
Abstract
Chronic hepatitis C virus (HCV) infection appears to trigger the onset of immune exhaustion and apoptosis to potentially assist viral persistence inside the host, eventually leading to exacerbated conditions of inflammation and hepatocarcinogenesis. Growing evidence suggests that spontaneous apoptosis of peripheral blood mononuclear cells (PBMCs) could be one of the potential immune impairment mechanisms in chronic viral infection. Interleukin-6 (IL-6) is a pleiotropic cytokine that plays an essential role in regulating immune and inflammatory responses. Owing to its known role in priming T cell growth, differentiation, and inhibition of lymphocyte apoptosis, we investigated the protective effect of IL-6 in rescuing lymphocytes from apoptosis and functional exhaustion in chronic HCV infection. The expression pattern of antiapoptotic (Mcl-1 and Bcl-2), proapoptotic (caspase-3 and Bim) genes along with interferon gamma (IFN-γ) and T cell inhibitory receptor (TIM-3) was analyzed before and after in vitro IL-6 treatment of patient-derived PBMCs. It was observed that the expression of antiapoptotic genes, Mcl-1 and Bcl-2 increased (threefolds and twofolds, respectively) and there was a considerable downregulation in T cell inhibitory receptor (TIM-3) and caspase-3. Similarly, the capacity of PBMCs to produce IFN-γ was also significantly increased (p < 0.001) depicting the promising nature of IL-6 in enhancing lymphocyte effector function. Summing it up, the study supports the positive role of IL-6 in rescuing PBMC population; however, the cytokine alone is not sufficient to sustain the adaptive immunity. It could be used as a potential candidate for combinational therapy along with other regulatory factors for ex vivo enhancement of lymphocyte and may help in moving one step toward adoptive T cell therapy in chronic HCV infection.
Introduction
H
HCV-specific cytotoxic T lymphocytes (CTL) play a leading role in overcoming infection (16). Individuals who clear HCV infection hold a strong and rapid CTL response against multiple viral epitopes (3). Unfortunately, during persistent infection (PI), continuous exposure to antigen renders the T cells inefficient by impeding their effector functions and allowing viral evasion of the immune system, a term known as “T cell exhaustion” (31). In chronic HCV infection, HCV-specific CD8 is depleted by Bcl-2-like protein 11 (BIM)-mediated attrition, and remaining cells are functionally exhausted. T cell apoptosis contributes greatly to the development of chronic HCV persistence (21). The procedure of T cell death during chronic viral infection is determined by a complex group of pro- and antiapoptotic proteins of the Bcl-2 family, such as Mcl-1 and Bim (15). Development of immunomodulatory strategies could be beneficial in restoring the number and efficacy of T cell pool.
Interleukin-6 (IL-6) is a prominent regulator of transition from nonspecific innate immunity to a more specialized and highly specific adaptive immunity against infection. T cell homeostasis and functionality are critically monitored by IL-6 that ensures an efficient and sustained adaptive immune response (25). Several lines of evidence support the notion that IL-6 markedly influences the proliferation and survival of T cells by secreting IL-2 followed by activation of STAT3 and Bcl-2 (2,28,33). IL-6 has an immense potential in differentiating CD8 T cells to CTL subsets (22).
Restoration of T cell function could occur by correcting the levels of Bim and Mcl-1 expression. Previously, IL-6 has been shown to upregulate expression of Mcl-1 messenger RNA (mRNA) and prolong Mcl-1 protein half-life in cholangiocarcinoma cell lines (11) and basal cell carcinoma cells (12). Furthermore IL-6 has been shown to promote accumulation of Mcl-1 (long) in hepatocytes, signifying its antiapoptotic role in liver regeneration (4).Therefore, this study was designed to investigate the antiapoptotic potential of IL-6 in rescuing peripheral blood mononuclear cells (PBMCs) derived from chronic HCV patients. In vitro expansion and inhibition of apoptosis in lymphocytes will help in developing an effective immune response against chronic HCV infection and thus moving one step toward better survival and immunotherapy of these patients.
Materials and Methods
Human subjects
A total of 20 patients chronically infected with HCV were recruited from Armed Forces Institute of Pathology (AFIP) over a period of 6 months. The age group of selected individuals was between 18 and 65 years. Only confirmed cases of chronic HCV treatment-naive category patients were included in the study. Participants having coinfection with other viruses such as hepatitis A, B, or D or having any other comorbidity were excluded from the study. Equal numbers of healthy individuals were used for control and optimization experiments. Peripheral blood was collected from all patients and healthy individuals. The study was approved by the ethical committee of AFIP, Rawalpindi. Written and informed consent was taken from all patients involved in the study.
PBMCs isolation and culture
PBMCs were isolated from fresh blood samples of 20 HCV patients and 15 healthy subjects within 3 h of extraction using Histopaque (Sigma-1077) by Ficoll density gradient centrifugation. Pellet was washed with phosphate-buffered saline (PBS) and either used directly or cryopreserved for later use. To analyze the effect of IL-6 stimulation on PBMCs, cells were cultured in RPMI-1640 media (Gibco) supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin (Sigma). PBMCs were cultured in 12-well plate (2 × 106/well) and wells were divided into control (untreated) and experimental (treated) group. The experimental wells were treated with 5 ng/mL human recombinant IL-6 (Bio Basic). The optimum dose was confirmed through literature and dose–time optimization experiment (Fig. 1). PBMCs were incubated at 37°C and 5% CO2 for 3 h. Recombinant human IL-6 was purchased from Bio Basic.
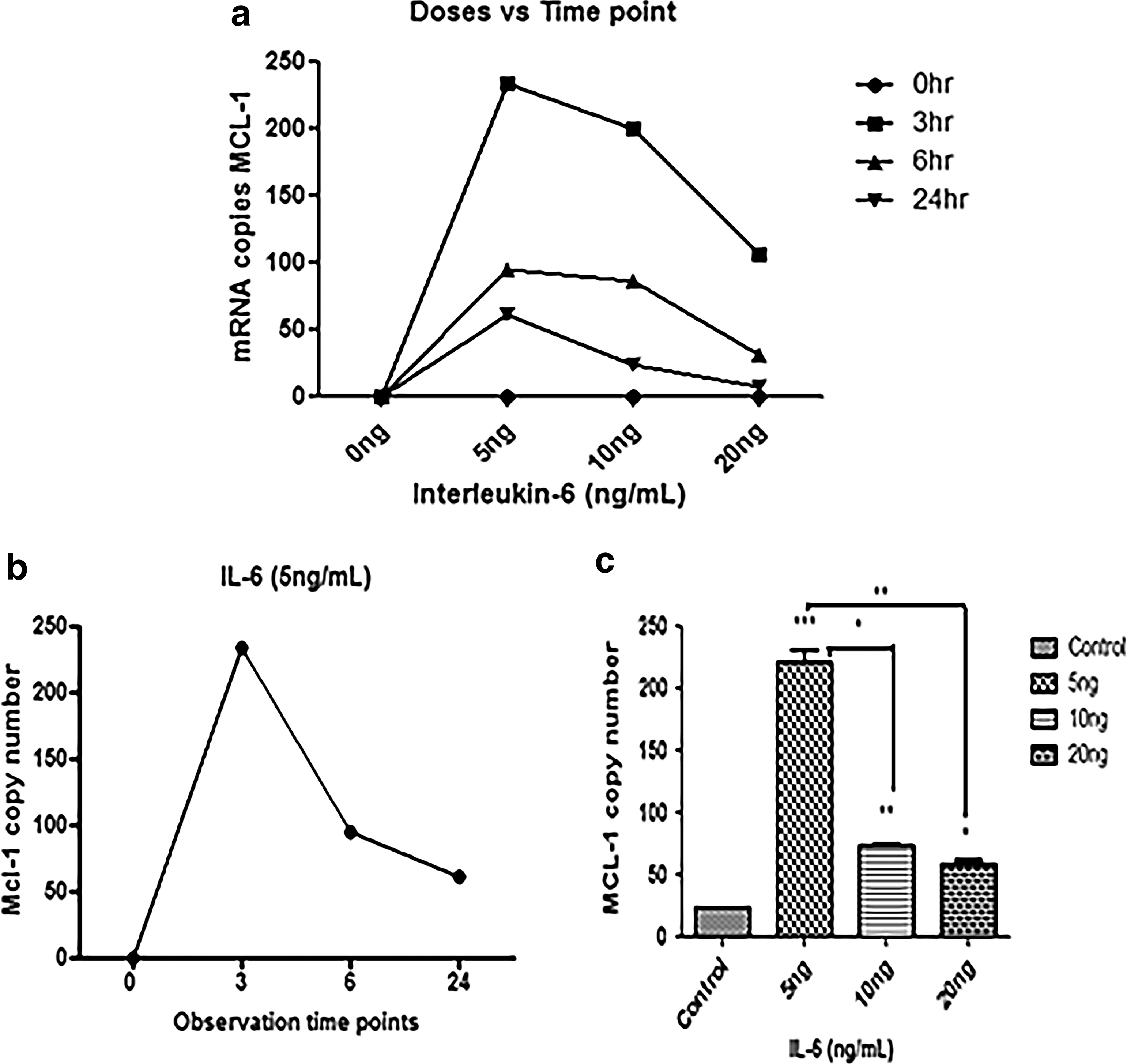
Variation in copy number of MCL-1 with respect to dose and time.
RNA extraction and cDNA synthesis
Cellular RNA was isolated using TRIzol reagent (Invitrogen). Extracted RNA was treated with DNase I (Fermentas). The quantity and quality of extracted RNA were determined using Nanodrop. Furthermore, RNA was qualitatively analyzed on 1% agarose gel stained with ethidium bromide. The complementary cDNA strand was synthesized from this extracted RNA by using oligo (dT) 18 primer (Fermentas) and RevertAid H minus reverse transcriptase (Fermentas) in the presence of Ribolock RNase Inhibitor (Fermentas).
Gene expression analysis in PBMCs
The mRNA levels of interferon gamma (IFN-γ), TIM-3, BIM, MCL-1, BCL-2, and caspase-3 in PBMCs were determined by quantitative real time PCR (qRT-PCR). Targeted genes were quantified on Applied Biosystems 7300 Real-Time PCR system using SYBER Green qPCR Master Mix (2 × )(Fermentas). Rhodamine X reference dye for qRT-PCR was used as passive dye. Gene-specific primers were designed using Primer3 the sequence of which is provided in Table 1. Relative mRNA quantification was calculated by 2−ΔΔCt. All experiments were performed in triplicates and GAPDH was used as housekeeping gene.
Annexin-V staining of PBMCs
Extent of apoptosis was analyzed in both IL-6 stimulated and unstimulated wells, after 3-h culturing. After harvesting and centrifugation, ∼5 × 105 cells were washed with Facs buffer (FACS-PBS) that is used to resuspend cells before and after staining. (PBS supplemented with 0.1% bovine serum albumin, 0.03% sodiumazide, pH 7.2) and fluorescently labeled with anti-CD3 FITC (BD Bioscience) for 30 min at 4°C. Cells were than washed with FACS–PBS and fixed with 7% formaldehyde in dark at room temperature. Next, the cell pellet was resuspended in 100 μL of annexin binding buffer (1 × ) and stained with annexin-V PE antibody (ExBio). Samples were kept in dark for 20 min followed by immediate analysis by CellQuest BD FACS software (BD Biosciences). Both live and apoptotic cells were included in the analysis and cellular debris was excluded. At least 10,000 events were collected by BD flow cytometer and used for further analysis.
Statistical analysis
Statistical analysis was performed using GraphPad Prism version 6.0. Student's t-test and nonparametric Mann–Whitney U-test were performed to analyze statistical difference between two groups; one-way analysis of variance was used for statistical comparison between more than one group. For further evaluation and correlation studies between variables, Spearman rank order test was applied. Significant differences marked on figures are *p < 0.05, **p < 0.005, and ***p < 0.001.
Results
Cytokine dose and time optimization
To determine the ideal dose and time point for the expression of our target gene Mcl-1, we stimulated PBMCs derived from healthy individuals with three different doses (5, 10, and 20 ng) at three time points 3, 6, and 24 h, respectively. Cells were harvested at each time point and further processed for quantitative analysis of Mcl-1 mRNA levels through qRT-PCR. Results were analyzed by GraphPad Prism 6 and copy numbers of Mcl-1 were plotted against respective doses and time points (Fig. 1). Maximum Mcl-1 expression was observed with 5 ng/mL IL-6 dose and after 3 h of stimulation. It was inferred that the expression of Mcl-1 reduced with further increase in IL-6 dose and prolonged time of stimulation experiment.
IL-6 inhibits mitochondrial apoptosis pathway
To investigate how IL-6 regulates other set of genes involved in mitochondrial apoptosis, transcriptional expression of pro- and antiapoptotic genes was evaluated in control and experimental groups 3 h after stimulation of IL-6 (5 ng). As the optimum dose of IL-6 was already determined based on reference gene Mcl-1, rest of the experiments conducted on PBMC samples were performed with 5 ng/mL of IL-6 stimulation for 3 h. The expression level of antiapoptotic genes Mcl-1 and Bcl-2 was significantly increased. However, there was no significant difference in expression of proapoptotic genes, caspase-3 and Bim (Fig. 2). Collectively, this shows that the overall expression of antiapoptotic members is increased and no significant variation was observed in proapoptotic member Bim.
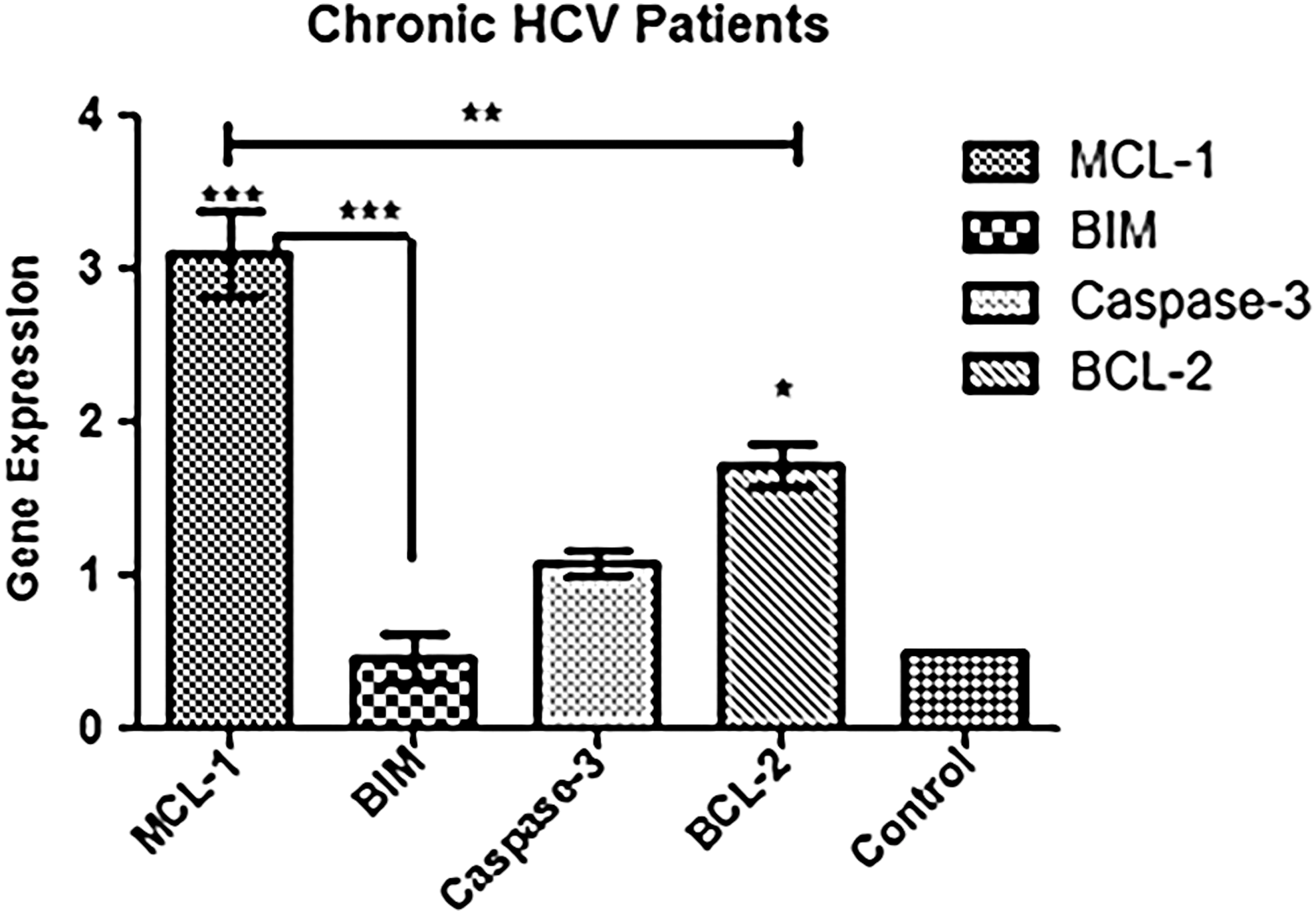
Effect of IL-6 on Bcl-2 family proteins. qPCR analysis of mRNA levels of indicated genes in PBMC samples treated with IL-6 5 ng/3 h. Data are presented in the form of fold change relative to corresponding mRNA levels in untreated control. Relative gene expression was calculated using 2-DDCt method with GAPDH as reference gene for normalization. Error bars represent mean ± standard deviation of three independent experiments. The transcriptional analysis showed that there was a significant increase in 3-folds and 1.5-folds in gene expression analysis of Mcl-1 and Bcl-2, respectively. However, no significant difference was observed in case of BIM and caspase-3. Data were analyzed by applying one-way ANOVA. *p < 0.05, **p < 0.005, ***p < 0.001 showed different levels of significance. PBMC, peripheral blood mononuclear cell; qPCR, quantitative real time PCR.
IL-6 negatively effects expression of T cell inhibitory receptors
It was inquired whether IL-6 has any effect on T cell inhibitory receptor Tim-3 whose expression is significantly increased on lymphocytes and is an important hallmark of T cell dysfunction and exhaustion. We observed that the transcriptional expression of Tim-3 was reduced after 3-h postculture with IL-6. However, the change was not statistically significant (Fig. 3).
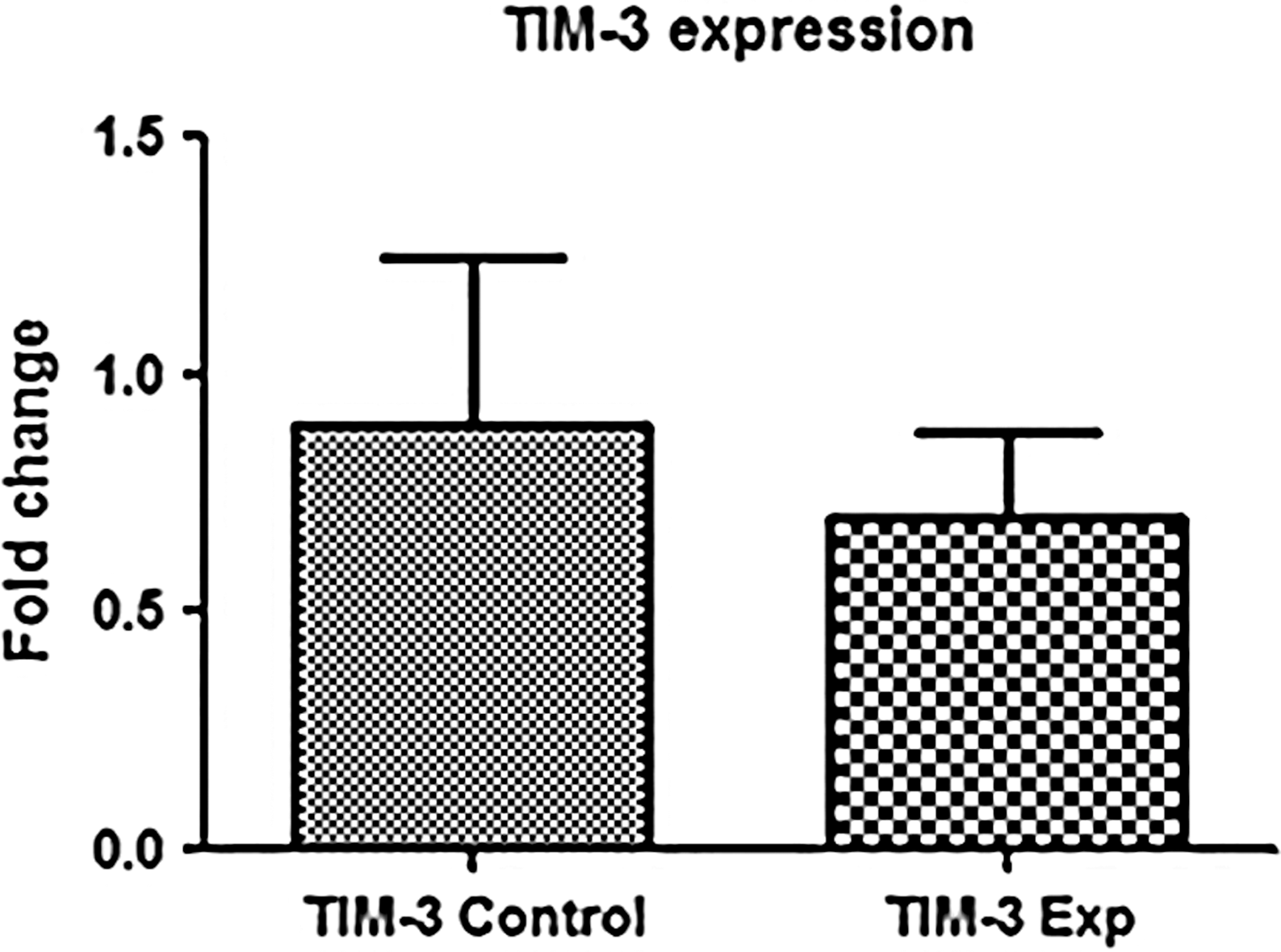
Effect of IL-6 on T cell inhibitory receptor (TIM-3). It was observed that the expression of TIM-3 was reduced in experimental groups 3 h poststimulation with IL-6. Data were analyzed by nonparametric Mann–Whitney U-test and p < 0.05 was considered significant.
IL-6 supports IFN-γ production in lymphocytes
The ability of effector lymphocytes to produce IFN-γ is greatly reduced in patients with persistent HCV infection. We inspected the capacity of IL-6 to increase IFN-γ production in PBMCs, poststimulation. Surprisingly the mRNA levels of IFN-γ were significantly increased in PBMC cultures treated with IL-6 compared with those without IL-6 (Fig. 4). This shows that IL-6 has a noticeable role in reviving lymphocyte effector function in ex vivo cultures.
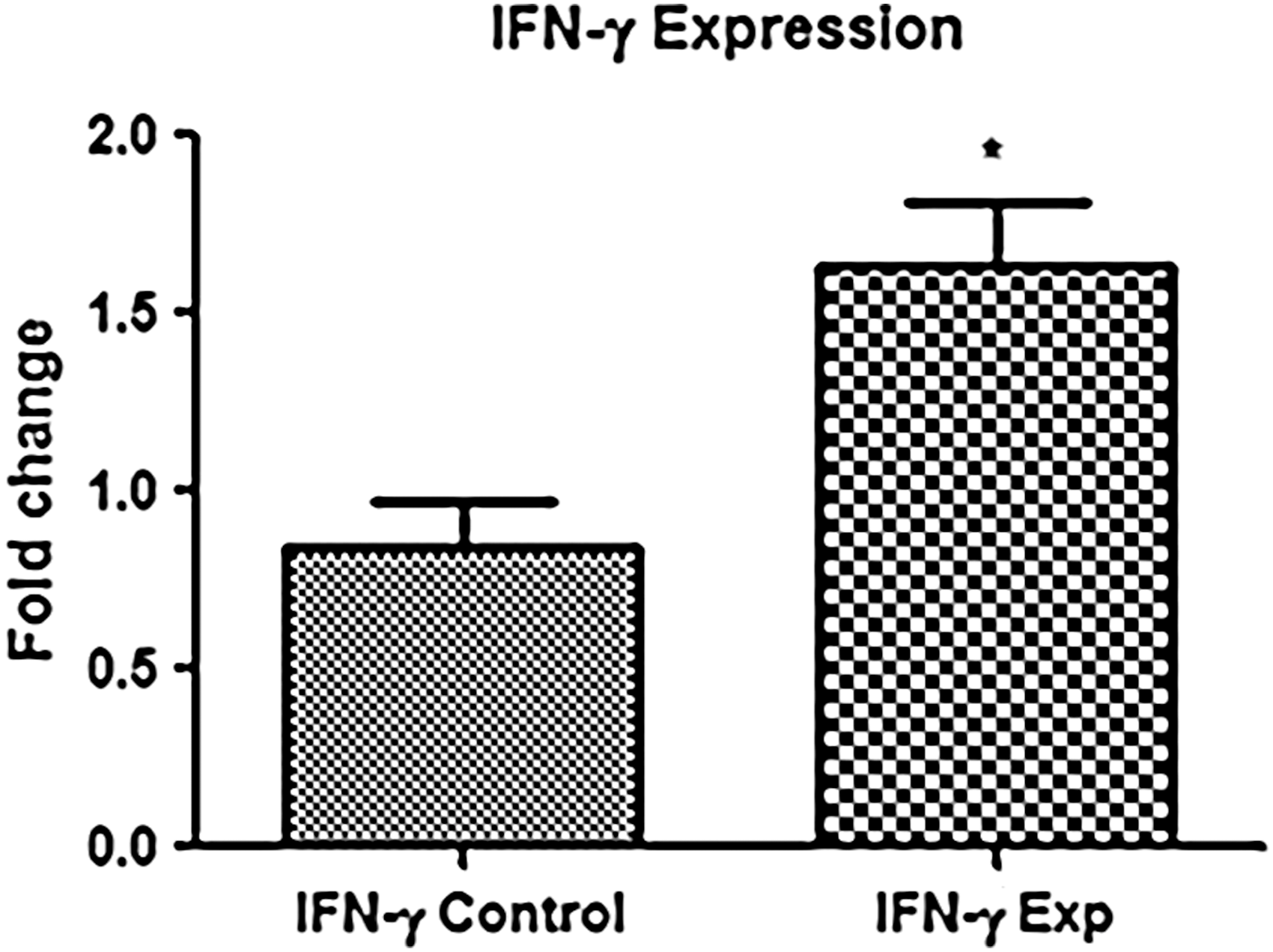
Effect of IL-6 on IFN-γ production. It was observed that the expression of IFN-γ was significantly increased in experimental groups 3 h poststimulation with IL-6. Data were analyzed by nonparametric Mann–Whitney U-test and *p < 0.05 was considered significant. IFN-γ, interferon gamma.
IL-6 restrains spontaneous apoptosis in PBMCs derived from HCV subjects
The effect of IL-6 treatment was evaluated in PBMCs from chronic HCV-infected subjects (n = 20). Ex vivo cultures of PBMCs were treated with or without recombinant IL-6 (5 ng/mL) for 3 h. Cells were then harvested and stained with annexin V and anti-CD3 to determine the extent of apoptosis. Analysis of cell surface-stained cells was performed by flow cytometry. The proportion of annexin-positive cells was higher in control samples than IL-6-stimulated cultures (Fig. 5). The introduction of exogenous IL-6 resulted in a significant reduction of apoptotic cells in all tested experimental groups. The percentage of apoptotic cell population was significantly decreased in IL-6-treated samples compared with control groups (Fig. 6). Furthermore screening of PBMCs with anti-CD3 indicates that IL-6 may have a potential in maintaining survival of lymphocyte population (Fig. 7). The other reason could be that IL-6 induces T cell division thus producing more cells.
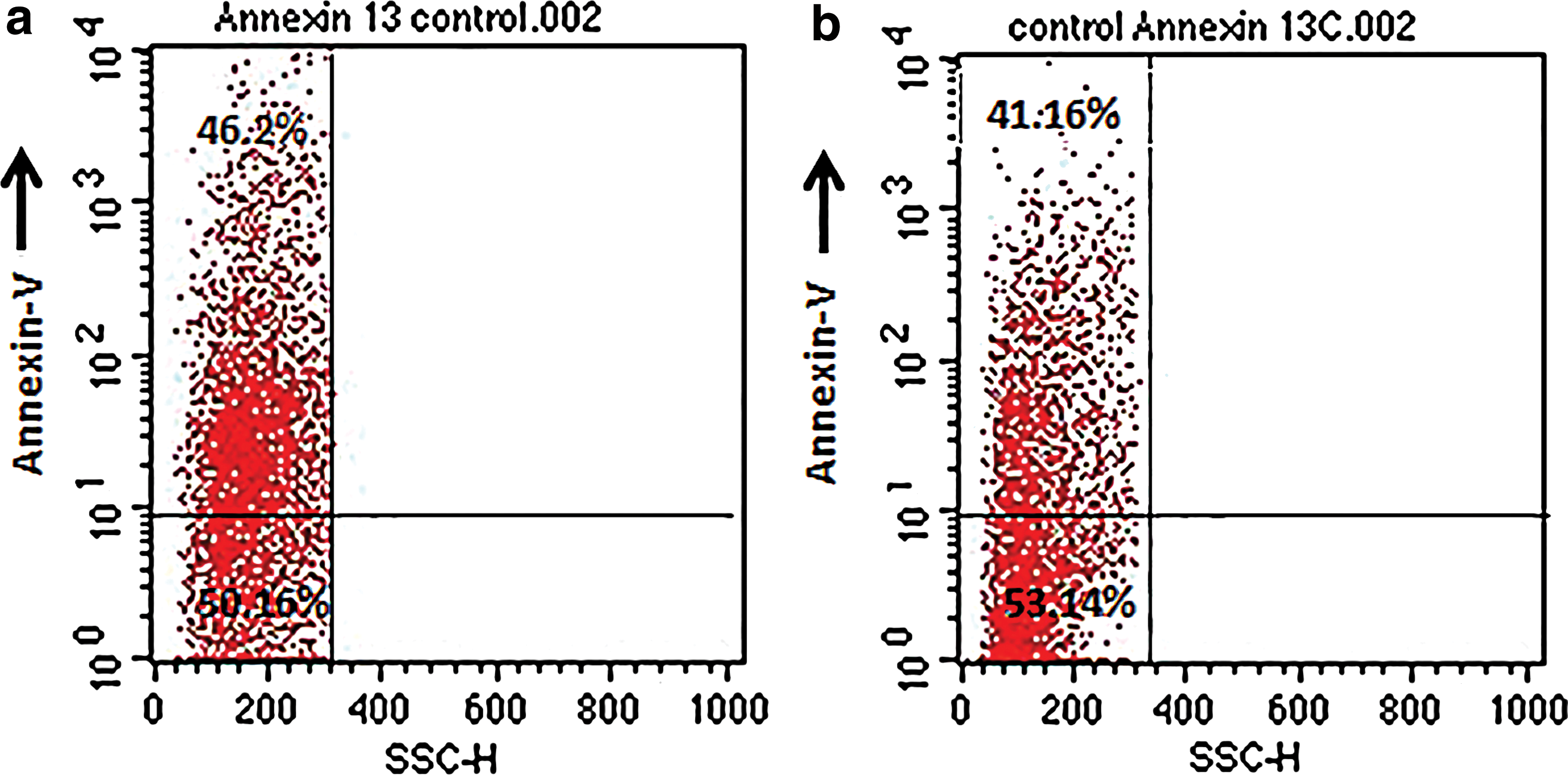
IL-6 treatment increases survival of PBMCs.
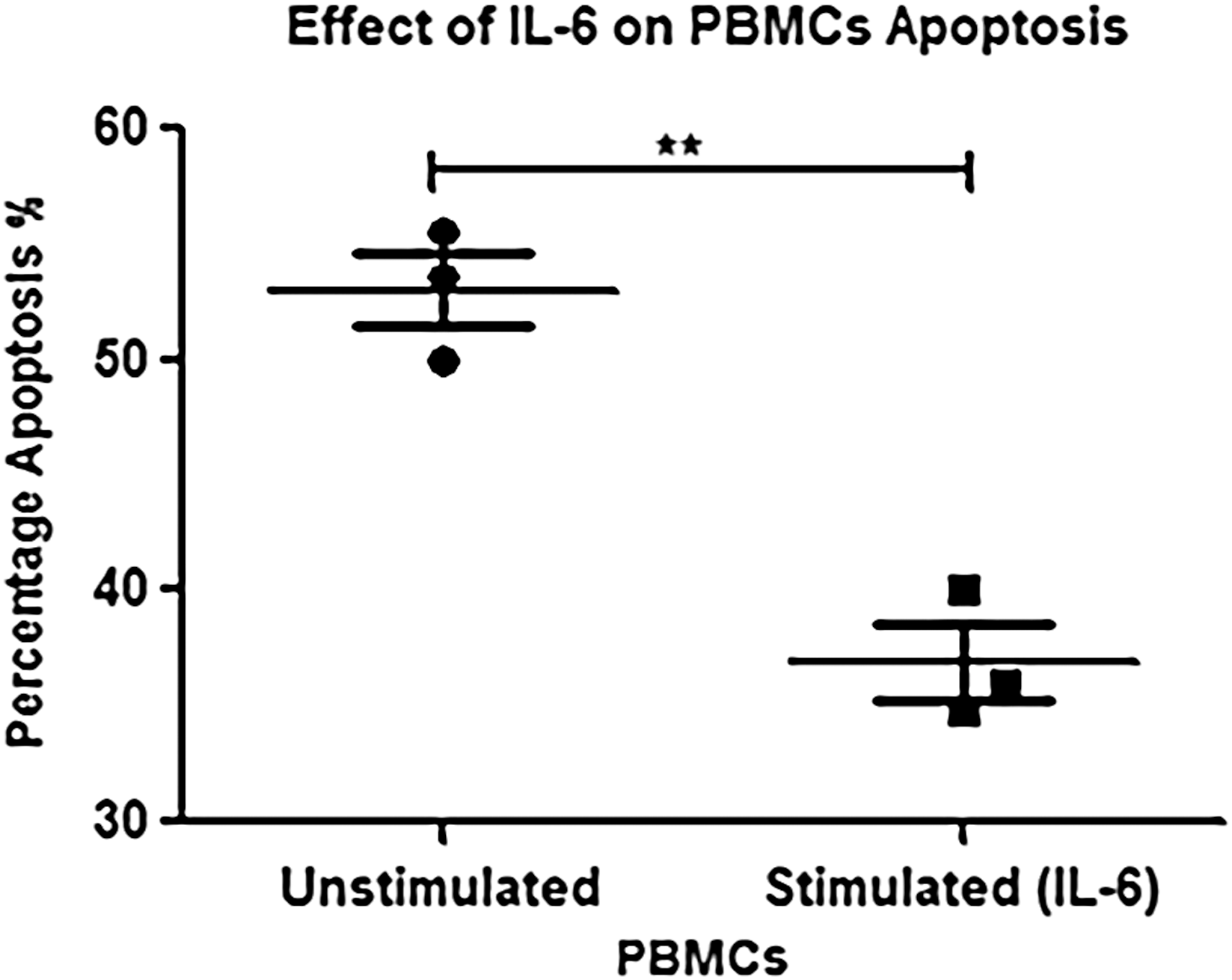
Effect of IL-6 on percentage apoptosis of PBMCs. Significant difference (**p < 0.001) was observed in IL-6-stimulated peripheral mononuclear cells versus unstimulated cells showing a reduction in percentage of cell death.
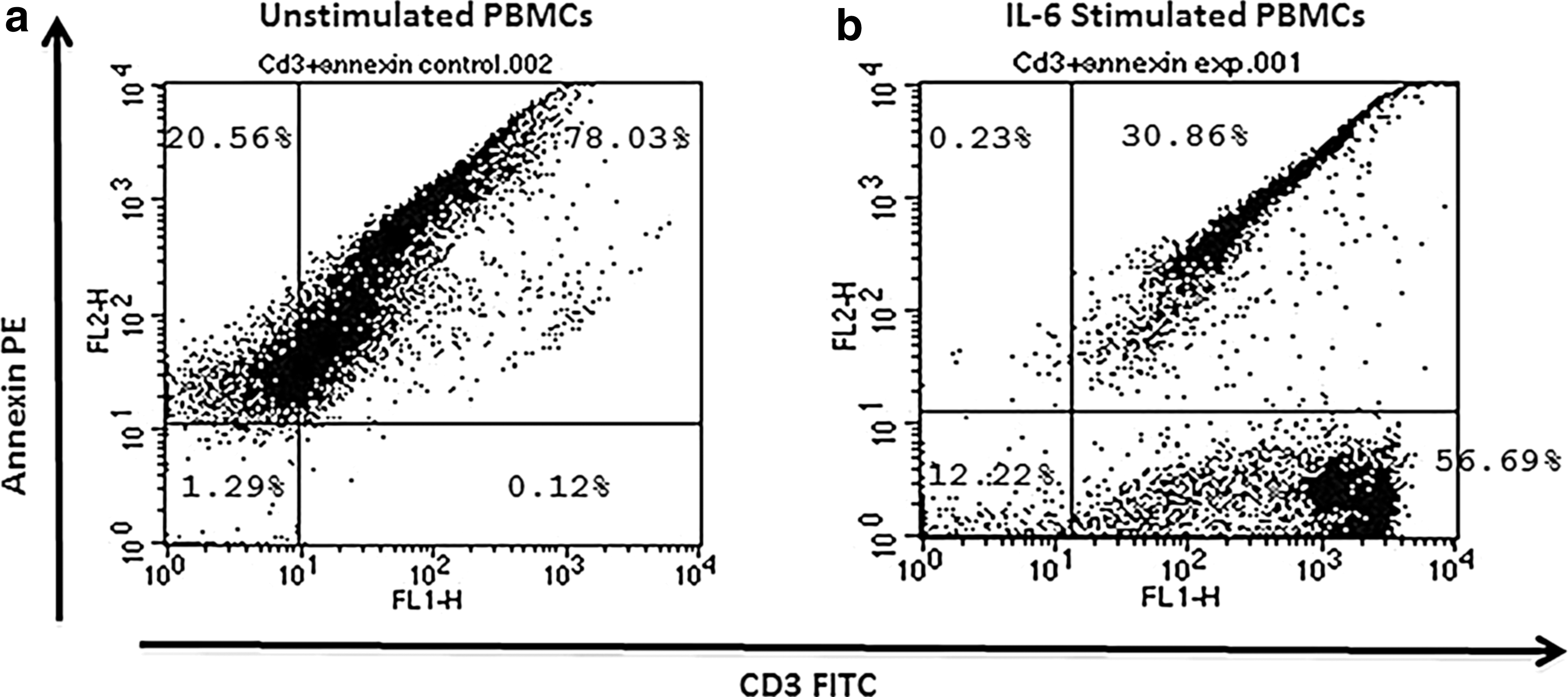
Effect of IL-6 on survival of lymphocyte population.
Discussion
Multiple mechanisms combine to drive T cell exhaustion and limit effective antiviral responses in the setting of chronic viral infections. In individuals where HCV infection is not resolved, the virus promotes functional exhaustion of HCV-specific T cells, anergy, and deletion because of lack of positive stimulation, upregulation of inhibitory receptors and death markers, and rapid expansion of regulatory T cells. In fact CD8 T cells lose their effector property and are selectively deleted in numerous cases, resulting in failure to control viral infection (14). Exhaustion of these cells is associated with expression of inhibitory receptors (PD-1, TIM-3, 2B4, CTLA-4, and CD160), reduced expression of cytokine receptors, and INF-γ production (6). During PI, the frequency of virus-specific peripheral CD8 T cells falls below the detection threshold suggesting deletion or severe impairment of these immune subsets, whereas presence of detectable levels could represent less intense exhaustion (23). HCV-specific CTLs are essential effector tools to control HCV infection, but they are anergic and prone to apoptosis during PI (8). Our study was aimed at elucidating the potential of IL-6 in restricting apoptosis and functional exhaustion in overall population of T lymphocytes in pool of PBMCs derived from chronic HCV patients.
Previously IL-6 has been shown to rescue T cells from entering apoptosis, a process that seems to rely on STAT3-dependent upregulation of antiapoptotic regulators (Bcl-2 family) and modulation of Fas surface expression in other malignancies (5). IL-6 directly acts as a survival factor for CD8/CD4 T cells in in vitro experiments and also suppresses TGF-β-induced apoptosis in CD8 T cells (20). We explored the cytokine IL-6 as a potential homeostatic cytokine that can expand T cell space by inducing expression of antiapoptotic genes, such as Mcl-1 and Bcl-2 and by downregulating expression of T cell inhibitory receptor Tim-3. Similarly, other genes such as Bim, caspase-3, and IFN-γ were investigated to decipher the effect of IL-6 on their respective mRNA expressions. As IL-6 expression is highly upregulated during immune activation and inflammation, IL-6-mediated T cell survival could represent a mechanism to maintain the T cell pool when signaling by other homeostatic cytokines becomes limiting.
It has been previously established that IL-6 has antiapoptotic potential and plays a major role in developing carcinogenesis by disturbing the normal balance of Bcl-2 family proteins in inflammatory conditions. However, the important role of IL-6 in regulating apoptosis could be beneficial in rescuing PBMCs and lymphocyte subsets from spontaneous cell death and exhaustion induced as a result of viral persistence. Therefore, this study was designed to explore the opportunities of using IL-6 for ex vivo modification and enhancement of PBMCs derived from chronic HCV patients. Interestingly the expression of proapoptotic members, Mcl-1 and Bcl-2, was greatly increased after stimulating PBMC cultures with IL-6 (5 ng/mL). Moreover, no significant change was observed in case of Bim, an important member of Bcl-2 family, which sequesters Mcl-1 and initiates mitochondrial apoptosis pathway. Resistance to apoptosis by upregulating antiapoptotic proteins can allow prolonged survival of T cells in the absence of nutrients and growth factors (19). T cell death as a result of exhaustion or deprivation of cytokine and growth signal is controlled by Bcl-2 family members such as Bim and Mcl-1. Along with other proapoptotic members, Bim promotes cell death through mitochondrial/intrinsic apoptosis and its action on intrahepatic T cells is counteracted by Mcl-1 (9,10). Similarly another study supports that IL-6 has a great potential in preventing T lymphocytes from entering apoptosis by upregulating antiapoptotic proteins (Bcl-2, Bcl-xl) and modulating Fas receptor expression (13).
Another interesting aspect of this study was to investigate the effect of IL-6 on inhibitory receptor Tim-3, one of the important hallmarks of T cell dysfunction and exhaustion. The expression of Tim-3 was reduced in experimental groups compared with controls without IL-6 showing a promising nature of IL-6 in regulating T cell inhibitory receptors. Although the variation in expression was not significant, there is still hope that IL-6 may influence expression of Tim-3 along with other inhibitory receptors PD-1, CTLA-4, BTLA-4, and LAG-3. Exposure to chronic infection alone is sufficient to impair cytotoxic activity and differentiation of CD8+ T cells. Inflammatory pathways induced by IL-6 and IL-27 may assist in controlling viral infection and reducing the risk of CD8+ T cell exhaustion. Together these studies indicate that IL-6 can help in overcoming chronic viral infection and preventing T cell exhaustion through other downstream effector cytokines (32). Consistently, they also have a well-known potential in modulating CD4+ T cell differentiation during chronic viral infections that further strengthens our hypothesis (32).
To investigate the capacity of PBMCs to produce IFN-γ, we determined levels of IFN-γ poststimulation. Surprisingly the mRNA levels of IFN-γ were significantly increased in PBMC cultures treated with IL-6 compared with those without IL-6. Immunity to viral infections greatly rely on the production of IFN-γ-producing T cell response. Previous studies have suggested a protected role of IL-6 on T lymphocyte subsets. It was demonstrated that IL-6 is necessary for generation of IFN-γ response during Mycobacterium tuberculosis infection (26). It is now a well-established fact that IL-6 can monitor the initiation and development of both innate and adaptive immune response (29).
The data presented here strongly support the idea that IL-6 has a definitive role in maintaining immunity that cannot be compensated for by other cytokines. Annexin V staining of PBMC populations in this study also revealed interesting findings. The number of stained cells was reduced in experimental groups showing a limited number of cells undergoing programmed cell death. This further clarifies that IL-6 can act as a promising candidate and a survival factor for lymphocytes. Supporting our findings, it has been documented that IL-6 signaling promotes T cell proliferation and prevents them from undergoing apoptosis by producing IL-2 and activating STAT3 (27). IL-6 also assists in T cell survival, regulating helper T cell decisions and chemokine receptor expression, thereby assisting recruitment of T cells to tissues. Accordingly, loss of IL-6 signaling may rescue dysfunctional T cells and influence T cell differentiation and recruitment during inflammation (30). Other proinflammatory cytokines (IL-1β and IL-18) also have prominent role in shaping T cell activation and determining their fate. Administration of exogenous IL-1β allows naive CD8 T cells to respond to antigen and lipopolysaccharides to form numerous functionally effective memory cells (24). Similarly IL-18 that resembles IL-1β in structure also contributes in Th1 cell differentiation and when used in combination with IL-12 plays an important role in activating CTLs to produce IFN-γ (7).
Conclusion
The findings in this study conclude that IL-6 has a strong potential in regulating mechanism of mitochondrial apoptosis in lymphocytes derived from chronic HCV patients. IL-6 has been found to upregulate antiapoptotic proteins, reduce the expression of T cell inhibitory receptors, and increases the production of IFN-γ thereby rescuing lymphocytes from exhaustion and apoptosis induced by PIs. Besides regulating the expression of candidate genes involved in apoptosis and exhaustion, IL-6 also helps in maintaining integrity of PBMCs as only few cells were found to be depleted through programmed cell death. It could be beneficial in moving one step toward ex vivo expansion and modification of lymphocytes for adoptive T cell therapy of chronic HCV patients. Although IL-6 alone is not sufficient to produce a stable and robust response, when used in combination with other entities it could prove to be a successful candidate. Further studies need to be conducted to completely understand the nature of IL-6 in regulating survival of lymphocytes especially in persistent viral infections where they are prone to functional exhaustion and cell death that ultimately collapses the immune system of an individual.
Footnotes
Author Disclosure Statement
No competing financial interests exist.