
Abstract
Majority of norovirus (NoV) outbreaks and sporadic infections in the past 20 years have been caused by GII.4 variants. In 2014, NoV GII.17 genotype replaced GII.4 strains in several Asian countries and major outbreaks were reported in other continents. As GII.17 is a recently evolved NoV strain, there is a gap especially in immunogenicity data. In this study, we investigated GII.17 virus-like particle (VLP) binding to various cellular ligands, histo-blood group antigens (HBGAs), using human saliva, pig gastric mucin, and synthetic oligosaccharides as HBGA sources. The level of GII.17 immunological cross-reactivity was determined in mice immunized with different monovalent and multivalent NoV VLP compositions. Furthermore, healthy adult volunteers with natural NoV exposure history were analyzed for GII.17-specific seroresponses. The results showed that GII.17 Kawasaki VLPs bind to a wide range of HBGAs, even though fewer than GII.4 VLPs. Immunization of mice with a multivalent VLP formulation containing different genogroup II NoV VLPs was required to obtain the highest magnitude of cross-reactive binding antibodies to GII.17. However, coimmunization with different VLP genotypes did not improve potentially neutralizing antibodies to GII.17, which remained very moderate. Low pre-existing cross-reactive antibodies to GII.17 observed in human adults indicate the presence of a large pool of susceptible individuals in this population. These data suggest that GII.17 and alike newly emerging distinct NoV genotypes should be considered as an antigenic component for vaccine formulations, which currently widely rely on GII.4-specific immunity.
Introduction
N
For the last two decades, NoVs belonging to the GII.4 lineage have caused globally most of the outbreaks and at least six pandemics (14). Exceptionally, in 2014–2015 winter season, there was a rapid increase in outbreaks caused by the novel GII.17 Kawasaki variant in several East Asian countries, including China, Japan, and Hong Kong, surpassing GII.4 Sydney as the predominant strain (8,11). At the same time, GII.17 epidemics were reported in other areas such as in the United States and Europe (7,40,50). Emergence of new variants may result in larger outbreaks and even pandemics, when population lacking pre-existing immunity is exposed to antigenically distant viruses (13,49).
NoV-specific antibodies that block binding of virus-like particles (VLPs) to the histo-blood group antigens (HBGAs) are considered the best correlate of protection against NoV (5,34,38,42). In general, there is no cross-protection induced between different NoV genogroups, and despite the possible cross-reactivity within the genogroup, the level of protection may be low due to highly specific nature of blocking (neutralizing) antibodies. Our group and others have previously demonstrated that sequential heterologous NoV infections occur frequently, especially in children, due to limited protective immune responses between different NoVs (6,8,41).
The expression pattern of HBGAs on human gastrointestinal cells, affected by the ABO blood type, secretor, and Lewis (Le) genotypes of an individual, is important in NoV susceptibility since they function as attachment factors/receptors for cellular entry and infection (16,18). Comparison of novel GII.17 Kawasaki strain with older GII.17 variants has revealed residue substitutions in HBGA-binding interfaces and in antigenic epitopes, which probably have introduced a wider HBGA binding spectrum for GII.17 (8,36,43,52). Approximately 20% of the population have a nonsecretor phenotype that do not present ABO blood group or Leb/y antigens. Typically, nonsecretor status protects from infection with several NoV genotypes (37), but novel GII.17 NoV, as well as other strains such as GI.3 and GII.3, has been shown to be infectious also to some nonsecretor (Lea/x positive) individuals (37,47,52). These changes that improve GII.17 viral fitness have raised concerns of the global spread of the novel GII.17 strain.
In this study, we assessed the binding of GII.17 Kawasaki-2015 VLPs to various HBGAs from different sources, in relation to ancestral pandemic GII.4-1999 (GII.4-99) or phylogenetically more closely related GII.12 NoV VLPs. Using naive BALB/c mice, we determined the level of cross-reactive binding and blocking antibodies induced against GII.17 after immunization with two different GII.4 variant (GII.4-99 and GII.4 New Orleans-2009) VLPs or GII.12 VLPs. Furthermore, we investigated GII.17-specific seroresponses in healthy adult volunteers. Current vaccine approaches consider GII.4 as a main NoV vaccine antigen (9,29), and, therefore, it is of interest to determine the level of GII.4 VLP-induced cross-reactive responses to GII.17.
Materials and Methods
VLP production and characterization
Five NoV VLPs, including GI.1 (2001, Genbank accession number: AY502016), GI.3 (2002, reference strain acc. no.: AF414403), GII.4-99 (1999, ref. strain acc. no.: AF080551), GII.4 New Orleans (NO) (2010, ref. strain acc. no.: GU445325), and GII.12 (1998, ref. strain acc. no.: AJ277618), were produced in Sf9 insect cells using Bac-to-Bac baculovirus expression system (Cat. 10359-016; Invitrogen). VLPs were purified by sucrose-gradient ultracentrifugation and analyzed for total protein concentration, purity, integrity, and endotoxin content (<0.1 EU/10 μg of protein), and transmission electron microscopy confirmed VLP morphology as previously described by our laboratory (4,17,44). GII.17 Kawasaki 308 VLPs (2015, acc. no.: BAR42289) used in analytical methods only were produced by Icon Genetics GmbH (Halle, Germany) (15).
Alignment and phylogenetic analysis
Major capsid VP1 aa sequences of the six NoV VLPs used in this study and sequence of the most recent pandemic strain GII.4 Sydney (2012, Genbank acc. no.: AFV08795.1) were aligned using Basic Local Alignment Search Tool for sequence identity. Evolutionary analyses were conducted in MEGA X (22) using ClustalW alignment of the aa sequences. The evolutionary distances were computed using the Poisson correction method (53) and are in the units of the number of aa substitutions per site.
Generation of mouse antiserum
Seven-week-old female BALB/c (H-2d) mice (Envigo RMS BV, Horst, The Netherlands) were immunized twice with 10 μg of each monovalent NoV VLPs (GII.4-99, GII.12, GII.4 NO, GI.3, GI.1) through intramuscular route with 3 weeks interval (Table 1). Other groups of mice received bivalent combination of GI.3 and GII.4–99 VLPs (10 μg of each per injection), or trivalent GII-specific mixture of VLPs (20 μg of each GII.4–99, GII.4 NO, and GII.12 per injection, “GII-mix”), or the mixture of five genotype VLPs (GII-mix combined with GI.1 and GI.3, 20 μg each, i.e., “GI + GII-mix”). Negative control mice received 50 μL of carrier (phosphate-buffered saline, PBS) only. No adjuvants were used. At least five mice per each experimental group were included. Serum samples were collected at termination 2 weeks after the second immunization. Mice welfare was monitored throughout the study and experiments were authorized and performed in accordance with the guidelines of the Finnish National Animal Experiment Board.
NO, New Orleans; NoV, norovirus; VLP, virus-like particle.
Collection of human plasma
Venous blood samples and saliva samples of 10 healthy adult volunteers (laboratory personnel, age range 26–56 years) were collected in the spring of 2016; plasma was separated and stored at −20°C as previously described (32). Human saliva samples were typed for the presence of A, B, and O antigens by monoclonal antibodies (Immucor, Houston, TX) as described by others (16). Informed consent was obtained from each volunteer before the sample collection in accordance with the Declaration of Helsinki.
HBGA carbohydrate binding assay
GII.17 VLP binding was analyzed using three different sources of HBGAs: human saliva, pig gastric mucin (PGM) type III (Sigma Chemicals, St Louis, MO) (23), and synthetic HBGAs. Binding assays were performed using procedures described in detail elsewhere (30,48). In brief, 96-well half-area polystyrene plates (Costar, Corning, NY) were coated with either 2.5 μg/mL PGM or 1:3,000 diluted types A, B, and O saliva from adult volunteers with previously determined ABO phenotype (48). For synthetic HBGA assay, 2.5 μg/mL of the biotinylated Lea-PAA-Biotin, Leb-PAA-Biotin, Led (H type 1)-PAA-Biotin, H (type 3)-PAA-Biotin, Blood A (trisaccharide)-PAA-Biotin, or Blood B (trisaccharide)-PAA-Biotin (all from Glycotech, Gaithersburg, MD) was coated on SuperBlock pretreated High Binding Capacity NeutrAvidin plates (Pierce, Rockford, IL).
Serially twofold diluted GII.17 VLPs (1.6–0.2 μg/mL) were incubated on plate for 1 h and bound VLPs were detected using rabbit polyclonal anti-NoV immunoglobulin G (IgG) (ICON Genetics87 GmbH) and goat antirabbit IgG-horseradish peroxidase (HRP) (Abcam, Cambridge, MA). Blank wells were incubated with buffer lacking VLPs. Substrate SIGMAFAST™ OPD was read at λ = 490 nm using the Victor2 1420 Multilabel Counter (Wallac; Perkin Elmer, Waltham, MA) plate reader and results are expressed as the average optical density (OD) of duplicate wells after background (wells lacking VLPs) subtraction. Positive reactivity is defined as a mean OD >0.1 after background subtraction.
IgG antibody assays
GII.17-specific IgG antibodies in immunized mouse serum and human plasma samples were detected by enzyme-linked immunosorbent assay (ELISA) as described in detail earlier (4,34,44). In brief, mouse serum or human plasma samples were added as twofold dilution series starting at 1:100 dilution on GII.17 VLPs (at 1 μg/mL) coated 96-well half-area polystyrene plates. Bound IgG antibodies were detected either with antimouse (Sigma-Aldrich, Saint Louis, MO) or with antihuman (Novex® by Thermo Fisher Scientific) HRP-conjugated IgG. Absorbance (OD) of the o-phenylenediamine dihydrochloride (OPD) substrate (Sigma-Aldrich) was measured at 490 nm in a microplate reader. End point titers, defined as the highest serum dilution giving an OD reading more than the set cutoff value (negative control mean OD +3 × standard deviation and >0.1 OD), were determined for each sample.
Serum antibody blocking assay
Blocking of GII.17 VLP binding to HBGA carbohydrates was analyzed by blocking assay based on human saliva type A HBGAs, widely used in our laboratory (5,45,48). In brief, twofold diluted group-wise pooled mouse sera (starting dilution 1:10) or individual human plasma samples (starting dilution 1:25) were preincubated 1 h with GII.17 VLPs (0.2 μg/mL) before plating on blocked human saliva A-coated 96-well polystyrene plates. After 1.5 h incubation period on plate, bound VLPs were detected as already described for binding assay, using rabbit polyclonal anti-NoV IgG followed by goat antirabbit IgG-HRP and OPD substrate.
In addition, the analysis of mouse serum blocking activity was verified by using the PGM-based blocking assay using the same reagents as described for saliva A assay for detection. Maximum binding signal (OD) was obtained with VLPs lacking mouse serum. The blocking index was calculated as 100%− [OD (wells with serum)/OD (wells without serum, maximum binding)] × 100%. Results are expressed as the mean blocking index of duplicate wells with similar results from minimum of two independent experiments. Blocking titer 50 (BT50) was determined as the reciprocal of the final plasma dilution that blocked at least 50% of the maximum VLP binding to the HBGA.
Statistical analysis
Mann–Whitney U test was used for evaluating the significance of the cross-reactive IgG values between the groups. The analysis of statistical difference in human seroresponses against different NoV genotypes was performed using Wilcoxon Signed Ranks test for related samples. For correlation analyses, Spearman's rank correlation was used. Statistical analyses were performed using IBM SPSS Statistics (SPSS, Chicago, IL) version 25.0. Statistical significance was defined as p-value of <0.05.
Results
GII.17 is distant from pandemic GII.4 NoVs
A phylogenetic tree was constructed from the alignment of the VP1 aa sequences of GII.17, GII.12, GII.4–99, GII.4 NO, GII.4 Sydney, GI.1, and GI.3 to show the phylogenetic relationship of different NoV genotypes (Fig. 1). Within the phylogenetic analysis, recent pandemic strains GII.4 NO (2010) and GII.4 Sydney (2012) showed the most conserved aa distance values and, therefore, the closest relatedness, ancestral GII.4–99 being in proximity in the same cluster. GII.17 Kawasaki 308 is a distant genotype to GII.4 NoVs, representing another GII lineage. GII.17 and GII.4–99 capsid showed 66% aa sequence identity. To the contrary, GII.12 is more closely related to GII.17, having 75% aa identity. GI.1 and GI.3 belong to GI genogroup and are part of a separate evolutionary lineage.

Phylogenetic analysis of seven NoV major capsid protein VP1 amino acid sequences. The tree was inferred by neighbor-joining method with a bootstrap of 1,000 by using MegaX. The scale bar shows the genetic distance, expressed as amino acid substitutions per site. NoV, norovirus.
GII.17 VLP binds to several HBGAs
NoV GII.17 VLPs were assessed for their ability to bind to cellular ligands, HBGAs present in human saliva (types A, B, O, and nonsecretor), commercially available PGM, and synthetic HBGAs including types A and B trimers, H type 1, H type 3, Lea, and Leb (Fig. 2, Table 2). Strong binding of GII.17 VLPs to HBGAs in saliva samples of types A and B was observed. GII.17 VLPs binding to type O saliva were somewhat lower, whereas no binding to saliva samples of nonsecretor donors was detected above the cutoff (OD 0.1) (Fig. 2). GII.17 VLPs bound efficiently to PGM, containing H, A, and Ley antigens, and to the synthetic B trimer, whereas binding to A trimer or Lea HBGAs was low. GII.17 completely lacked binding to synthetic HBGAs H type 1, H type 3, and Leb.
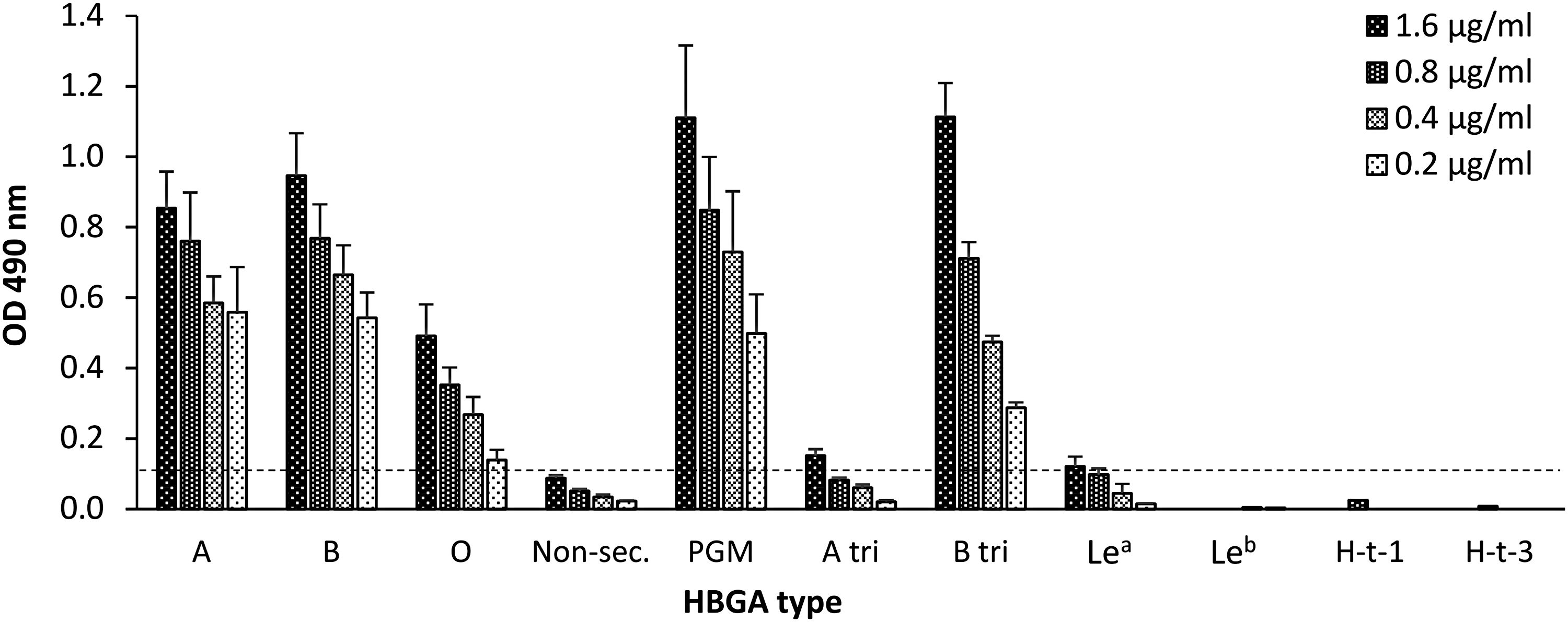
Binding profile of GII.17 VLPs to HBGAs. GII.17 VLP binding to different HBGAs was determined in binding ELISA using series of twofold dilutions of VLPs. Shown are mean OD values with standard errors of VLP binding to up to four different secretor positive saliva A, B, or O and two nonsecretors (Nonsec) saliva. Binding to PGM and synthetic HBGAs (Le, Lewis antigen) are mean ODs of repeated analysis of duplicate samples. Horizontal line indicates a positivity cutoff. ELISA, enzyme-linked immunosorbent assay; HBGAs, histo-blood group antigens; OD, optical density; PGM, pig gastric mucin; VLP, virus-like particle.
Data extrapolated from Uusi-Kerttula et al., 2014 (48).
HBGAs, histo-blood group antigens; Le, Lewis; PGM, pig gastric mucin.
Heterologous immune mouse sera cross-react with GII.17
Serum IgG of mice immunized with monovalent VLPs or bi-, tri-, and pentavalent genotype mixtures of NoV VLPs was analyzed for GII.17-specific cross-reactivity in ELISA. All mice immunized with GII but not GI VLPs showed cross-reactive IgG response to GII.17 (Fig. 3A, B). Mice immunized with GII.4–99 and GII.12 VLPs showed the highest cross-reactive responses against GII.17, whereas GII.4 NO immunized mice had significantly lower anti-GII.17 cross-reactivity (p < 0.05). Immunization of the mice with the mixture of three (GII-mix) or five (GI+GII-mix) genotype VLPs showed additive effect of cross-reactive serological immune responses induced by each GII-derived monovalent VLP immunization, hence the significantly different response was observed (p < 0.05). There was no mutual interference or inhibition observed when several genotypes were administrated simultaneously.
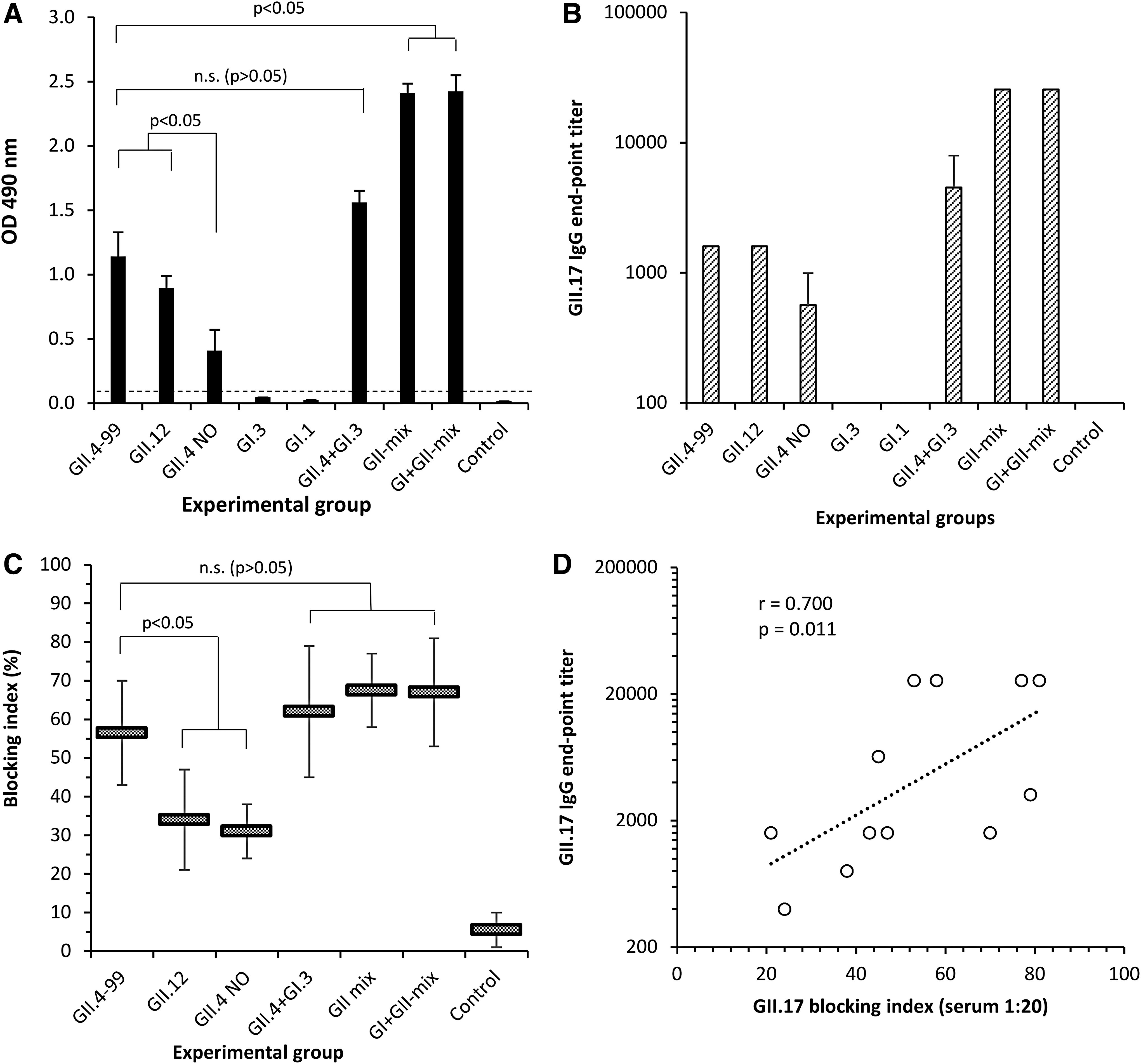
GII.17-specific IgG responses in heterologous mouse immune sera. Mice immunized twice with monovalent NoV VLPs or mixtures of two, three (GII-mix), or five (GI+GII-mix) genotype VLPs (Table 1) were analyzed for cross-reactive GII.17 responses. Shown are mean OD of 1:100 diluted individual mice sera with standard error of the means (SEMs), horizontal dashed line indicates a positivity cutoff (OD >0.100)
Heterologous immune mouse sera moderately block GII.17 VLP binding to the receptors
The groups of animals with anti-GII.17 cross-reactive IgG antibodies in immune sera were further analyzed for blocking of GII.17 HBGA binding. Pooled sera of each test group and the negative control group were tested first in saliva HBGA-based assay (Fig. 3C) and results further confirmed using the PGM-based assay (data not shown). GII.17 cross-blocking activity was generally low, blocking index at 1:20 serum dilution was <80% in all groups (Fig. 3C). The sera of mice immunized with GII.4–99 VLPs alone blocked the GII.17 VLP binding significantly better (p < 0.05) than GII.12- and GII.4 NO-specific immune sera (Fig. 3C). In contrast to the GII.17-specific cross-reactive serum IgG responses already described, no additive increase in the blocking was observed when VLPs were given as a mixture (p > 0.05). Overall, a positive correlation was detected between the serum GII.17-specific IgG end point titers and the blocking indices (r = 0.971, p = 0.001) (Fig. 3D).
Low levels of pre-existing antibodies to GII.17 in adult human sera
The IgG titers against GII.17, GII.12, and GII.4–99 were measured from plasma samples of adult volunteers having different blood types and secretor status (Table 3). GII.4-specific end point titers were significantly higher (geometric mean titer, GMT 10,397) than GII.17-specific (GMT 4,222) or GII.12-specific (GMT 6,859) IgG levels (all p < 0.05). In all seven donors with considerable NoV-specific antibody levels [end point titer >12,800, (34)], the highest IgG titers were GII.4-99 specific, followed by GII.12-specific titers, whereas all GII.17-specific end point titers were ≤12,800.
Sec. type, secretory type.
BT50 titers with 50% blocking of NoV GII.17 VLP binding. BT50 ≥200 indicates protection from NoV infection and disease (1).
BT50 of 12.5 is an arbitrary titer assigned to samples lacking blocking at a serum dilution 1:25.
GMT, geometric mean titers.
Correlation between the GII.17, GII.4–99, and GII.12-specific antibody levels was assessed using the Spearman rank correlation coefficient (Fig. 4A, B). No significant correlation was found between GII.17 and GII.4–99 IgG end point titers (r = 0.410, p = 0.239) (Fig. 4A) or between GII.17 and GII.12 IgG end point titers (r = 0.528, p = 0.117) (Fig. 4B). In addition, the ability of plasma samples to block GII.17 VLP binding to human saliva type A HBGAs was assessed for each volunteer. GII.17 blocking titers were quite low in each adult volunteer, BT50 being ≤100 (Table 3). As expected, a significant positive correlation (r = 0.771, p = 0.009) was observed between GII.17 IgG titers and GII.17 BT50 in plasma (Fig. 4C).

Correlation of GII.17-, GII.4-99-, and GII.12-specific NoV IgG seroresponses of human donors. Plasma samples of 10 adult donors were titrated to determine genotype-specific end point titers. Correlation between anti-GII.17 IgG end point titers with anti-GII.4-99
Discussion
Since the novel GII.17 NoV variant emerged and surpassed the predominating GII.4 variants in East Asian countries in 2014, there has been a concern of global spreading of GII.17 NoV. The risk related to GII.17 NoV global epidemic can be evaluated when the susceptible host range, the level of pre-existing GII.17-specific immunity, and possible GII.17 cross-reactivity with other circulating strains is known. Moreover, this information is important when predicting the essential genotype compositions for NoV vaccine development, currently relying much on predominant GII.4 strains (9). This study aimed to gather information on these critical aspects by analyzing the HBGA binding spectrum of GII.17, the cross-reactivity induced in naive mice immunized with distinct NoV genotype VLPs, and by measuring pre-existing GII.17-specific immunity in adult human volunteers.
We have previously published an extensive study on HBGA recognition of six different NoV genotype VLPs and shown the high degree of variability in ligand binding range depending on the genotype (48). Here the work extends to determine the HBGA binding spectrum of GII.17 VLPs. Overall, when compared with very broad binding profile of GII.4-specific VLPs (48), the GII.17 binding range is more limited and lacks binding to several synthetic HBGAs, such as the biologically relevant H type 1 (35). However, GII.17 VLPs seem to bind more efficiently to different HBGA types than, for example, GII.12, GI.3, or GI.1 VLPs tested previously (48). Furthermore, concordantly with the previous findings (19,52) and resembling pandemic GII.4 variants (48), GII.17 was shown to bind well HBGAs present in types A, B, and O salivas. High binding to PGM, containing A, H, and Ley HBGAs (23), was observed, supporting previous reports (26). Very weak binding to Lea was observed in this study, probably explaining the reported low GII.17 infection rate of nonsecretors through Lea/x recognition (19,52).
In general, the broader binding spectrum to different HBGAs may be associated with efficient spreading of the genotype, due to the larger pool of susceptible population (46). Concomitantly, pandemic GII.4 strains have been shown to possess extremely versatile HBGA binding, whereas non-GII.4 strains show modest selection of binding HBGAs (48). In this regard, the risk of global pandemic with GII.17 Kawasaki can be considered less likely than that of GII.4 variants. Before recently emerged GII.17 variants, GII.17 strains were considered as slowly evolving NoVs, compared with GII.4 viruses with rapid VP1 mutation rate (41). However, new GII.17 variants have been reported to evolve even faster than GII.4 (8).
As GII.17 is one of the most recent emerging NoV variants, the immunogenicity data on GII.17 cross-reactivity with other NoV genotypes are still limited (10). In this study, the cross-reactivity between GII.17 and several other NoV genotypes was evaluated by using naive inbred Balb/c mice immunized with monovalent, bivalent, or multivalent combination of different NoV VLPs. There was no GII.17-specific intergenogroup response detected in mice immunized with GI VLPs, but there was an additive effect to GII.17-specific immunity when GII VLPs were combined. However, even though GII.17 cross-reactive IgG titers were increased by adding several VLPs in a mixture, GII.17 cross-blocking activity was not significantly improved. This is understandable considering the high genotype specificity of blocking antibodies, as observed earlier by us and others (23,27,34,42).
Antigenically, GII.12 is the closest genotype to GII.17 used in this study, VP1 protein sequences of GII.17 and GII.12 VLPs share 75% identity, whereas more distant GII.4–99 shares 66% of the aa sequence with GII.17. Considering the degree of genetic identity (Fig. 1), we anticipated GII.12 to be the most potent inducer of GII.17 cross-reactive and cross-protective responses in mice. However, after the monovalent VLP immunization, the highest anti-GII.17 IgG cross-reactivity was observed in GII.4–99 immune sera, whereas modest GII.17 cross-reactivity was induced by the recent pandemic strain GII.4 NO. Furthermore, higher cross-blocking of GII.17 by GII.4–99 mouse immune sera than by GII.12 mouse immune sera was detected.
Congruently, in our previous studies comparing the immunogenicity of six different NoV genotypes, we observed that GII.4–99 VLP immunized mice generated the broadest cross-reactive antibody responses against both close and more distant NoV genotypes (33). The mechanism behind this finding is unknown but it may be due to stronger affinity of antibodies that GII.4–99 VLPs induce (12,28,31,33,39). Furthermore, there are reports of conserved neutralization antibody epitopes located outside the HBGA binding pocket, for example, blocking epitope F with “NERK” motif (24), which may be less prone to sequence variations than HBGA binding sites and still block NoV binding to HBGA through allosteric interference. Moreover, two residues of “NERK” have been linked to GII.10 NoV blocking mechanism, suggesting possible involvement of this motif on NoV neutralization on larger scale (20).
However, comparison of five putative GII.4 blockade antibody epitope sequences (12,25) (Table 4) did not provide more insights into superiority of GII.4–99 to induce GII.17 blocking antibodies, as GII.17 shared 12 aa residues with genetically more closely related GII.12 and only 6 aa residues with GII.4–99. In this respect, so far unidentified conserved epitopes and aa residues may play a significant role in blocking of NoV HBGA binding.
Amino acids shared between two genotypes are in bold.
The serological analysis of human adult responses to GII.17, GII.12, and GII.4–99 NoVs revealed that the highest antibody responses were developed against ancestral GII.4–99, whereas pre-existing immunity to less frequently encountered GII.12 and GII.17 was low. Even though the number of volunteers in this study was limited, the data showing low pre-existing GII.17 immunity are consistent with the data others have reported (8,10). Using the epidemiological pediatric samples in our previous studies (34,38), NoV genotype-specific IgG end point titer >12,800 was defined as protective. In this study, there were no protective GII.17 titers detected in the samples collected in the beginning of 2016.
When comparing the levels of GII.4-, GII.12-, and GII.17-specific antibodies, the best responses were targeted to GII.4–99, an ancestor for GII.4 strains dominating for more than two decades. No correlation between GII.4–99 and GII.17 antibody levels was found, congruently to what was reported by Dai et al. (10). Surprisingly, no positive correlation was detected between GII.17- and GII.12-specific IgG titers, although these genotypes are genetically and antigenically more related (Fig. 1).
HBGA-blocking antibodies, in particular BT50 ≥ 200, were associated with protection to NoV infection and disease (1,2,34). According to this, none of the tested adults in this study would be protected from upcoming GII.17 infection. Consistent with our results, study with GII.17- and GII.4-infected patients has shown limited cross-reactivity and the lack of cross-blocking between these genotypes (10). The lack of pre-existing GII.17-specific herd immunity generates the pool of susceptible individuals and supports the rapid emergence of GII.17 in a population (8).
Our data suggest that recent increasing prevalence of GII.17 NoV may be explained by broad selection of binding HBGAs, low cross-reactivity with other circulating NoV strains, and low pre-existing immunity in human population. The results in mice show for the first time that even a vaccine formulation mixture of up to five heterologous VLPs does not induce significant protective response to GII.17 genotype. Regarding NoV vaccine design, our results suggest that in addition to NoV GII.4 VLPs, the optimal vaccine design would need to consider newly emerging or predicted circulating NoV genotypes.
Footnotes
Acknowledgments
We are grateful to Icon Genetics GmbH for providing magnICON(R)-produced GII.17 VLPs. The personnel of Vaccines Research Center of University of Tampere are thanked for technical assistance.
Author Disclosure Statement
No competing financial interests exist.