
Abstract
Respiratory syncytial virus (RSV) is a very frequent viral respiratory pathogen of the young (<5 years old) with a significant portion of young toddlers having been infected before 2 years of age. Although we understand that some of the morbidity associated with RSV in neonates is due to immunological maturation that favors immunosuppression over antiviral innate and/or adaptive immune responses, the rapid development of the immune system right after birth suggests that each age group (newborn, early infant, older infant, toddler, and older) may respond to the virus in different ways. In this study, we summarize the morbidity associated with infection in young children in the context of immunological maturation of monocytes/macrophages and the ramifications for poor innate control of viral pathogenesis. We also summarize key mechanisms that contribute to the diminished antiviral innate immune responses of these young children.
Introduction
Respiratory syncytial virus (RSV) is the primary cause of neonatal (birth–1 month) and infantile (2–12 months old) bronchitis and is the most common cause of lower respiratory tract infections in young children (49,58). Furthermore, the virus is also commonly associated with secondary bacterial infections in the middle ear or pneumonia (62,85,88) in infants and toddlers (12–36 months old). Each year, RSV infection results in ∼120,000 hospitalizations among children <5 years of age in the United States and 160,000 yearly deaths worldwide (16). RSV infection is about equally likely to cause infant death whether in a developed or a developing country (68).
Although the association between RSV and lower respiratory disease is known among neonates, in infants (2–24 months old), and toddlers (24–36 months old), the mechanism(s) causing increased morbidity during these infections (without compounding factors such as secondary bacterial infections) remain poorly defined (79). However, prior studies have shown possible prolonged viral replication in the lung and limited immune responses against RSV in infants suggesting diminished immune responses could be an issue in young children (90,91). Risk factors for enhanced RSV morbidity include low birth weight, premature birth, immunodeficiency, and congenital lung or heart disease, but often those hospitalized do not have these compounding cofactors (6,19).
Treatment for RSV is limited with alleviation of the symptoms of infection rather than the underlying cause more common (20). However, in the premature neonate, monoclonal antibody palivizumab therapy has been used, but the cost, true efficacy of the therapy, and frequency of dosing has limited its use in other age groups, including toddlers (32).
Neonates rely heavily on maternal antibodies for protection from RSV and secondarily on their innate immune defenses as their adaptive immunity is slowly developing and becoming more proinflammatory with age (55). However, maternal-derived RSV-specific antibodies have an estimated half-life of 1.5 months (55). This protection is, therefore, diminished in older infants or toddlers, which may be a contributing factor to their risk of RSV infection. Since immunity to RSV can wane within the same viral season or to the next (14,71), prior exposure to the virus may not protect the older infant or toddler from infection (71,77) although protection from severe disease may still occur. However, immune memory to RSV G in the older child or adult may play some role in the reduced incidence of RSV hospitalizations (18). Therefore, limitations in the innate immune system to suppress early viral replication could be a key mechanism for enhanced morbidity in these age groups.
As shown in Figure 1, key limitations have been identified in neonates that may contribute to RSV-mediated pathology, but less is known about mechanisms in the older infant and toddler. In this study, we summarize key contributions to RSV suppression by monocytes/macrophages and how immunological maturation may limit the full capacity of the innate immune response from better suppressing this virus.
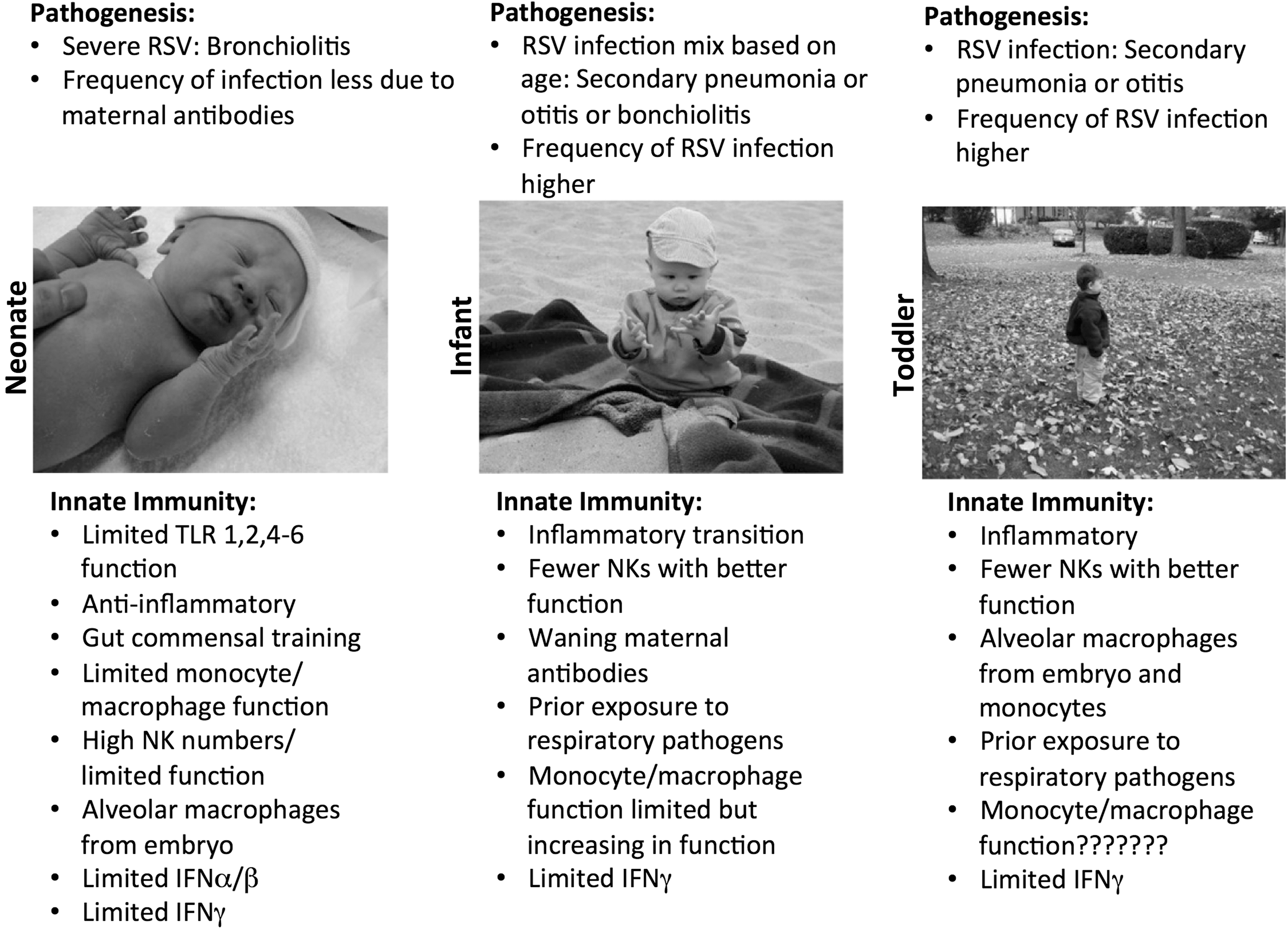
Summary of RSV pathogenesis and innate immunological maturation in young children. RSV, respiratory syncytial virus. Photos used with permission.
Monocyte/Macrophages During RSV
The importance of the mononuclear phagocyte system in early control of respiratory viral infection, including RSV, has been well demonstrated in prior studies (27,44,81). Within this system, macrophages and extravascular monocytes and dendritic cells each play a role in lung antigen surveillance, potentially facilitating innate suppression of respiratory viruses, and also helping to ensure activation and recruitment of later adaptive immune cells. Resident macrophages (e.g., alveolar or interstitial) present within the lungs are important for various functions, including inflammatory antipathogen responses or tissue repair (37,65).
Alveolar macrophages are the most abundant immune cell in the alveolar sacs and are elevated in numbers in the young child (28). Importantly, alveolar macrophages play an important role in early responses to respiratory pathogens and establishing immunity through cytokine production, recruitment of lymphocytes, antimicrobial activity, and tissue repair (15,52,59). They can be resident, arising during embryogenesis, or inflammatory, arising from monocyte recruitment to the lungs during infection (17,51). These cells are known to be limited in neonates and toddlers, which could possibly affect anti-RSV innate responses (28).
Interstitial macrophages (IMs) exist as three subsets in uninfected lungs (23). Of interest, dichloromethylene-diphosphonate treatment (liposomal depletion) not only significantly depletes alveolar, but not IMs, but also increases lung pathology during RSV infection suggesting the former may be more important to anti-RSV immunity than the latter (2). In addition, despite increased toll-like receptor (TLR) 3/7 expressions in IMs during infection, limited interleukin (IL)-33 is secreted from these cells upon infection unlike alveolar macrophages (60). Thus, the contributions of IMs to innate immunity to RSV or tissue damage are not well known, but they may be more important for phagocytizing RSV antigens for presentation to later infiltrating adaptive immune cells than having direct innate anti-RSV functions.
Although location can impart differences in macrophage activity/function, polarization can also dramatically affect function as well. Type 1 macrophages (classical activation, induced mainly through interferon gamma: IFNγ) express proinflammatory cytokines, display strong phagocytic activity, and contribute to the acute inflammatory response, resulting in clearance of pathogens. Type 2 macrophages [alternative activation, mainly through IL-4 or IL-13 (26)] express anti-inflammatory cytokines, contribute to maintenance of chronic inflammation, and are less efficient at clearing pathogens (8,74). Alternatively activated macrophages secrete IL-10, which produces arginase-1 that competes with inducible nitric oxide synthase, limiting their potential for killing intracellular pathogens (7,89). Alternatively activated macrophages are known to limit RSV-induced lung damage (73). Although potentially reducing lung damage in the neonate, any type 2 skew in lung cytokine milieus [macrophage bias toward Th2 cytokines (43)] may result in decreased production of proinflammatory cytokines (IL-1β, IL-6, IL-12, and TNFα) by innate cells that reduce the ability of the innate immune system to suppress viral replication. RSV-infected infants may be prone to Th2-type cytokine bias due to the absence of sufficiently strong polarization of T cells toward Th1 response (40). However, not all neonates or infants may be globally predisposed toward Th2 cytokines thus capable of Th1 type cytokine responses (29).
Extravascular monocytes maintain minimal differentiation in uninfected lungs and transport antigens back to the draining lymph nodes, which differ significantly during infection when inflammatory monocytes are recruited and begin to differentiate into inflammatory macrophages (35). Similar to alveolar macrophages, inflammatory monocytes may significantly not only contribute to the inflammation common during RSV infection but also contain viral spread and help initiate the adaptive response (78). However, along with protection afforded by both of these cells, RSV may also infect them and contribute to pathogenesis by prolonging viral replication within the lungs (36,61,69) through possible cell death or disruption to antiviral function.
From neonatal studies, we know that monocytes from preterm infants (highly susceptible to RSV) have significant functional impairments such as diminished IL-6 production (34) or TNFα (87) upon stimulation or diminished HLA-DR expression (67). Even term-born neonates may still exhibit less functional monocytes than adults (54,66). Differences in older infant or toddler monocytes and adults have not been well studied, and impairments that continue into this age or are unique to them could have dramatic effects on anti-RSV innate responses.
Immunological Maturation Effects on Innate Immunity in the Young
The immune system undergoes anatomic, quantifiable, and functional changes in cell fates through development and often is influenced by the developing commensal intestinal flora (10,25). Starting at 4–6 weeks of gestation, the immune system is initiated from the yolk sac and aortic-gonad-mesonephros containing erythroid cells and macrophages that serve as tissue remodeling cells. Stem cells from the embryonic organs migrate to the liver, thymus, bone marrow, and spleen that influence the development of lymphocytes for Th1 and Th2 responses. Circulating monocytes appear around 18–20 weeks gestation and by 30 weeks they make up 3–7% of formed blood cells circulating the body (24).
Neonates, infants, toddlers, and older children all undergo immunological maturation, and their response to pathogens can be quite different from one another (84). Neonates are highly susceptible to invasive microbial infections likely due to immaturity of the immune system, and its immunosuppressive nature, likely to prevent deleterious reactions to colonizing microbes (5). Pre-existent low levels of lung function within the first year of life are associated with subsequent incidence of lower respiratory tract infection, suggesting decreased lung function is important in bronchiolitis caused by RSV (93). Increased risk of infections in toddlers could be a direct result of an immature immune system, but more research needs to be done as they do have differences from neonates with respect to innate and adaptive immune maturation likely due to differences in commensal exposures or prior exposures to RSV or other respiratory viruses. Although the rate of hospitalization decreases with infant age, the frequency of infection also increases with age (75). These data suggest that levels of immune development and lung physiological changes could affect the level of complications associated with RSV. Neonates have diminished cell surface-associated TLR responses although internal TLRs such as TLR7 or 8 appear to be more functionally similar to adults (76). Despite these more functional viral-sensing TLRs, neonatal plasmacytoid dendritic cell (pDC) exposed to viruses still release significantly less type 1 interferons than adults suggestive that other regulators such as epigenetics are limiting their capacity to respond (76). Similar findings for monocytes after dengue viral stimulation in the neonate have also been described (83).
Infant airway clearance of RSV relies on the innate immune response (macrophages and neutrophils) for early suppression of viral replication. Bias toward Th2 immunity in neonates appears to be an evolutionary adaptation against fetal and placental damage that continues after birth (45). During the transition from fetal life to birth, the neonate immune system must avoid excessive proinflammatory responses, so microbial-rich areas such as the gastrointestinal tract and skin can develop colonization of microbes for protection against foreign pathogens (21). However, the older infant and toddler switch toward a more inflammatory Th1 type response over Th2, but the extent of this switch is not well described.
IL-12 initiates antigen presentation in macrophages and contributes to a Th1 response, most notably inducing the production of IFNγ, whereas IL-18 (IFNγ inducing factor) contributes to protective immunity against pathogens and helps active T cells toward Th1 polarization as well. Neonates have reduced levels of IL-12, IL-18, and IFNγ in response to RSV infection (39). Infants with RSV continue to exhibit low IFNγ levels (4). In addition, infants (average 5 months old) with RSV-induced bronchitis can also exhibit low IL-18 secretion (42). However, RSV-infected IL-12p40−/− mice (although “adult mice”) demonstrated normal levels of Th1 responses and viral clearance but enhanced pathology linked to increased IL-5 and IL-13, which was further exacerbated by IL-18 neutralization (86).
Although low IL-12 and IFNγ production from innate and Th1 cells may fail to control viral replication to the same extent as older adults, a lack of these cytokines could also induce higher or simultaneous Th2 cytokine production that contributes to viral-induced morbidity. Furthermore, in vitro depletion studies of peripheral blood show that MHCII+ cells (possibly dendritic cells or monocytes) are the predominate IL-12 producing cell type in young children and that this cytokine increases with age while still remaining below adult levels (approximately half the levels in a 5 year old compared with adult and even lower for cord blood vs. adult) (82).
Pathogen-induced Th2 (IL-4 and IL-13) responses tend to arise with constrained TLRs signaling suggesting that low IL-12 from limited TLR ligation could possibly contribute to the younger child's propensity toward production of Th2 cytokines. However, not all Th1 stimuli switch to Th2 in the absence of IL-12 suggesting other mechanisms could also be at play (53). Furthermore, as stated prior, whereas TLRs such as TLR4 are limited in neonates, viral PAMPs such as RLR or TLR7/8 are not limited with respect to activating signals and downstream signaling. RSV, which expresses NS1 and NS2 proteins (80), can also contribute to viral-mediated suppression of type 1 interferons and IFNγ (72) and thus limit the capacity to release these cytokines from infant innate cells could be further compounded by viral suppression as well. Thus, it is evident that neonates may have an ineffective immune response to RSV that stems in part from the immunosuppressive nature of their immune system, but at what age these are overcome is not well known.
In contrast, the effects of the virus itself on immunity also cannot be ruled out. In infants sampled within 2–4 days of infection, nasal secretions typically contained lower levels of IFNγ and IL-4 [can be produced by innate lymphoid cells (56) or eosinophils] than induced by similar infants infected with influenza (48) further suggesting unique effects of viral-induced innate immune modification is possible.
Cross talk between macrophages and natural killer (NK) cells are a first line of defense against respiratory viral infections (50). Thus, alveolar macrophage depletion can significantly reduce and T cell recruitment to the lungs, therefore, reducing morbidity similar to direct NK cell depletion (30). Of interest, not only are NK cell numbers higher in the peripheral blood of neonates, but also their capacity for activation are inversely related to their numbers. Thus, not only is the type of mononuclear cells important to RSV innate immunity, but also is the context of how these cells are activated. Moreover, NK cells can make up about 10% of the lymphocyte population in adult lungs and, through secretion of IFNγ, can help clear RSV or suppress the virus in the early stages of infection (11).
Again, however, IFNγ is heavily epigenetically regulated in the neonate and reduced levels continue into toddlerhood suggestive that NK cells, through immunological immaturity, may not participate as fully in anti-RSV activities as found in adults. Few NK cells have been found in the lungs of infants during severe RSV infection (57) despite as stated prior that this population is found in higher numbers in the blood of infants than adults. This may stem to the fact that NK cell recruitment is dependent, in part, on macrophage signaling and that these cells may not be as active in the young child.
Decreased IFNγ Correlates with Decreased Activation of Macrophages in RSV Infection of the Young
IFNγ is a potent interferon that stimulates antimicrobial effects from macrophages in both an autocrine and paracrine manner, including the upregulation of antigen processing and presentation and effects are age related (15). Polymorphisms in IFNγ expression during neonatal RSV infection are linked to increased severity of illness, duration of stay in the intensive care unit, and frequency of otitis (22). Although low levels of IFNγ are thought to protect postnatal lung development, elevated levels are associated with milder RSV disease. Low production of IFNγ that may lead to less activation of monocytes/macrophages may be common in neonates and possibly toddlers, possibly contributing to RSV pathology.
In neonates, the production of cytokines by macrophages is limited when compared with adults, likely due to decreased IFNγ available in the lung antiviral cytokine milieu, but other factors such as IL-27 production could also contribute to the immunosuppressive nature of the neonate response (38). When adult mice are infected with RSV, a significant increase in IFNγ is present, but when neonates were infected with the same weight-based dose of RSV, there was an absence in IFNγ production, suggesting age-dependent activation of alveolar macrophages enhanced by IFNγ (15). In humans, there is increased capacity for production of IFNγ that continues through 8 years of age, which may represent the gradual development of the immune system from neonates, toddler, children, and adults (31). IL-12 expression in monocytes, a cytokine influencing Th1-directed immune response, was found to have correlation with age and intracellular IFNγ (31). In addition to limits on IFNγ production intrinsic in immune cells of neonates to toddlers, RSV viral infection of macrophages also can disrupt their ability to make other type 1 and 2 interferons (72) as stated previously.
Macrophage hyporesponsiveness to IFNγ (possibly from autocrine signaling or paracrine from NK cell production) may not be attributed to lower expression of IFNγ receptors or decreased affinity of the receptors, but rather a deficiency in the phosphorylation of IFNγ-receptor-associated kinase-signal transducer and activator or transcription STAT-1 (46). STAT-1 is important for a proinflammatory macrophage response and the poor signals may influence the decreased immune response (33). Whether this limitation in STAT-1 phosphorylation continues into toddlerhood (diminished IFNγ does) is not known. The neonatal and infant macrophage may deploy alternative cytokines to gain some control over viral replication in the milieu of differing immunosuppression, but this could also be at the determinate of the host (64).
Epigenetic Influence on Cytokines During Maturation of the Immune System
Epigenetics is important for controlling the mechanisms of the developing immune system from fetus to adult. Biological processes affected include inflammatory responses, humoral immunity, somatic recombination of immune receptors, patterning of effector immune responses, cell to cell signaling, and cellular homeostasis (47). Neonate cells exist in an epigenetic state that is markedly different from adults limiting immune responses and possibly, leaving them vulnerable to infection (3). Epigenetic modifications affecting macrophages include DNA methylation and histone modifications (3). DNA methylation changes with age, gender, and environmental factors (47).
Between birth and 12 months of age, immune pathways show significant changes in DNA methylation, specifically cell signaling and communication pathways (47). There are significant modulations in >484 demethylated and 330 methylated CpG sites within genes during 3 months to 5 years of age (1), showing the importance for analyzing epigenetic immunological maturation of toddler-aged children.
The JAK-STAT pathway (STAT4 and STAT6), which is important for IFNγ receptor phosphorylation, signal transduction, and inducing biological responses, is differentially methylated at birth, 2.5, and 5 years of age, showing the difference in immune maturation and CD4 T cell activation from birth to young children (13). In NK cells isolated from neonates, low pSTAT4 was observed when activated with IFNα or failed to translocate to the nucleus after IL-12 stimulation, which contrasted with infant NK cells that responded similarly to adults (63). Lower STAT1 phosphorylation has also been detected in stimulated neonatal macrophages upon addition of IFNγ over adults (46). Infant mice were found to have potential epigenetic regulation of genes downstream of STAT4 that failed to produce IFNγ in CD4 T cells during influenza infection (84), but whether this mechanism occurs in macrophages/monocytes are unknown.
Since STAT4 also plays a role in IFNγ production in dendritic cells and macrophages, further studies into epigenetic modifications of this pathway in young children and how they may be different in the older child might help better understand antiviral innate responses toward RSV. Furthermore, differences in DNA methylation of the IFNγ receptor pathway with age suggest a strong need for research on toddler immunological maturation and how it affects the fight against RSV.
IFNγ are not the only cytokine epigenetically regulated in the young child. IRF5, a transcriptional factor that controls TNFα expression, has also been shown to be downregulated in newborn type 1 macrophages upon TLR4 stimulation (70) and thus other cytokines may also be influenced by epigenetic mechanisms in young children too. Finally, a lack of IL-12 production in newborn monocytes was found to be possibly associated with other epigenetic regulation of transcription factors rather than direct regulation on the IL-12p35 or IL-12p70 promoters (9). Thus, understanding how immunological maturation may be controlled by epigenetic changes with age could be important to fully understanding mechanistic limits placed on the neonate/infant/toddler innate control over RSV.
Histone modifications impact maturation and development of the neonate immune system by activating or silencing gene transcription without affecting the DNA sequence (3). H3K4me3 is important for many functions, including developmental and immunological pathway that can lead to activation of gene transcription (3) especially with monocyte to macrophage differentiation pathways (92). In monocytes, a decrease in the poised enhancer mark H3K4met1 matches an increase in the active promoter mark H3K4me3 as development progresses (3). There is a linear increase in H3K4me3 as monocytes develop due to time and microbial exposure with influences on cytokines such as IL-12 or in microRNA regulators (3).
As maturation occurs, a large portion of H3K4me3 modifications are in metabolic pathways, which may be important to not only immune cell activation/proliferation/function but also signaling between cells as current research in immunometabolism suggests strong contributions from shifts in glycolysis/oxidative phosphorylation toward influencing responses. Neonatal immune cells have difficulty undergoing the metabolic shift from oxidative phosphorylation to glycolysis/lipid metabolism (12) for upregulation of proinflammatory genes, whereas adult monocytes present with increased H3K4me3 binding to those areas.
The lack of H3K4me3 in neonate monocytes is associated with a reduction in gene expression in vital immunological pathways, and may be a contributing factor to the heightened infection risk during the first few years of life (3). Macrophages from the upper and lower tract can diverge with respect to glycolysis versus continuation of oxidative phosphorylation (41). Adults display increases in H3K4me3 in monocytes with respect to glycolysis and lipid metabolism as compared with term infant monocytes (3). Thus, key differences in metabolic regulation could occur that only animal models or human cadaver studies (mucosal tissues) may answer. Furthermore, more detailed analysis of changes in gene methylation patterns in the developing infant to toddler to older child could further help understand regulation of their antiviral responses.
Conclusion
Although we know a lot about neonate immunity and its effect on RSV due to use of cord blood, not a lot of studies have focused on older infants or toddlers, who also do not have a fully mature immune system. This is striking because although neonates and young infants have a significant number of hospitalizations due to viral-induced bronchitis, older infants/toddlers (1–3 years old) have a high propensity for secondary bacterial pneumonia or otitis during RSV infection (85,88).
It is evident that the immune systems of these populations of young children are still developing; yet there are a limited amount of studies that look mechanistically at the causes outside cord blood studies. It is possible that the production of IFNγ is still remarkably reduced suggesting a cause for higher morbidity of RSV than older children or adults. Studies that analyze macrophage contributions and immunological maturation by grouping neonates and infants or combining toddlers into older age groups or simply grouping all children younger than five are not accounting for clear immunological differences due to maturation. Thus, we know remarkably little about how innate pulmonary immunity matures with age and the factors influencing it because of the difficulty accessing structural lung cells in children with bronchiolitis or in healthy controls.
There is a positive correlation between age and capacity for activation of macrophages, so although the immune system of a toddler should be more developed than infants, there are still limitations preventing them from fully limiting RSV on par with older peers. Poor recruitment, survival, or activities of alveolar macrophages could not only affect RSV suppression but could also have ramifications on lung repair after viral clearance. Clearly, age and immunological maturation are important concepts to study to better understand the pathogenesis of RSV in the young.
Footnotes
Author Disclosure Statement
No competing financial interests exist.