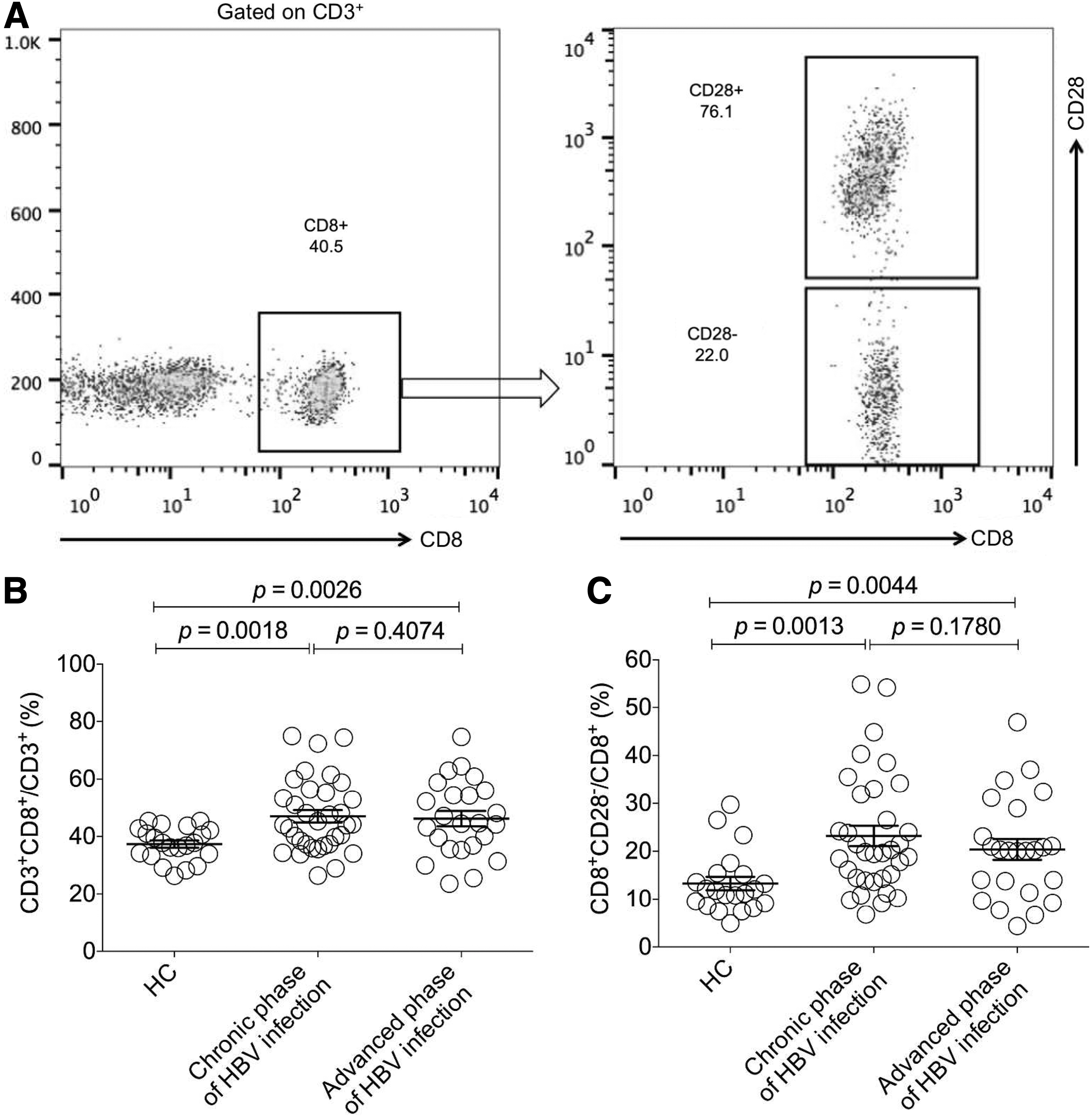
Determination and analysis of the frequency of CD8+CD28− regulatory T cells in chronic and advanced phases of HBV-infected patients.
Further analysis demonstrated that there was a significant increase of the frequency of CD8+CD28− regulatory T cells in both case groups compared with the HC group (p < 0.05) (Fig. 1C), suggesting that this regulatory T subset also increases in HBV infection with similarities to that in HCV and HIV infection (8,26). However, no significant difference between the two case groups was found (Fig. 1C).
Investigation of the frequency of CD4+CD25high regulatory T subsets from patients with HBV infection
CD4+CD25+ regulatory T cells have been well reported as contributing to the suppression of anti-HBV immune responses and associating with clinical parameters in chronic phase of HBV infection (31). However, the dynamics of frequency of CD4+CD25+ regulatory T cells during HBV infection from chronic to advanced phase is still not systematically described. In this study, we sought to investigate its variation along with the progression of HBV infection.
The population with a CD4+CD25high phenotype was adopted to characterize CD4+CD25+ regulatory T cells according to previous reports (19,31). The gating strategies for CD4+CD25high population are shown in Figure 2A. First, the CD4+CD25+ population was analyzed and the results demonstrated that compared with HC, the frequency of CD4+CD25+ population within CD4+ T cells from the case groups in chronic and advanced phases was, respectively, elevated (p < 0.05) (Fig. 2B), while no significant difference between the two case groups appeared. Then, we investigated the CD4+CD25high population and found that there was a significant increase of the frequency of CD4+CD25high T cells from patients in both chronic and advanced phases compared with the HC group (p < 0.05) (Fig. 2C), whereas there was no significant difference between the two case groups (Fig. 2C).
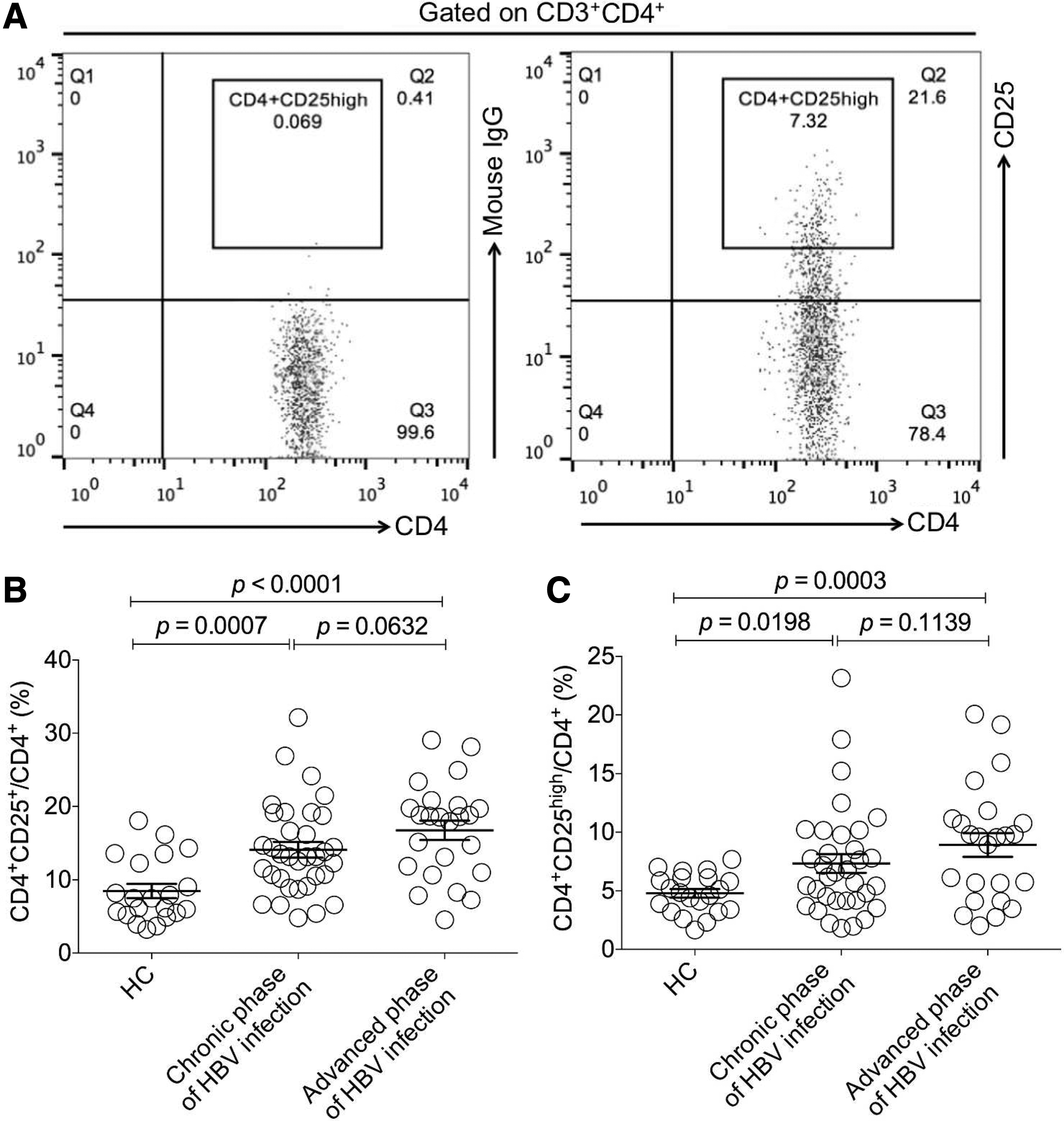
Determination and analysis of the frequency of CD4+CD25high regulatory T cells in chronic and advanced phases of HBV-infected patients.
Since the expression of Foxp3 is a more appropriate marker for CD4+CD25+ regulatory T cells, we further validated Foxp3 expression in CD4+CD25+ regulatory T cells and investigated whether the frequency of CD4+CD25high population was consistent with that of CD4+CD25+Foxp3+ regulatory T cells by using the method of FACS. The samples included HC (n = 8) and patients with HBV infection in chronic phase (n = 10) and advanced phases (n = 6). The gating strategies for populations with a phenotype of CD4+CD25+Foxp3+ or CD4+CD25high are shown in Figure 3A. As expected, a subpopulation within CD4+CD25+ T cells expressed Foxp3 and the frequency of CD4+CD25high population was basically in line with that of CD4+CD25+Foxp3+ subset (Fig. 3B), implying that CD4+CD25high can be regarded as a reliable indicator to reflect CD4+CD25+Foxp3+ regulatory T cells in our study.
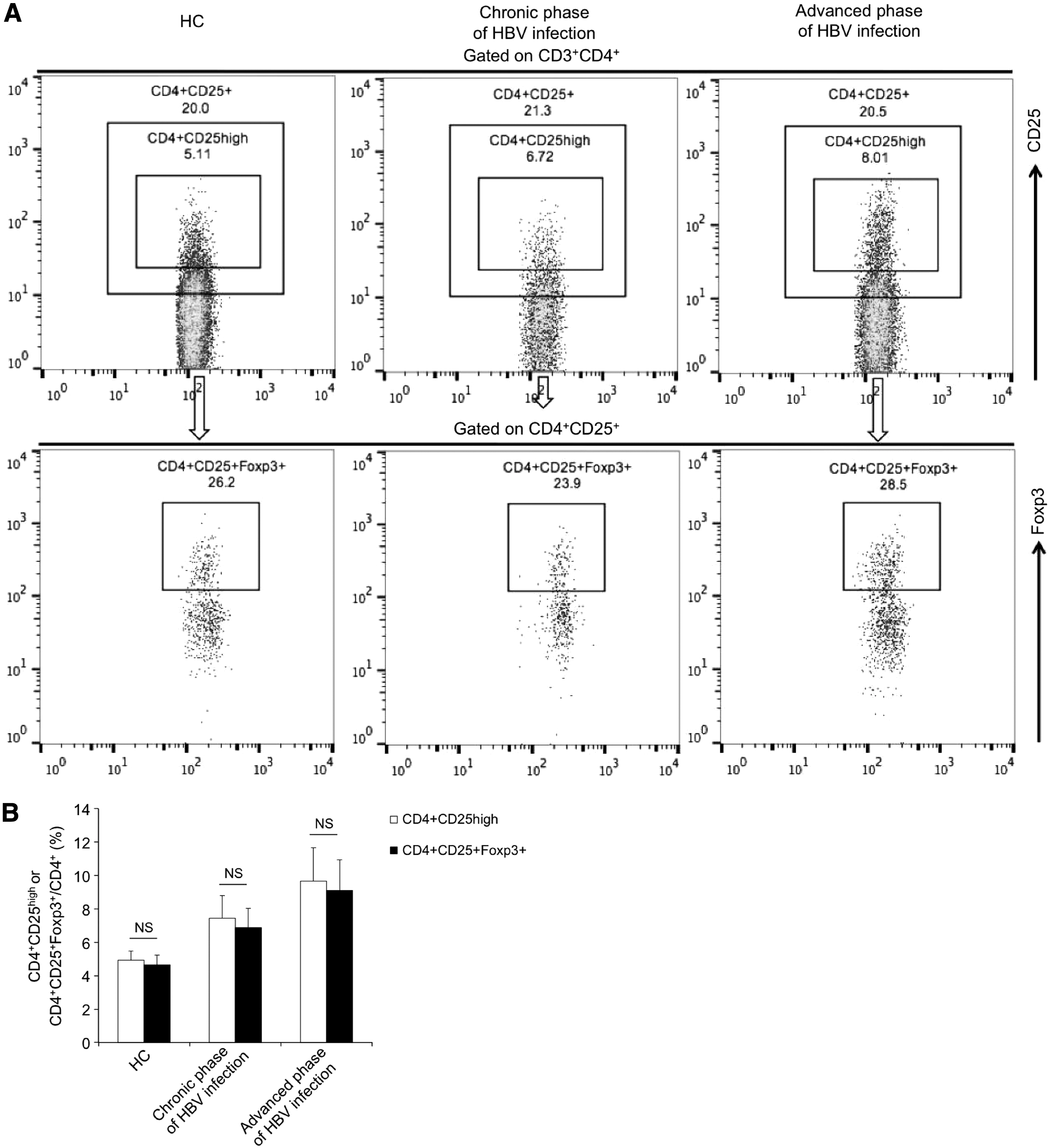
Determination of Foxp3 expression in CD4+CD25+ T cells and comparison of the frequencies of CD4+CD25high and CD4+CD25+Foxp3+ T cells in HC (n = 8), chronic (n = 10), and advanced phases (n = 6) of HBV-infected patients.
Comparison of the correlations of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells with HBV DNA load, and ALT and AST levels from patients in chronic phase of HBV infection
The increase of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells probably hampers effective anti-HBV immune responses, resulting in viral amplification and the exacerbation of infection during CHB. However, it is unknown whether the impact of CD8+CD28− subset on the development of HBV infection is distinct from that of CD4+CD25high regulatory T cells. Therefore, we analyzed the correlation of frequencies of these two subsets with clinical parameters, including HBV DNA load and levels of ALT and AST, respectively.
Amazingly, the results showed that in the chronic phase of HBV infection, the frequency of CD8+CD28− subset was not associated with HBV DNA load (r = −0.0368, p = 0.4181) (Fig. 4A), and negatively correlated with the levels of ALT (r = −0.3789, p = 0.0136) (Fig. 4B) and AST (r = −0.3800, p = 0.0133) (Fig. 4C), respectively, while that of CD4+CD25high regulatory T cells was positively correlated with HBV DNA load (r = 0.6303, p < 0.0001) (Fig. 4D), and ALT (r = 0.5253, p = 0.0007) (Fig. 4E) and AST (r = 0.3808, p = 0.0132) (Fig. 4F) levels, respectively.
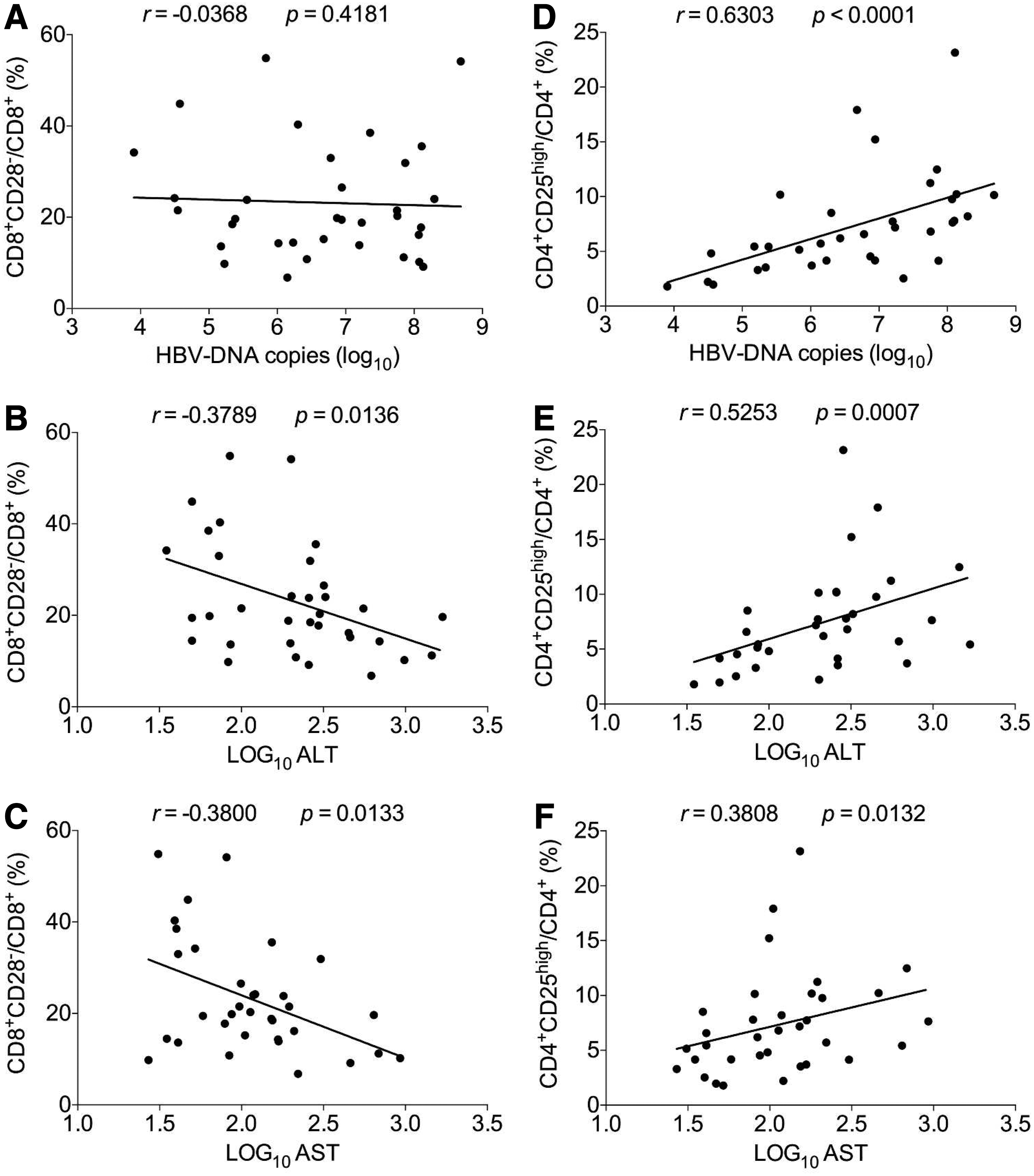
Analysis of the correlations of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells with HBV DNA load, and ALT and AST levels from patients in chronic phase of HBV infection.
Comparison of the correlations of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells with HBV DNA load, and ALT and AST levels from patients in advanced phase of HBV infection
Persistent CHB, generally causing immunomediated liver injury, probably develops into an advanced phase characterized by LC as well as HCC. In this phase, anti-HBV immune function is severely suppressed, the composition of immunocytes tends to be chaotic, and the impact of immune subsets on HBV amplification as well as liver injury probably varied. Thereby, we further investigated the correlations of frequencies of CD8+CD28− and CD4+CD25high regulatory cells with clinical parameters and made a comparison. Interestingly, in the advanced phase of HBV infection, the frequency of CD8+CD28− subset was not found to correlate with any clinical parameter determined in our study (Fig. 5A–C), which is basically different from what happened in the chronic phase of HBV infection.
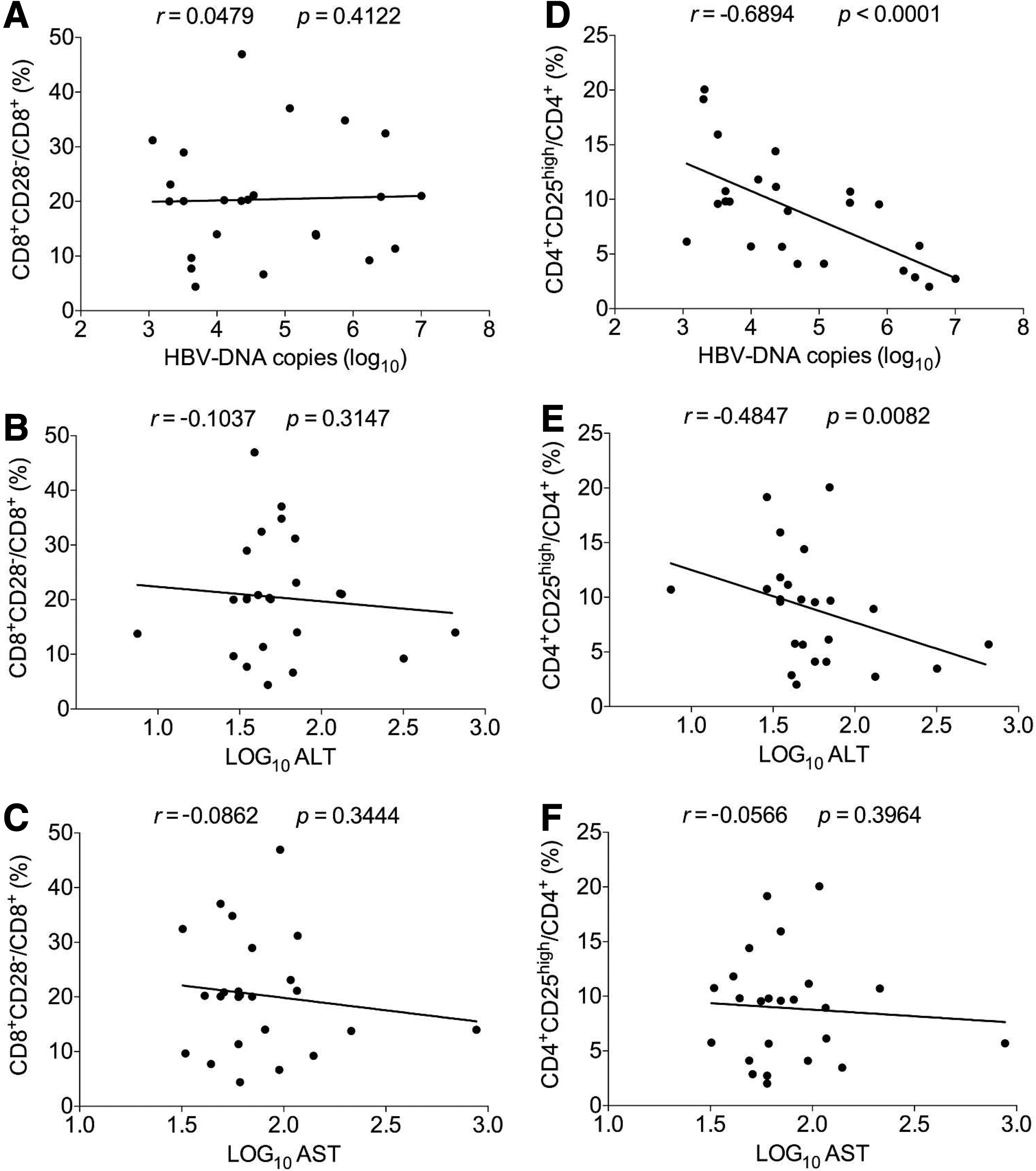
Analysis of the correlations of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells with HBV DNA load, and ALT and AST levels from patients in advanced phase of HBV infection.
However, in this phase, the frequency of CD4+CD25high regulatory T cells was negatively associated with HBV DNA load (r = −0.6894, p < 0.0001) (Fig. 5D) and ALT levels (r = −0.4847, p = 0.0082) (Fig. 5E), respectively, and was not related to AST levels (Fig. 5F), which were almost the opposite of what presented in the chronic phase of HBV infection.
Investigation of the correlation of frequency of CD8+CD28+ T cells with HBV DNA load, and ALT and AST levels from patients with HBV infection
Also, we analyzed the clinical significance of CD8+CD28+ T cells from patients with HBV infection. First, we found that compared with the HC group, the frequency of CD8+CD28− T cells from the two case groups significantly decreased (p < 0.05) (Fig. 6A), while there was no significant difference between the two case groups (Fig. 6A). Further investigation demonstrated that in the chronic phase of HBV infection, the frequency of CD8+CD28+ subset was not associated with HBV DNA load (r = 0.0372, p = 0.4190) (Fig. 6B), and positively correlated with the levels of ALT (r = 0.3796, p = 0.0142) (Fig. 6C) and AST (r = 0.3825, p = 0.0135) (Fig. 6D), respectively, while in the advanced phase, it was not associated with any clinical parameter determined in our study (Fig. 6E–G).
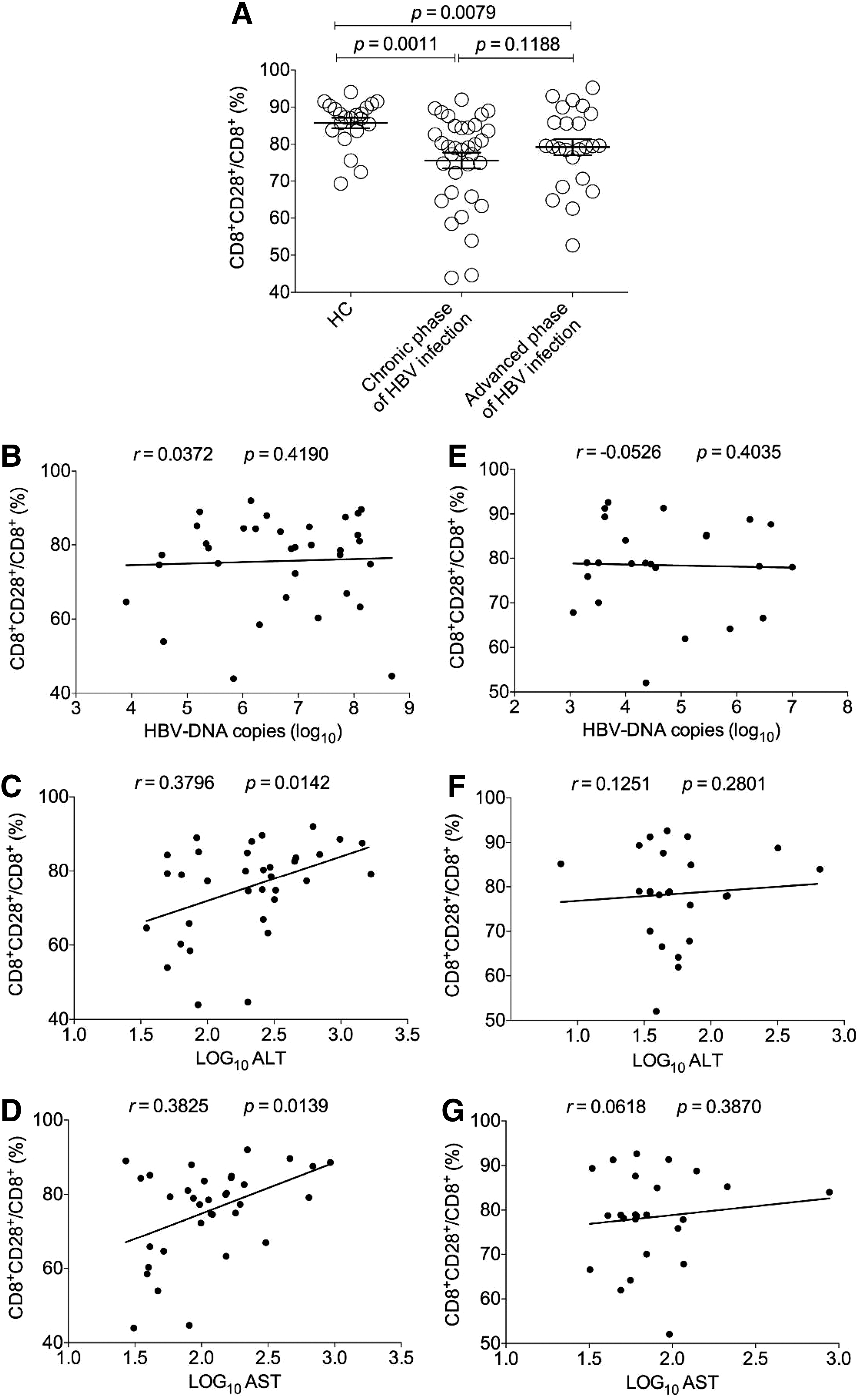
Analysis of the correlations of frequency of CD8+CD28+ T cells with HBV DNA load, and ALT and AST levels from patients with HBV infection.
Investigation of frequencies of CD8+CD28− and CD4+CD25high regulatory T cells in relation to HBeAg
Moreover, we explored whether there was a relationship between HBeAg and the frequencies of CD8+CD28− and CD4+CD25high regulatory T cells from patients with chronic infection of HBV. The results demonstrated that no significant differences were detected in the frequencies of both CD8+CD28+ and CD4+CD25high regulatory T cells between the HBeAg-negative and HBeAg-positive patients (Fig. 7A, B).
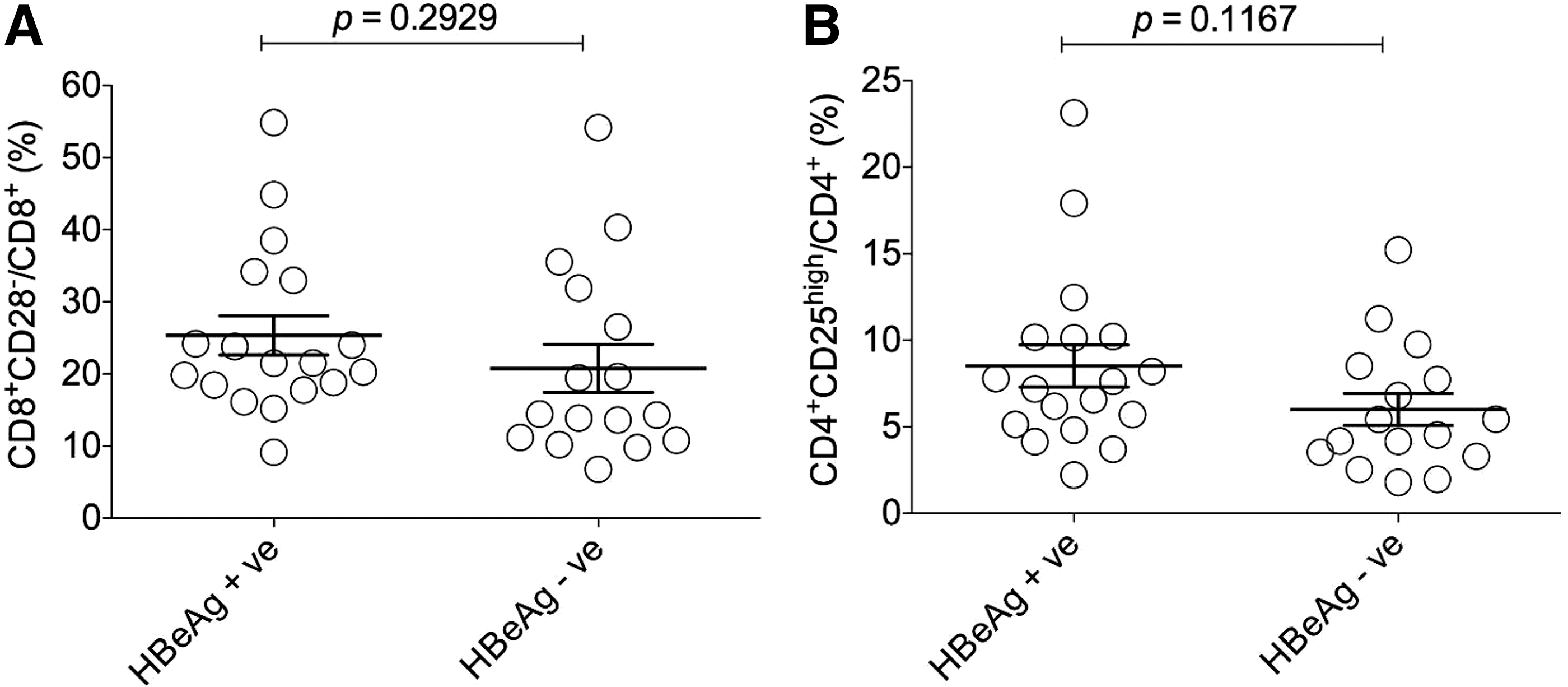
Investigation of the frequencies of CD8+CD28− and CD4+CD25high regulatory T cells in relation to HBeAg in chronic HBV patients.
Discussion
Persistent nonspecific T cell and other immunocyte-mediated immune damage in CHB easily lead to liver injury and further probably cause LC and HCC, the advanced phase of infection progression. During this course, preexisting or induced regulatory factors, including modulatory molecules and regulatory immunocytes, help to relieve immunomediated liver damage and contribute to conservation of tissue integrity (4,27).
However, it is at the cost of prolonging virus clearance. Moreover, aberrant negative regulation usually impairs appropriate and effective anti-HBV immune responses, resulting in HBV immune escape and disease progression. For example, both circulating and liver resident CD4+CD25high regulatory T cells actively influence the antiviral immune response and disease progression in patients with hepatitis B (31). The upregulation of PD-1 on CD4+ and CD8 T+ cells suppresses anti-HBV immune function, resulting in the elevation of HBV load and associating with the increase of ALT and AST levels in CHB patients (32). The increase of PD-L1 expressed on CD4+CD25+Foxp3+ regulatory T cells at CHB stage, and on CD14+ monocytes and CD19+ B lymphocytes at CHB and LC stages is positively related to ALT and AST levels, although it is not relevant to HBV DNA load (7,11).
As a regulatory subset, CD8+CD28− T cell population is still not explored about its influence on the development of HBV infection and association with clinical parameters. This study clearly demonstrated that the frequency of CD8+CD28− T cells increased significantly during HBV infection and had a protective protection on liver integrity and function in chronic phase of infection.
First, we found that during HBV infection, the frequency of CD8+CD28− T cells increased in both chronic and advanced phases, suggesting that CD8+CD28− regulatory T subset is probably upregulated along with the increase of levels of anti-HBV immune responses in a feedback manner within the immune system, or by stimulation of viral antigens.
Second, we showed that the frequency of CD8+CD28− T cells was negatively correlated with the levels of ALT and AST in chronic phase of HBV infection, indicating that this regulatory T subset might exert its effect on suppressing excessive immune responses and prevent liver from immunomediated damage in this phase. However, in the advanced phase, such negative correlation was not found, probably due to downregulation of CD8+CD28− T cell inhibitory function or decrease of weight of CD8+CD28− T cells in regulating immune responses.
Third, we demonstrated that there was no significant relevance between the frequency of CD8+CD28− T cells and HBV DNA load in both chronic and advanced phases, implying that CD8+CD28− T cells do not have a significant influence on viral amplification.
At the same time, we illuminated that there was a discrepant clinical significance between CD28+CD8− and CD4+CD25high regulatory subsets during HBV infection. First, we found that in chronic phase, the frequency of CD28+CD8− T cells was negatively correlated with ALT and AST levels and not related to HBV DNA load, while that of CD4+CD25high regulatory T cells was positively correlated with HBV DNA load and ALT and AST levels. This discrepancy reflects that these two kinds of regulatory T subsets have a different impact on outcomes of regulating immune responses and HBV replication or undergo diverse changes in frequency under HBV antigen stimulation. The negative correlation of frequency of CD28+CD8− T cells within CD8+ population with ALT and AST levels may be because the increase of CD28+CD8− T cells exerts an inhibitory effect on immunomediated liver injury in chronic phase of HBV infection, whereas the positive correlation of frequency of CD4+CD25high regulatory T cells with those clinical parameters may be interpreted as the amplification of HBV and elevation of immune responses attacking virus-infected hepatocytes induce the increase of CD4+CD25high T cells in a feedback manner, for avoiding further expansion of immunomediated liver damage.
Then, we also revealed that in the advanced phase of HBV infection, the frequency of CD28+CD8− T cells did not associate with those clinical parameters, while that of CD4+CD25high regulatory T cells was negatively correlated with HBV DNA load and ALT levels. Such results indicate that in the advanced phase, CD28+CD8− T cells are no longer dominant in regulating immune responses and protecting liver from immunomediated damage, being replaced by CD4+CD25high regulatory T cells.
In addition, the correlation of frequency of CD4+CD25high regulatory T cells with clinical parameters between chronic and advanced phases is the opposite, implying that this regulatory T subset may be upregulated in frequency by HBV stimulation or enhancement of anti-HBV immune responses in a feedback manner within immune system in the chronic phase, and probably exerts its effect on protecting liver from immunomediated damage in the advanced phase.
Also, the clinical significance of CD8+CD28+ T cells in HBV infection was investigated. The decrease of CD8+CD28+ T cells in HBV infection demonstrated that partial CD8+ T cells lost CD28 expression and their immune function was impaired. Furthermore, the positive correlation between the frequency of CD8+CD28+ T cells and liver enzymes in chronic phase suggested that higher proportion of CD8+CD28+ T cells probably contributed to immunomediated liver damage.
A previous study reported that the CD8+CD28+/CD8+CD28− ratio had a weak correlation with HBV clearance in CHB patients (15), but in our study, no significant association between CD8+CD28+ or CD8+CD28+/CD8+CD28− ratio (data not shown) and HBV DNA load was found in both chronic and advanced phase. The range of HBV DNA load (log10 copies 4–9) in our study subjects is different from that (log10 copies 3–11) in their enrolled patients and may account for the inconsistency. Moreover, upregulation of PD-1 on CD8+ T cells in HBV infection (18,32) probably impairs CD28 function on T cell activation, inhibiting CD8+ T cell-mediated viral clearance, since CD28 is a primary target for PD-1-mediated inhibition (12).
In addition, CD8+CD28− and CD4+CD25high regulatory T cells in relation to HBeAg were investigated. The finding that there was no significant difference in the frequency of CD8+CD28+ T cells between HBeAg-negative and HBeAg-positive patients, which was consistent with the frequency of CD8+CD28+ T cells, did not manifest a correlation with viral load, and suggested that viral amplification and antigen release did not have a significant impact on variation of CD8+CD28+ population. At the same time, the frequency of CD4+CD25high regulatory T cells also did not present a significant difference between the HBeAg-negative and HBeAg-positive subjects, which was in line with the discovery in previous report (6).
In summary, in this study, we first substantiated that the frequency of CD8+CD28− T cells increased significantly during HBV infection and had a protective protection on liver integrity and function in chronic phase of HBV infection. At the same time, we clarified that there was a discrepant clinical significance between CD28+CD8− and CD4+CD25high regulatory subsets during HBV Infection. These findings provide potential prognostic markers and therapeutic targets for HBV-infected patients.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (91642103, 31570889), a project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions, Six talent peaks project in Jiangsu Province (2017-SWYY-065), a project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions, Jiangsu Youth Medical Talents Project (QNRC2016226), the Project of Scientific and Technical Development of Suzhou (SS201541, SS201656, LCZX201514), and the Project of Jiangsu Province Association of Preventive Medicine Scientific Research (Y2013023).
Author Disclosure Statement
No competing financial interests exist.