
Abstract
Nod-like receptor protein 3 (NLRP3), absent in melanoma 2 (AIM2), and interferon gamma inducible protein 16 (IFI16) are innate immune sensors for intracellular microbes, which can be activated by various dangerous signals and subsequently lead to caspase-1 (CASP1) activation and the maturation cleavage of effector molecules pro-IL-1β and pro-IL-18. Their roles in immunopathology of acute and chronic hepatitis B virus (HBV) infection are still unclear. In this study, we first investigated the activation of NLRP3, AIM2, and IFI16 inflammasomes in peripheral blood mononuclear cells (PBMCs) from patients infected with acute hepatitis B (AHB) and chronic hepatitis B (CHB) by quantitative real-time PCR and enzyme-linked immunosorbent assay. We next analyzed the impact of hepatitis B e antigen (HBeAg) on activation of AIM2 and IFI16 inflammasomes in PBMCs of CHB patients stimulated in vitro with AIM2 and IFI16 agonist ligands, poly (dA:dT) and VACA-70mer, respectively. The results showed that the mRNA expression levels of AIM2, IFI16, and CASP1 in PBMCs from AHB and CHB patients were both upregulated. Furthermore, the mRNA levels of AIM2 and IFI16 in CHB patients were significantly positively correlated with serum HBV loads. However, only in patients with AHB there was elevation of serum IL-1β and IL-18. There was no activation of NLRP3, AIM2, and IFI16 inflammasomes in CHB patients. Stimulation of PBMCs of CHB patients in vitro with poly (dA:dT) and VACA-70mer induced the activation of AIM2 and IFI16 inflammasomes, respectively. This ligand-induced activation was suppressed by HBeAg. Our results suggest that there exists activation of the AIM2 and IFI16 inflammasomes, but not the NLRP3 inflammasome, in AHB, and the activation of the AIM2 and IFI16 inflammasomes can be inhibited by HBeAg in CHB, which may contribute to HBV-induced immunotolerance.
Introduction
H
Various pattern-recognition receptors (PRRs) that recognize conserved motifs on pathogens are responsible for induction of host innate immune responses. Activation of some PRR family members, such as Nod-like receptor protein 3 (NLRP3), absent in melanoma 2 (AIM2), and interferon gamma inducible protein 16 (IFI16), initiates the formation of inflammasomes. The classical inflammasome consists of a PRR (which can be either a NOD-like receptor [NLR] or an AIM2-like receptor [ALR] protein), an adaptor molecule (apoptosis-associated speck-like protein containing a caspase recruitment domain [ASC]), and a procaspase-1 (21). Inflammasome activation leads to autoactivation of procaspase-1, resulting in the mature cleavage of pro-IL-1β and pro-IL-18. IL-1β and IL-18 are pleiotropic proinflammatory cytokines. IL-1β has been shown to exhibit protective properties against microbial infection (5). IL-18 promotes interferon-γ (IFN-γ) production in natural killer (NK) and T cells, and enhances proliferation and cytotoxic activity of CD8+ T and NK cells (18). Thus, inflammasomes play critical roles in host defense against microbes.
However, the role of NLR and ALR inflammasomes in HBV infection is less understood. It has been documented that many viruses can trigger activation of NLR and/or ALR inflammasomes, and meanwhile some viruses have evolved strategies to counteract these innate antiviral pathways and promote persistent infections (16). Previous studies have suggested that inflammasomes are involved in HBV infection (2,6,8,20,26,31). However, whether activation of inflammasomes promotes HBV clearance during acute infection or HBV can counteract activation of inflammasomes to facilitate persistent infection is yet to be determined. In this study, we determined the expression and activation of the NLRP3, AIM2, and IFI16 inflammasomes in acute hepatitis B (AHB) and chronic hepatitis B (CHB) patients, and explored the impact of hepatitis B e antigen (HBeAg) on activation of AIM2 and IFI16 inflammasomes in peripheral blood mononuclear cells (PBMCs) of CHB patients in vitro.
Materials and Methods
Subjects
From May 2015 to September 2017, 54 patients with HBeAg-positive CHB (n = 34) and AHB (n = 20) from Shenzhen people's hospital (Shenzhen, China) were enrolled in this study. Twenty-seven healthy individuals were enrolled as healthy controls (HCs). The clinical characteristics of HCs and patients are shown in Table 1. All of the attendees signed an informed consent form before sample collection. The study was approved by the Ethics Committee of Shenzhen people's hospital.
Values are n or the median with range.
χ 2 test.
Kruskal–Wallis H test.
Mann–Whitney U test.
AHB, acute hepatitis B patients; ALT, alanine aminotransferase; CHB, chronic hepatitis B patients; HBeAg, hepatitis B e antigen; HBV, hepatitis B virus; HC, healthy controls; ND, not determined; NS, not significant.
The CHB disease was diagnosed according to the guidelines of CHB (15). The AHB disease was diagnosed according to the Chinese Management Scheme of Diagnostic and Therapy Criteria of Viral Hepatitis (3). All CHB patients had a serum HBV DNA load >20,000 IU/mL and an alanine aminotransferase (ALT) level greater than two times the upper limit of normal. All patients were treatment naive and did not display any other diseases.
Isolation of PBMCs
Ten milliliters of venous blood was collected from each subject. PBMCs were separated using Ficoll-based (BD Biosciences) density gradients.
RNA extraction, reverse transcription, and quantitative real-time PCR
Total RNA of PBMCs was extracted with TRIzol (Invitrogen). Reverse transcription was conducted using the PrimeScript RT reagent kit (TaKaRa). The primers used in this experiment were as follows: NLRP3 (forward) 5′-GATCTTCGCTGCGATCAACA-3′ and (reverse) 5′-GGGATTCGAAACACGTGCATTA-3′; AIM2 (forward) 5′-CTGCAGTGATGAAGACCATTCGTA-3′ and (reverse) 5′-GGTGCAGCACGTTGCTTTG-3′; IFI16 (forward) 5′-GAAGTGCCAGCGTAACTCCTAA-3′ and (reverse) 5′-TGATTGTGGTCAGTCGTCCAT-3′; CASP1 (forward) 5′-AGTGCAGGACAACCCAGCTATG-3′ and (reverse) 5′-CAAGACGTGTGCGGCTTGA-3′; ASC (forward) 5′-CCTTGGACCTCACCGACAA-3′ and (reverse) 5′-ATGTCGCGCAGCACGTTAG-3′; IL-1β (forward) 5′-CCAGGGACAGGATATGGAGCA-3′ and (reverse) 5′-TTCAACACGCAGGACAGGTACAG-3′; IL-18 (forward) 5′-CTGCCACCTGCTGCAGTCTA-3′ and (reverse) 5′-TCTACTGGTTCAGCAGCCATCTTTA-3′; GAPDH (forward) 5′-GCACCGTCAAGGCTGAGAAC-3′ and (reverse) 5′-TGGTGAAGACGCCAGTGGA-3′. The quantitative real-time PCR (qPCR) was performed using a SYBR Premix ExTaq kit (TaKaRa) in an Applied Biosystems ViiA 7Dx Real-Time PCR system (Applied Biosystems, Shanghai, China). Standard curves for target genes were established using gradient dilutions of the corresponding DNA fragments amplified by real-time PCR (RT-PCR). Based on the corresponding standard curve, the mRNA copies of each gene were determined. The qPCR was performed in duplicate for each sample. The expression levels of each gene are depicted as values normalized against human 103 copies of GAPDH transcripts.
Cell culture and treatment
PBMCs were cultured at 5 × 105 cells/mL in RPMI 1640 containing 10% FBS, 2 mmol/L
Enzyme-linked immunosorbent assays
Protein levels of IL-1β and IL-18 in serum and supernatants were determined by ELISA kits (eBioscience) according to the manufacturer's instructions.
HBV DNA assays and serological assays
Serum HBV DNA levels were quantified by an ABI 7500 Real-Time PCR System (Applied Biosystems). The serum levels of hepatitis B surface antigen, HBeAg, and anti-HBe were detected using reagents from Roche Diagnostics Cobas e601 analyzer. Normal serum ALT levels ranged from 0 to 40 U/L.
Statistical analyses
Data are expressed as mean ± SD. The statistical analyses were conducted using GraphPad 6.0 (GraphPad Software, San Diego, CA). The Student's t-test and Mann–Whitney U test were used for two-group comparisons. When comparing more than two groups, analyses were conducted using one-way ANOVA. Spearman's rank correlation coefficient was used to calculate correlations. p < 0.05 was considered significant.
Results
Differential expression and activation of NLRP3, AIM2, and IFI16 inflammasomes in PBMCs from patients with AHB and CHB
To determine whether HBV infection could induce activation of NLRP3, AIM2, and IFI16 inflammasomes, we first analyzed their components' expression in patients with AHB and CHB. As shown in Figure 1C–E, compared with HC, the mRNA levels of AIM2, IFI16, and CASP1 were significantly upregulated in AHB and CHB groups. However, both the AHB and CHB groups had comparable ASC and NLRP3 expression levels with the HC group (Fig. 1A, B). These results, which are consistent with other observations (26,29), suggest that both acute and chronic HBV infections may result in the formation of AIM2 and IFI16 inflammasome complexes.
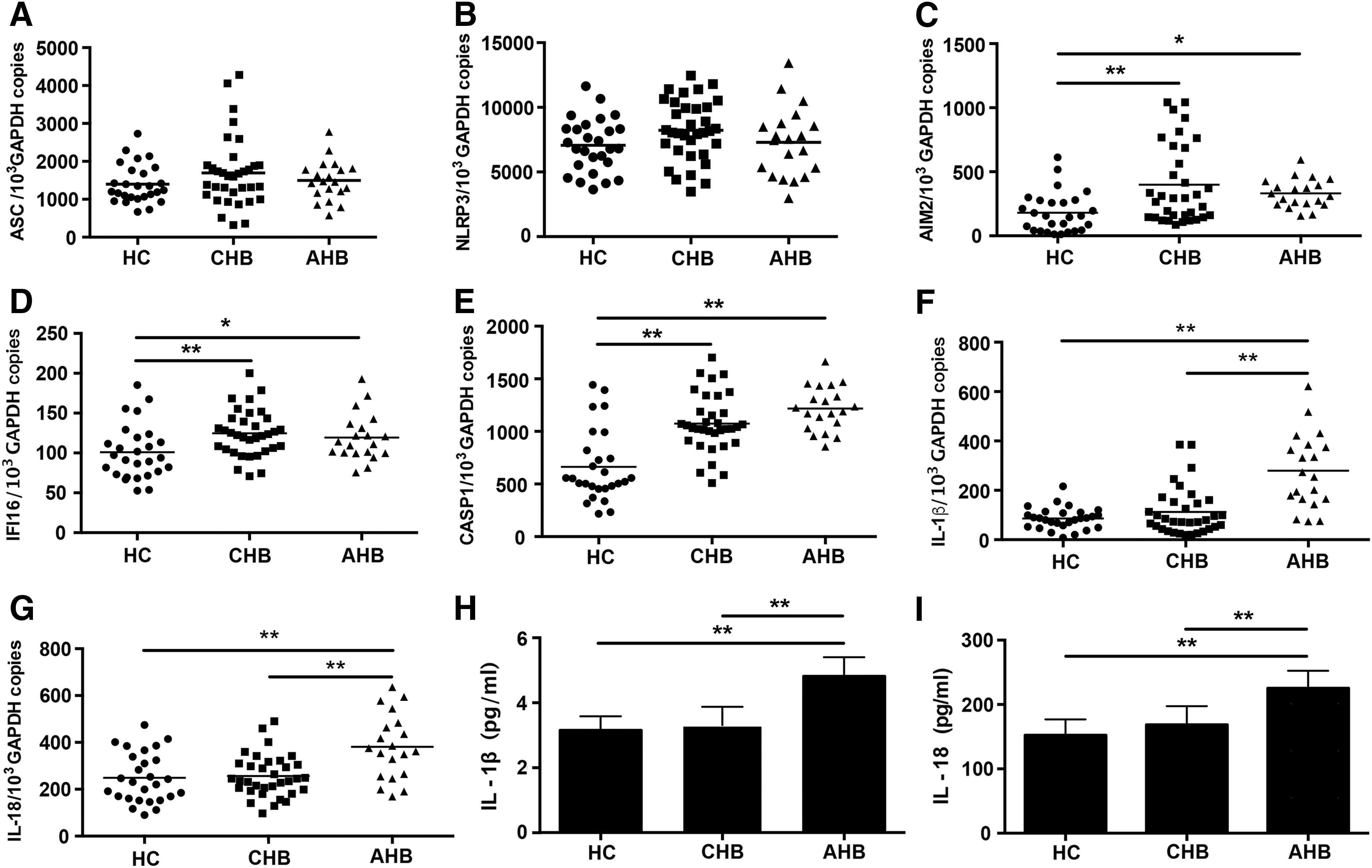
Differential expression and activation of NLRP3, AIM2, and IFI16 inflammasomes in PBMCs from patients with AHB and CHB. The amounts of ASC, NLRP3, AIM2, IFI16, CASP1, IL-1β, and IL-18 mRNA were quantified using qPCR
To clarify whether the upregulation of the components of AIM2 and IFI16 inflammasomes triggers increased secretion of IL-1β and IL-18, we measured these two cytokines mRNA levels in PBMCs and protein levels in serum from AHB and CHB groups. The results indicated that both the mRNA and protein levels of IL-1β and IL-18 were significantly increased in the AHB group (Fig. 1F–I). In contrast, no significances were observed in the expression and secretion levels of IL-1β and IL-18 between the HC and CHB group (Fig. 1F–I), which is consistent with results from previous observations (29), suggesting that the expression of IL-1β and IL-18 is suppressed in CHB.
AIM2 and IFI16 mRNA levels are significantly positively correlated with serum HBV DNA loads in CHB patients
To address the question which HBV component may stimulate the AIM2 and IFI16 mRNA expression, we analyzed correlation of AIM2 and IFI16 mRNA with serum HBsAg, HBeAg levels and HBV DNA loads. The results indicated that both AIM2 and IFI16 mRNA levels were significantly positively correlated with serum HBV DNA loads in the CHB group (Fig. 2), which is consistent with earlier reports (6). In contrast, IFI16 mRNA levels were negatively correlated with HBeAg levels. Due to the clinical characteristics of AHB (7), all the AHB patients enrolled at the first visit due to liver damage had low levels of HBV DNA. Therefore, no correlation was observed between the mRNA levels of AIM2 and IFI16 and HBV DNA loads in AHB patients (data not shown). Taken together, these data imply that upregulated expression of AIM2 and IFI16 mRNA may be stimulated by HBV DNA.
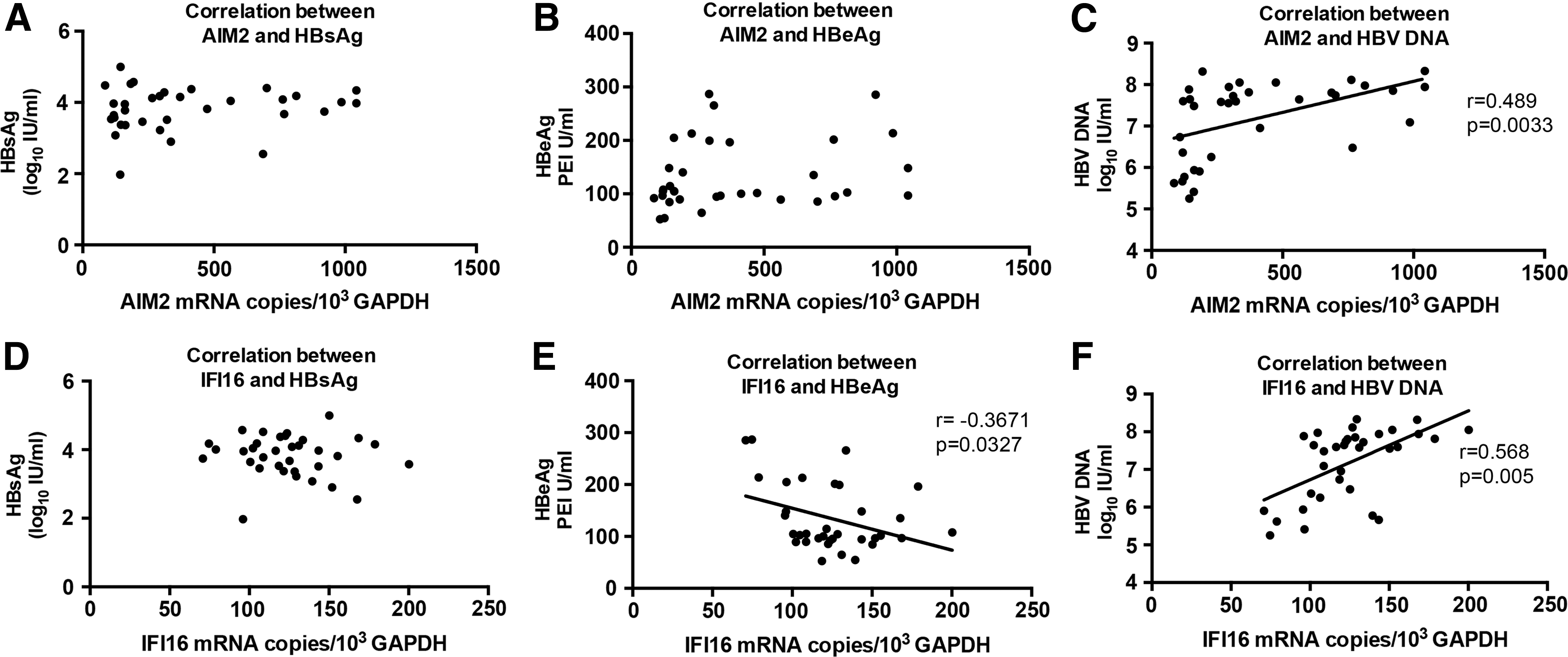
Correlation of AIM2 and IFI16 mRNA with HBsAg, HBeAg, and HBV DNA levels. Correlation of AIM2 mRNA with HBsAg, HBeAg, and HBV DNA levels
HBeAg can inhibit the activation of AIM2 and IFI16 inflammasomes in PBMCs stimulated by their respective ligands in vitro
AIM2 and IFI16 are intracellular innate DNA sensors (9,23), which can be induced by various viruses (16). Based on the correlation analyses, it can be inferred that HBV DNA may stimulate the upregulation of AIM2 and IFI16 expression in PBMCs of CHB patients. However, activation of the inflammasomes was not observed in CHB patients. As HBeAg could inhibit LPS-induced NLRP3 inflammasome activation (29), we speculate that HBeAg may also inhibit the activation of the AIM2 and IFI16 inflammasomes in CHB patients. Previous studies showed that poly (dA:dT) and VACA ds 70 mer, which are AIM2 and IFI16 ligands, respectively, could induce the transcription of AIM2 and IFI16 in PBMCs from woodchucks (28). Therefore, We isolated PBMCs from CHB patients, stimulated them with poly (dA:dT) and VACA ds 70 mer in vitro, respectively, and then treated with or without HBeAg to investigate the impact of HBeAg on the activation of AIM2 and IFI16 inflammasomes.
As shown in Figure 3, after transfection of PBMCs from CHB patients with poly (dA:dT) or VACA ds 70 mer, AIM2 and IFI16 expression increased strongly while NLRP3 and CASP1 expression was not altered. Accordingly, IL-1β mRNA and protein levels were both highly increased after transfection. Treatment with HBeAg after transfection resulted in profound suppression of IL-1β expression. These results indicated that HBeAg inhibited the activation of the AIM2 and IFI16 inflammasomes without affecting the expression of AIM2, IFI16, and CASP1 mRNA.
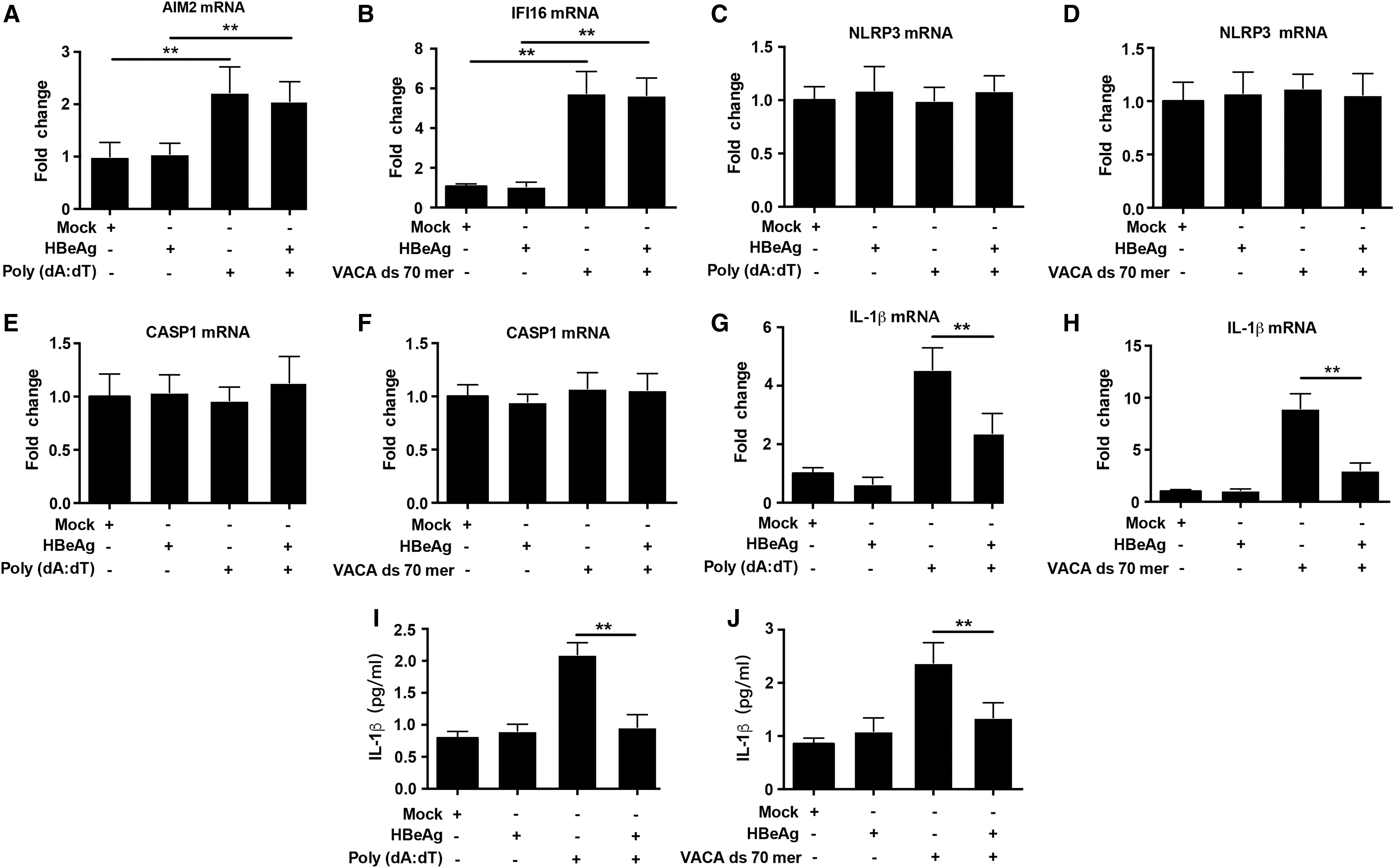
HBeAg inhibits AIM2 and IFI16 inflammasomes activation in PBMCs from CHB patients. PBMCs were mock transfected or transfected with poly (dA:dT) or VACA ds 70 mer (2 μg/mL) in vitro, then treated with or without HBeAg (2 μg/mL). Six hours later, the mRNA levels of AIM2
Discussion
As a constitutive part of innate immune system, inflammasomes have been recognized for their critical role in host defense against pathogens. Their role in HBV infection remains poorly defined. Several studies have explored the relevance of inflammasomes in the pathogenesis of hepatitis B (2,6,20,26,31). Askari et al. showed that mRNA levels of NLRP3 significantly increased in peripheral immune cells from CHB patients with low virus loads compared with HCs (2). However, our results revealed that NLRP3 mRNA levels were not altered in both AHB and CHB patients, consistent with the results recently published by Yu et al., which suggested that HBV could not activate the NLRP3 inflammasome (29). This discrepancy may be partly attributable to the different infection phases that the subjects belong to. AIM2 and IFI16 are members of the ALR family, which can also form inflammasome complexes (16). The AIM2 inflammasome has been shown to be associated with HBV infection (2,6,20,26,31). It was demonstrated that AIM2 mRNA levels were strongly upregulated in livers of CHB patients (8,20). Moreover, hepatic AIM2 protein expression levels were significantly increased in the high HBV replication group compared with the low HBV replication group. In another report, the expression levels of hepatic AIM2 and IL-1β were elevated in woodchucks with acute woodchuck hepatitis virus (WHV) infection (28). Similarly, our results showed that the AIM2 mRNA levels in PBMCs were higher in both AHB and CHB patients than in HCs, and were positively correlated with HBV DNA loads in CHB patients. IFI16, predominantly nuclear as a transcriptional activator, now serves as an innate intracellular DNA sensor that induces IFN-β and IL-1β production through stimulator of interferon genes (STING) and inflammasome activation, respectively (10,23). IFI16 was found to be crucial for suppressing replication of many viruses (4). Li and colleagues showed that the IFI16 and IL-1β mRNA levels were increased in the livers and spleens from woodchucks with acute WHV infection, whereas they were slightly decreased in the livers of woodchucks chronically infected with WHV (28). Upregulated expression of IFI16 was also observed in other chronic or persistent viral infections (1,19). In this study, we demonstrated for the first time that IFI16 mRNA levels in PBMCs were higher in both AHB and CHB patients than in HCs, and were positively correlated with HBV DNA loads in CHB patients. In addition, the IFI16 mRNA levels were negatively correlated with serum HBeAg levels. Our study also showed that in AHB patients despite with low HBV DNA loads, the expression of the AIM2 and IFI16 mRNA was still upregulated. As in the preclinical phase of acute HBV infection, the virus usually replicates to an extremely high level, we speculate that the massive viremia may thereafter stimulate the expression of AIM2 and IFI16 mRNA, which may in turn contribute to the inhibition of viral replication. Taken together, these results suggest that AIM2 and IFI16 may play important roles in the immunopathogenesis of HBV infection.
IL-1β is critical in the host defense against viral infection. A previous study revealed that IL-1β drastically decreased HBV envelope surface protein (HBs) secretion, HBV core protein (HBc) expression, and HBV DNA replication in HepaRG cells (24). IL-18, primarily known as IFN-γ-inducing factor, enhances NK cell activity and IFN-γ production in T cells and NK cells. It has been shown that injection of 10 μg dose of IL-18 could rapidly and noncytopathically inhibit HBV replication in the livers of HBV transgenic mice (11). Our study showed that there was remarkable expression and secretion of IL-1β and IL-18 in AHB patients. However, compared with the enhanced transcription of AIM2 and IFI16, the expression and secretion of IL-1β and IL-18 in CHB patients were not enhanced accordingly, which may contribute to the impaired antiviral immune response and immunotolerance to HBV infection.
HBeAg is a key player in the establishment and persistence of chronic HBV infection through interfering with host antiviral immune responses (13,27,30). A more recent report showed that HBeAg suppressed LPS-induced NLRP3 inflammasome activation and IL-1β production by repressing NF-κB pathway and ROS production (29). To address the question whether HBeAg could also inhibit activation of the AIM2 and IFI16 inflammasomes in PBMCs of CHB patients, we performed a functional assay in vitro. The results verified that HBeAg inhibited the transcription and mature secretion of IL-1β in PBMCs transfected with poly (dA:dT) or VACA ds 70 mer, while not affecting the mRNA levels of AIM2, IFI16, and CASP1. Since HBeAg is able to attenuate phosphorylation of NF-κB and repress the activation of NF-κB signal pathway (29), we speculate that it is likely by this means HBeAg interfered with the activation of the AIM2 and IFI16 inflammasomes. The exact mechanism remains to be further studied. Taken together, these results suggest that HBeAg plays an important role in modulating expression of IL-1β and IL-18 in CHB patients.
Collectively, we examined for the first time the role of the IFI16 inflammasome in humans with acute and CHB. Our data revealed that there exists activation of the AIM2 and IFI16 inflammasomes, but not the NLRP3 inflammasome in AHB, and the activation of the AIM2 and IFI16 inflammasomes is inhibited by HBeAg in CHB, which may contribute to persistent HBV infection. These findings enhance our understanding of HBV interaction with innate immune responses. Further investigation is necessary to understand the mechanisms by which HBeAg suppresses the activation of the AIM2 and IFI16 inflammasomes.
Footnotes
Acknowledgments
This work was supported by a grant-in-aid for basic research project from the Foundation of Knowledge Innovation Program of the Committee of Innovation of Science and Technology of Shenzhen (No. JCYJ20150403101028209).
Author Disclosure Statement
No competing financial interests exist.