
Abstract
Low pathogenic avian influenza virus (AIV) infection in chickens can result in economic losses and has impacts on human health. Poultry vaccination is a tool that can be used to decrease infection and transmission of AIVs. Prior research has demonstrated that Toll-like receptor (TLR) ligands can act as vaccine adjuvants and their addition to inactivated AIV vaccines can enhance immune responses elicited in chickens. The objective of this study was to compare the adjuvant capabilities of TLR5 ligand (flagellin) and TLR21 ligand (CpG ODN 2007) administered either alone or in combination with an intramuscular formaldehyde inactivated H9N2 whole virus vaccine in chickens. Along with the inactivated virus, chickens were administered either a single dose of CpG ODN 2007 (2 or 10 μg), flagellin (0.4 or 2 μg), or a combination of both ligands. An additional group received AddaVax™, an oil emulsion style adjuvant. Chickens were vaccinated twice and serum and lachrymal samples were collected weekly following the primary vaccination, and antibody-mediated immune responses were quantified. Results showed that vaccines containing CpG ODN 2007 induce significantly greater systemic and lachrymal antibody responses than vaccines containing flagellin or AddaVax. Combinations of flagellin and CpG ODN 2007 did not demonstrate inhibitory, additive, or synergistic effects on systemic or lachrymal antibody-mediated immune responses. Additionally, for both flagellin and CpG ODN 2007, a fivefold higher dose of each did not induce significantly higher antibody-mediated immune responses compared with the lesser dose. Future studies should examine the induction of cell-mediated immune responses when flagellin, CpG ODN 2007, or other TLR ligands are administered either alone or combined as adjuvants for inactivated H9N2 AIV vaccines.
Introduction
T
H9N2 viruses isolated from chickens in China have also demonstrated intraspecies transmission in pigs and minks, highlighting mammal to mammal transmission capabilities (13,25). In addition to the risk of direct human infection with H9N2, the virus has also contributed internal genes to a triple reassortant H7N9 virus that has caused 623 human deaths since February 2013 (10,24). Different strategies exist to control H9N2 infection in poultry and vaccination is an effective mechanism to facilitate this.
Inactivated virus vaccines typically are less immunogenic than other types of vaccines, such as live attenuated viral vaccines, however, they are easier to produce and cannot revert back to a virulent form. Adjuvants are a tool that can be used to enhance the immunogenicity of inactivated virus vaccines. Toll-like receptor (TLR) ligands are a well-studied, safe, and effective group of molecules that demonstrate a broad range of adjuvant capabilities (20). TLRs are a class of pattern recognition receptors expressed in endosomes and on the surface of various cells of the immune system (1). TLRs bind unique conserved structures present in or on microbes, collectively called pathogen-associated molecular patterns. This interaction induces intracellular signaling cascades leading to increased expression and production of proteins such as cytokines and co-stimulatory molecules that are essential for rapid innate responses, and future adaptive immune responses (5,17).
Chickens express 10 different TLRs; among them are TLR5 and TLR21 (17). TLR5 in chickens recognizes bacterial flagellin and has been demonstrated to enhance systemic antibody responses as an adjuvant in chickens against a formaldehyde inactivated intramuscularly administered H4N6 vaccine (18). TLR21 in chickens recognizes synthetic oligonucleotides, including CpG ODN 2007 that has been shown to increase antibody-mediated immune responses in chickens when combined with a formaldehyde inactivated H9N2 virus vaccine (15). Both flagellin and CpG ODN 2007 have separately been shown to be effective adjuvants at a dose of 2 μg for intramuscularly administered inactivated AIV vaccines in chickens (15,18).
What remains to be elucidated is whether different doses of these ligands or their combination have an effect on the immune response elicited in chickens. Therefore, both flagellin and CpG ODN 2007 were investigated as stand-alone and combination adjuvants for a formaldehyde inactivated H9N2 virus vaccine. The results demonstrated CpG ODN 2007 to be a superior adjuvant compared to flagellin for intramuscular inactivated H9N2 virus vaccines in chickens and do not suggest that combining the adjuvants has an additive or inhibitory effect on systemic antibody responses following vaccination.
Materials and Methods
Chickens
Eighty, 1-day-old specific pathogen-free (SPF) layer chicks were purchased from the Canadian Food Inspection Agency (CFIA) (Ottawa, Canada) and were housed in the isolation facility at the Ontario Veterinary College at the University of Guelph. Sampling methods and experimental procedures were approved by the University of Guelph Animal Care Committee and were conducted with compliance to the guidelines provided by the Canadian Council on Animal Care.
Formaldehyde inactivation of H9N2 AIV
H9N2 AIV (A/Turkey/Wisconsin/1/66) was propagated in 10-day-old embryonated SPF chicken eggs. Eggs were inoculated with 4 HA units in 100 μL volume each, and incubated at 35°C for 72 h before allantoic fluid was collected, mixed with formaldehyde (final concentration 0.02%), and incubated at 37°C for 72 h while being constantly shaken. Inactivation of the virus and stability of the hemagglutinin protein was confirmed by inoculation of 10-day-old embryonated chicken eggs and hemagglutination assay, respectively. Inactivated H9N2 virus was then purified by filtering the formaldehyde treated allantoic fluid with a 40 μm cell strainer to remove debris before ultra-centrifugation at 65,000 g (SW32 Ti rotor; Optima™ L-80 XP-Beckman Coulter, Inc.) through a 20% sucrose cushion at 4°C for 2 h. The pellet was resuspended in Tris-NaCl-EDTA (TNE) buffer (50 mM Tris, 140 mM NaCl, 5 mM EDTA) and the ultra-centrifugation was repeated without sucrose to wash the inactivated virus. The concentrated virus was stored at −80°C. Total virus protein concentration was determined with a Pierce BCA protein assay kit (Thermo Scientific, Rockford, IL) following manufacturer's recommendations.
TLR ligands
Ultrapure flagellin (>95% purity) isolated from S. typhimurium and synthetic class B CpG ODN 2007 were purchased from InvivoGen (San Diego, CA). The ligands were suspended in phosphate buffered saline (PBS) before use.
Vaccine formulation and experiment timeline
Eighty SPF chickens were randomly divided into 10 groups, each consisting of 8 chickens. Chickens were vaccinated via intramuscular injection in the thigh muscle on days 7 and 21 posthatch receiving 100 and 200 μL total volume, respectively. All chickens excluding those in the PBS control group received 15 μg of formaldehyde inactivated H9N2 virus per vaccination, corresponding to ∼960 HA units. Mixed with the inactivated H9N2 virus, chickens received either a high or low dose of flagellin or CpG ODN 2007, a combination of both, or AddaVax™, a squalene-based oil in water emulsion adjuvant. Group names as they appear on all figures are presented in brackets in Table 1. Serum and lachrymal secretions were collected from all chickens weekly beginning 7 days postprimary vaccination (ppv) and ending 28 days ppv.
All groups had 8 chickens and received primary and secondary vaccinations on days 7 and 21 posthatch, respectively.
PBS, phosphate buffered saline.
Hemagglutination inhibition assay
Freshly collected blood was kept at 37°C for 1 h before being centrifuged at 5,000 g for 30 min; the top layer of serum was then collected. For the hemagglutination inhibition (HI) assay, serum samples were serially diluted in twofold manner in 96-well V-bottom plates. Twenty-five microliters (four HA units) of H9N2 AIV was then added to each well. The plate was shaken for 30 sec, covered, and incubated for 45 min at room temperature. Then, 50 μL of 0.5% chicken red blood cells (1% fetal bovine serum [FBS]) was added to each well and the plates were again shaken, covered, and incubated for 45 min at room temperature. HI titer was expressed as the reciprocal of the highest dilution to show inhibition of red blood cell agglutination (expressed as log2 scale). Rates of seroconversion and seroprotection, defined as postvaccination HI titers that increase by fourfold or greater, and HI titers >40, respectively (15), were calculated using HI titers.
Enzyme-linked immunosorbent assay
Serum and lachrymal samples were analyzed for the presence of anti-H9N2 antibodies by enzyme-linked immunosorbent assay (ELISA). Maxisorp flat bottom 96-well plates were coated with 800 ng of heat killed H9N2 AIV in 100 μL per well and incubated overnight at 4°C. Plates were washed three times with 200 μL wash buffer (0.05% Tween 20 in PBS) and then blocked with 100 μL fish skin gelatin (0.25% in wash buffer) for 1 h followed by the addition of 100 μL of diluted chicken serum or lachrymal secretions. After 1 h of incubation, plates were washed three times and 100 μL of goat anti-chicken IgY or IgM antibody conjugated to horseradish peroxidase (1:5,000) was added (Bethyl Laboratories, Montgomery, TX). One hour later plates were washed three times and peroxidase substrate (ABTS peroxidase substrate system, Kirkegaard and Perry Laboratories Gaithersburg, MD) was added. Color development was stopped after 20 min with 1% SDS and absorbance at 405 nm was determined using an ELISA plate reader (Bio-Tek Instruments, Winooski, VT). Relative serum antibody titer was calculated in relation to a serially diluted high titer serum sample that was used on every plate using methods described previously (11). For lachrymal secretions, optical density (OD) at 405 nm was measured and average OD was compared between groups.
Statistical analysis
Antibody titers from serum and lachrymal secretions were compared using R© software with Duncan's Multiple Range Test and differences in means were considered significant if p < 0.05.
Results
Seroconversion, seroprotection, and HI antibody titer
Serum was collected weekly following primary vaccination on day 7 posthatch. This resulted in four time points of serum collection, two of which occurred after the secondary vaccination, which was administered 21 days posthatch and 14 days ppv. All vaccine formulations induced significantly elevated HI antibody titers against H9N2 relative to the PBS control group at all time points postvaccination (p < 0.05) (Fig. 1A–D). Seroconversion rate was calculated for all groups on day 7 and 14 ppv.
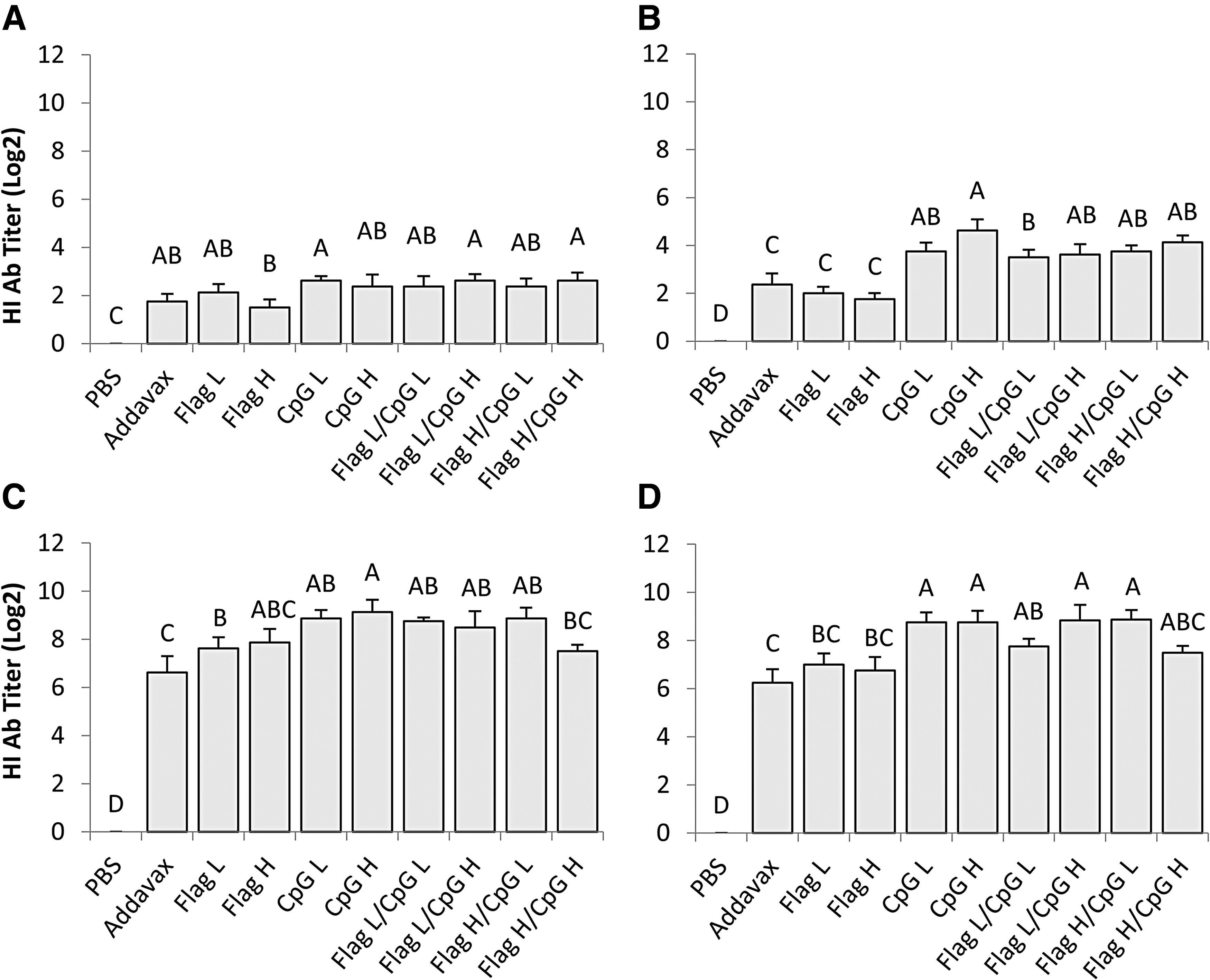
Serum HI antibody titers against H9N2 AIV. Average serum HI antibody titers of all groups from
On day 7 postvaccination, the group that received a 2 μg dose of CpG ODN 2007 in addition to one group that received a combination of adjuvants displayed 100% seroconversion, double what was observed in the group that received AddaVax (Table 2). By day 14 ppv, only the groups that received CpG ODN 2007 either alone or with flagellin demonstrated 100% seroconversion (Table 2). Seroprotection rate was also calculated for all groups on day 21 and 28 ppv (day 7 and 14 postsecondary vaccination [psv]). On both days 21 and 28 ppv, all chickens that received CpG ODN 2007 alone or in combination with flagellin demonstrated 100% seroprotection; the group that received 0.4 μg of flagellin also demonstrated 100% seroprotection on both days. The group that received the AddaVax adjuvant displayed seroprotection rates of 25% and 75% on days 21 and 28 ppv respectively (Table 2).
Chickens were vaccinated on day 7 and 21 posthatch.
ppv, postprimary vaccination.
Mean serum HI titers against H9N2 virus were also compared between groups on all four time points ppv. The groups of chickens that received CpG ODN 2007 alone (2 or 10 μg) demonstrated significantly higher HI titers than chickens receiving either dose of flagellin (0.4 or 2 μg) alone or AddaVax on day 14 and 28 ppv (p < 0.05) (Fig. 1B, D). On day 14 ppv, all groups that received CpG ODN 2007 alone or in combination with flagellin demonstrated significantly higher serum HI titers than groups that received flagellin alone (p < 0.01) or AddaVax (p < 0.05) (Fig. 1B). This trend was also observed on day 28 ppv with the exception of two groups that received different combinations of CpG ODN 2007 and flagellin.
In this study, combining adjuvants had no significant additive or synergistic effects on HI titers compared with CpG ODN 2007 alone. On day 14 and 21 ppv, separate individual groups that received a combination of TLR ligand adjuvants had significantly lower HI titers compared with the group that received a 10 μg dose of CpG ODN 2007 alone as adjuvant (p < 0.05) (Fig. 1B, C). For both flagellin and CpG ODN 2007, a fivefold difference in dose when administered alone did not result in any significant differences in mean serum HI antibody titers (Fig. 1).
Serum IgY and IgM antibody titers
ELISA was performed to gain a better understanding of the isotype makeup of serum antibody responses. On day 7 ppv the group that received a 2 μg dose of CpG ODN 2007 had significantly greater titers of IgM compared to groups that received flagellin alone or AddaVax (p < 0.05) (Fig. 2A). On day 21 ppv all groups that received CpG ODN 2007 either alone or combined with flagellin demonstrated significantly greater serum IgM titers compared with groups that received flagellin alone or AddaVax (p < 0.05) (Fig. 2C). Also, on day 14 ppv all groups that received CpG ODN 2007 alone or in combination had significantly higher titers of serum IgM antibodies than groups that received flagellin alone (p < 0.05) (Fig. 2B). Serum IgM titers peaked on day 21 ppv for all groups and by day 28 ppv IgM titers began to decrease.
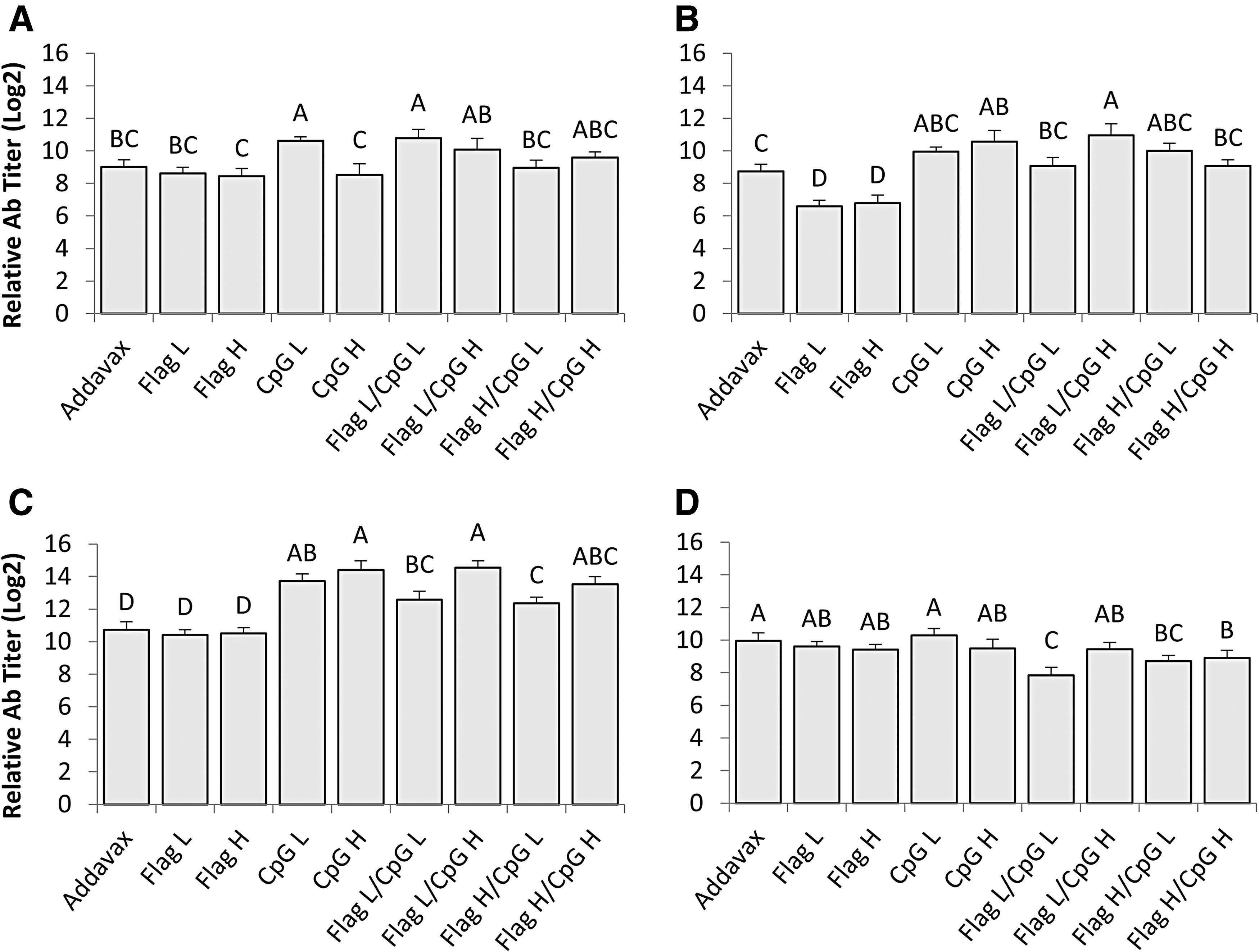
Serum IgM antibody titers against H9N2 AIV. Average serum IgM antibody titers of all groups from
Relative serum IgY titers were also determined for all groups on days 7, 14, 21, and 28 ppv (Fig. 3A–D). On days 21 and 28 ppv all groups that received CpG ODN 2007 either alone or in combination with flagellin demonstrated mean serum IgY titers, which were significantly higher than groups that received flagellin alone or AddaVax (p < 0.05) (Fig. 3C, D); a similar trend was observed on day 14 ppv as well, but IgY titers from groups that received CpG ODN 2007 were only significantly greater than the groups that received flagellin alone and not AddaVax (p < 0.05) (Fig. 3B). The combination of CpG ODN 2007 and flagellin had no consistent effects on serum IgY titers when compared to CpG ODN 2007 alone; the only significant difference noted occurred on day 28 ppv between a group that received a combination of adjuvants and the group that received a 10 μg dose of CpG ODN 2007 as adjuvant (p < 0.05) (Fig. 3D). For both CpG ODN 2007 and flagellin there were no significant differences observed in average serum IgY titers between low and high doses at any time points.
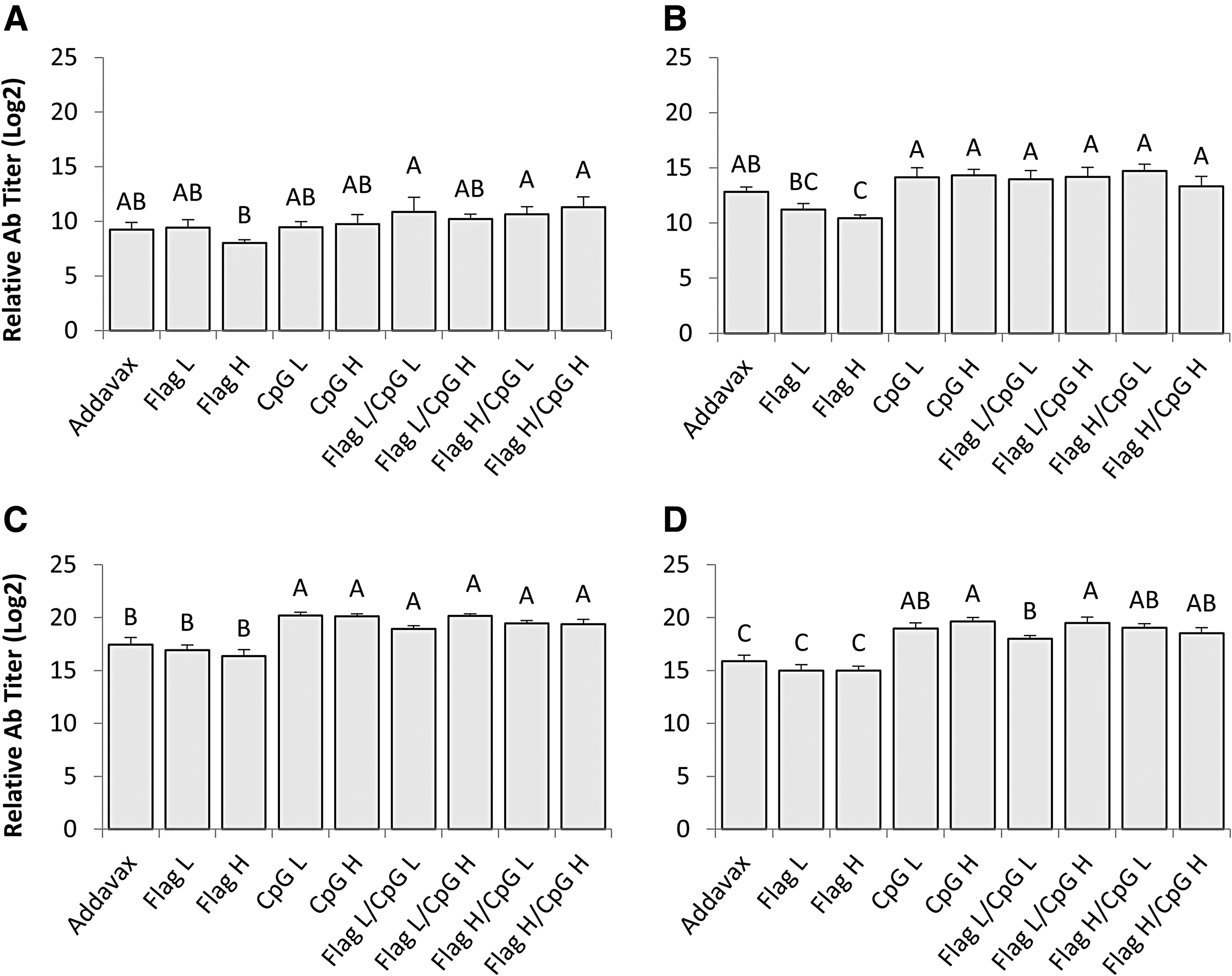
Serum IgY antibody titers against H9N2 AIV. Average serum IgY antibody titers of all groups from
IgY antibody titers in lachrymal secretions
Lachrymal secretions were collected on day 7, 14, 21, and 28 ppv and were analyzed for the quantity of IgY. On day 7 ppv, all vaccinated groups except for the AddaVax group demonstrated significantly higher levels IgY in lachrymal secretions compared with the nonvaccinated PBS group (p < 0.05) (Fig. 4A). By day 14 ppv, all groups that received CpG ODN 2007 and the AddaVax group were found to have significantly higher levels IgY in lachrymal secretions compared with groups that received flagellin alone (p < 0.05) (Fig. 4B), this trend was also seen on day 21 ppv (Fig. 4C). On day 28 ppv, there were no significant differences in amounts of lachrymal IgY between groups that received CpG ODN 2007, however, all of those groups that did receive CpG ODN 2007 demonstrated levels of lachrymal IgY that were significantly greater than those from groups that received flagellin alone (p < 0.05) (Fig. 4D). On day 28 ppv, only the group that received a high dose of CpG ODN 2007, and the group that received a combination of a high dose of flagellin and a low dose of CpG ODN 2007 demonstrated significantly greater levels of lachrymal IgY compared with the group that received AddaVax (p < 0.05) (Fig. 4D).
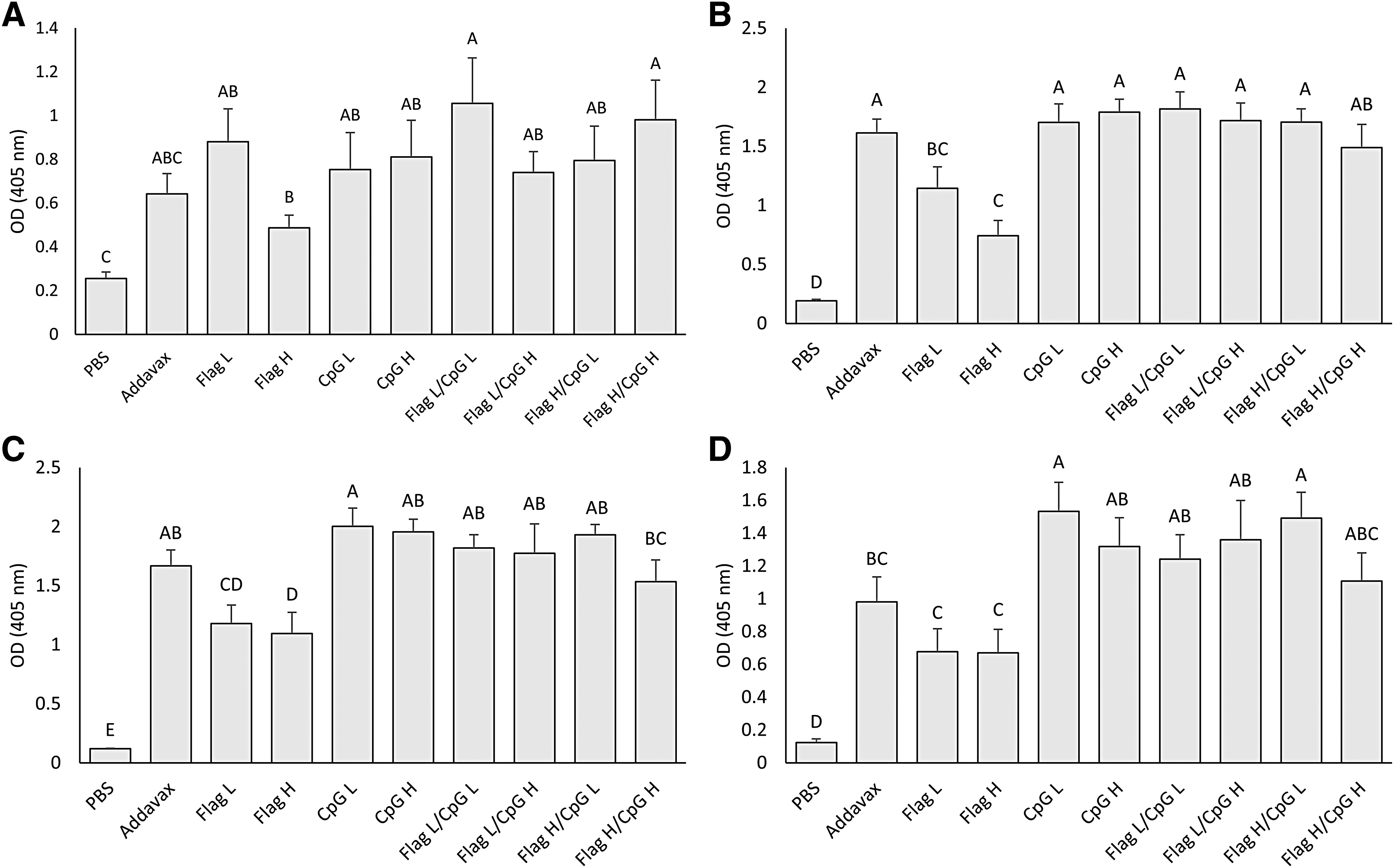
IgY antibodies against H9N2 AIV in lachrymal secretions. Average IgY antibodies in lachrymal secretions of all groups from
Discussion
In this study, the adjuvant capabilities of TLR5 and TLR21 ligands, flagellin and CpG ODN 2007 respectively, were demonstrated. Previous works have shown both of these ligands to be effective at enhancing systemic antibody-mediated immune responses when administered intramuscularly in chickens compared with vaccines containing only inactivated influenza virus (15,16,18). In addition to comparing the ligands at previously studied doses (2 μg), fivefold different doses of each ligand were also compared to determine whether antibody-mediated immune responses following vaccination would be affected.
The findings from this study suggest that CpG ODN 2007 is a superior adjuvant compared to flagellin and AddaVax when considering the induction of antibody-mediated immune responses. Antibody titers against H9N2 AIV were consistently elevated in the serum of chickens that received CpG ODN 2007 when compared with chickens that received AddaVax. Additionally, compared to flagellin, CpG ODN 2007 consistently elevated the levels of IgY antibodies in the lachrymal secretions of chickens.
Prior research has shown that a 2 μg dose of CpG ODN 2007 is not more effective than AddaVax at inducing lachrymal IgY antibodies (14). This study made similar findings, however, it should be noted that lachrymal IgY titers were significantly elevated 2 weeks psv. Additionally, CpG ODN 2007 has been shown to induce superior HI antibody responses against an inactivated H9N2 vaccine when compared to Addavax in the past (15). Herein, this finding was confirmed, and it was also demonstrated that serum IgY and IgM antibody titers were increased more by CpG ODN 2007 than AddaVax.
We also examined different doses of flagellin and CpG ODN 2007. Overall, we observed that a fivefold lower dose of both ligands was equally effective as the higher dose at inducing antibody-mediated immune responses. This is in accordance with previous results that demonstrated a 2 μg dose of CpG ODN 2007 to be more effective than a 20 μg dose at inducing antibody-mediated immune responses against an inactivated H9N2 virus vaccine in chickens (16). When comparing the ligands against each other, CpG ODN 2007 was clearly superior to flagellin at inducing serum and lachrymal antibodies against the H9N2 virus vaccine. Previous research has made similar findings regarding TLR ligand adjuvants, showing that in chickens, CpG ODN 2006, a class B CpG ODN similar to CpG ODN 2007, induced greater HI antibody titers against an inactivated H5N1 virus vaccine than a TLR2/4 (Bacillus subtilis spores) and a TLR3 (poly I:C) ligand (8).
There is evidence to suggest why CpG ODN 2007 induces increased levels of antibody-mediated immune responses in chickens. Specifically, injection of CpG ODN 2007 in chickens was shown to enhance expression of interferon (IFN)-γ and major histocompatibility complex (MHC)-II transcripts in the spleen (19). Formation of adaptive immune responses in the spleen could be enhanced by the increased expression and production of these proteins that are crucial for the development of adaptive immune responses. Additionally, previous work has shown that CpG ODN 2006 induces in vitro proliferation of human naïve B cells, however, this was not observed for flagellin (4). Hence, the ability to stimulate B cell proliferation could be a mechanism to explain why CpG ODN 2007 was able to induce significantly greater antibody responses compared with flagellin in the present study.
Another main question addressed in this study was whether combining CpG ODN 2007 and flagellin would have any effects on the induction of antibody-mediated immune responses. Flagellin is recognized by TLR5, which is present on the extracellular surface of cells, and CpG ODN 2007 is recognized by TLR21, which is present in endosomes (6). After ligand recognition, both receptors utilize the intracellular adaptor protein MyD88 to eventually activate the transcription factor NF-κB, which leads to rapid innate responses and future adaptive immune responses (17).
We hypothesized that combining these ligands would have some level of additive or possibly synergistic effect on serum and lachrymal antibody-mediated immune responses. However, the results of the study did not provide evidence for any additive, synergistic, or inhibitory effects of combining these ligands when considering antibody-mediated immune responses. This could possibly be due to the similarities in signaling pathways these ligands share once bound to their cognate receptor. However, it has been shown that in vitro co-stimulation of human peripheral blood mononuclear cells (PBMC) with flagellin and CpG leads to a synergistic increase in production of the proinflammatory cytokines, interleukin (IL)-6 and IL-1β (22), and a significant increase in production of IFN-γ (9).
Other TLR ligand combinations have been explored in vitro, for example, co-stimulation of chicken PBMCs with CpG ODN and poly I:C has been shown to synergistically increase the expression or production of certain cytokines, enzymes, and chemicals, such as nitric oxide, inducible nitric oxide synthase, and IFN-γ (2,3). These synergistic responses could be explained by differences in signaling pathways, as the receptors use separate adaptor proteins following stimulation, MyD88 for TLR21 and TRIF for TLR3. This relationship suggests that these ligands could be an effective adjuvant combination for chicken vaccines. Nevertheless, vaccination experiments in chickens using an inactivated H4N6 AIV vaccine have shown that combining CpG ODN 2007 and poly I:C as adjuvants does not lead to synergistic or additive increases in antibody-mediated immune responses compared with administering either ligand alone (16). Nevertheless, poly I:C and CpG ODN 2007 combinations should still be studied in chickens as adjuvant combinations for inactivated H9N2 AIV vaccines.
In conclusion, we have demonstrated that CpG ODN 2007 is more effective than flagellin at inducing serum and lachrymal antibody-mediated immune responses against a formaldehyde inactivated H9N2 AIV vaccine in chickens. It was also shown that a 2 μg dose could exert these effects equally compared with a 10 μg dose, and that combining CpG ODN 2007 with flagellin did not have any additive, synergistic, or inhibitory effects on antibody responses. Future experiments should examine the effects CpG ODN 2007 has on adaptive immune responses when combined with H9N2 virus vaccines inactivated using different inactivation methods.
Footnotes
Acknowledgments
We would like to thank the staff of the isolation unit at Ontario Veterinary College at the University of Guelph for their help with care and housing of chickens. This work was funded by Agriculture and Agri-Food Canada, the Ontario Ministry of Agriculture Food and Rural Affairs, Egg Farmers of Canada, Chicken Farmers of Saskatchewan, and the Canadian Poultry Research Council. This research is supported in part by the University of Guelph's Food from Thought initiative, thanks to funding from the Canada First Research Excellence Fund.
Author Disclosure Statement
No competing financial interests exist.