
Abstract
Enterovirus 71 (EV-A71) has attracted widespread attention in the past decade because of its higher neurotropicity in addition to causing hand-foot-and-mouth disease (HFMD). Mapping epitopes of viral proteins may promote our understanding of antiviral humoral immunity, and is useful for clinical application. In this study, the linear B cell epitopes within nonstructural proteins of EV-A71 were identified using bioinformatics methods combined with peptide enzyme-linked immunosorbent assay (ELISA). Twenty epitopes were predicted and located at 2ABC (7), 3ABC (5), and 3D (8) protein, respectively. Of 20 epitope-containing peptides, 14 were verified by ELISA (S/CO >2.1), 9 of which had higher reactivity (S/CO >5.0). Furthermore, synthetic peptide SP09 (EV-A71-3ABC26–41) can react with healthy children sera, and its immunoreactivity was closest to that of EV-A71-VP1 protein. The protein BLAST analysis revealed that SP09 may contain a common epitope due to the high homology of amino acid sequences with other members of human Enterovirus species A. These findings may be useful in the development serological tests for the diagnosis of HFMD caused by a broad range of human Enterovirus species A.
Introduction
Enterovirus 71 (EV-A71), belonging to Enterovirus genus of family Picornaviridae, is a nonenveloped single-stranded positive-sense RNA virus. Since first isolated in 1969 (12), EV-A71 has caused multiple outbreaks worldwide, particularly in the Asia–Pacific region (15). EV-A71 infection can cause hand-foot-and-mouth disease (HFMD), which is more common in infants and children younger than 5 years old (10). It is well known that EV-A71 is associated with a higher rate of central nervous system complications than other HFMD pathogens, such as coxsackievirus A6 (CV-A6), coxsackievirus A16 (CV-A16), and coxsackievirus A10 (CV-A10) (19). So far, however, there have been no effective antiviral drugs available.
The humoral immune response elicited by viral proteins is one of the important defense mechanisms of the host to eliminate viral infections. Sometimes, viral-specific antibodies may also contribute to the immunopathogenesis (17). So, immunodominant epitope identification will be helpful to develop new vaccines or diagnostic reagents, and even understand immunopathological mechanisms. The genome of EV-A71 contains only an open reading frame encoding four structural proteins (VP1, VP2, VP3, and VP4) and seven nonstructural proteins (2A, 2B, 2C, 3A, 3B, 3C, and 3D) (18). Until now, the most epitopes of EV-A71 have been identified within its structural proteins, especially VP1 (6 –8,22). However, the epitopes within its nonstructural proteins have not yet been well studied.
In this report, we screened and identified a series of linear B cell epitopes on nonstructural proteins of EV-A71 using bioinformatics prediction methods combined with peptide enzyme-linked immunosorbent assay (ELISA). Then, the immunoreactivity of these epitopes was evaluated using a panel of healthy child sera, and compared with that of VP1 protein of EV-A71.
Materials and Methods
Epitopes prediction and peptides synthesis
To predict the linear B cell epitopes on nonstructural proteins of EV-A71, three complete amino acid sequences (2ABC, 3ABC, and 3D) deduced from genome of EV-A71 strain HeN09 (GenBank accession no. JN256064) (13) were submitted to online servers, including ABCpred (
Expression and purification of recombinant EV-A71 nonstructural proteins
Three recombinant proteins (2ABC, 3ABC, and 3D) were expressed and purified according to the procedures described previously (3). In short, BL21(DE3) cells were transformed with the recombinant plasmids, and then grew in LB broth supplemented with 40 μg/mL kanamycin at 37°C. When the cultures reached OD600 of around 0.6, the cells were induced with isopropyl β-
Antisera specimens
As described previously, antisera against three recombinant EV-A71 nonstructural proteins were prepared by immunizing New Zealand white rabbits (21). In brief, each of the recombinant proteins was emulsified with an equal volume of Freund's complete adjuvant. New Zealand white rabbits (6–8-month old; 2–3.5 kg) were subcutaneously injected with 100 mL emulsion containing 50 mg proteins, and boosted twice with inoculation of the same antigen emulsified in Freund's incomplete adjuvant at 14-day intervals. Antisera were collected 7 days after the last immunization. All animals were housed in specific pathogen-free conditions and the studies were conducted according to the regulations of animal care and administration formulated by the State Science and Technology Commission of China.
In addition, a commercial rabbit antiserum against EV-A71 was purchased from Beijing Keygen Gene Technology Co., Ltd. Human serum specimens were collected from 80 healthy children under the age of 5 years in the Yuncheng district of Shaoxing city, Zhejiang, China. Human sera collection was approved by the ethics committee of center for disease control and prevention of Yuncheng district in Shaoxing city, and written informed consent was obtained from each subject.
Peptide ELISA
Polystyrene microtiter plates with high adsorption (Sangon, Shanghai, China) were used in the ELISA. For coating of the plates, 100 μL synthetic peptide (5 μg/mL) in carbonate buffer (pH 9.6) was added to each well and incubated at 4°C overnight. The plates were blocked for 1 h at 37°C with 5% skim milk powder in phosphate-buffered saline (PBS). Rabbit sera diluted 1:100 in PBS containing 5% skim milk powder and 0.05% Tween 20 (100 μL/well) were tested against each peptide. Preimmunized rabbit sera were used as the negative control. After incubation for 1 h at 37°C, the plates were washed six times in PBS containing 0.05% Tween 20. Subsequently, horseradish peroxidase (HRP)-conjugated goat antirabbit IgG (1:10,000 dilutions; KPL) was added and incubated for 1 h at 37°C. After six washes, the reaction was developed by 100 μL TMB (Sigma Aldrich) substrate, and then terminated by 50 μL 2 M H2SO4. The optical densities at 450 nm were determined in the microplate reader (Bio-Rad).
All samples were analyzed in triplicate and the results were expressed in S/CO value. Cutoff value was calculated as mean OD value of the negative control. The reaction with an S/CO ratio of <2.1 was defined as negative, ≥2.1 for positive, and ≥5.0 for strong positive (8). To identify the reactive peptides, human sera from healthy children were also tested in similar ELISA protocol except that the secondary antibody was replaced as HRP conjugated goat antihuman IgG (1:10,000 dilutions; KPL).
Results
Prediction, selection, and design of B cell linear epitopes
To improve the accuracy of prediction, the nonstructural protein sequences, including 2ABC, 3ABC, and 3D, were submitted to three online servers for B cell linear epitope prediction. The results obtained by the methods of AAP, BCPred, FBCPred, ABCpred, and BepiPred, respectively, are given in Table 1. Based on these results, overlapping peptide fragments from three or more methods were selected as the potential B cell epitopes. A total of 20 linear B cell epitopes were predicted: 7 from 2ABC, 5 from 2ABC, and 8 from 3D protein. To evaluate the immunoreactivity of these predicted epitopes, 20 peptides of 16 amino acids in length were designed and synthesized, and its amino acid sequences and locations are given in Table 2.
B Cell Linear Epitopes Within Nonstructural Proteins of Enterovirus 71 Predicted by Five Methods
Sequences and Localizations of Synthetic Peptides from Enterovirus 71 Nonstructural Proteins
Amino acid sequence according to EV-A71 strain M188-1181F/HeN/CHN/2009 (Genebank accession no., JN256064).
EV-A71, Enterovirus 71.
Epitopes verified by indirect ELISA
Antisera were prepared from rabbits immunized by three purified recombinant proteins, including 2ABC, 3ABC, and 3D, respectively. The immunoreactivity of 20 synthetic peptides was tested by indirect ELISA with corresponding antisera (Fig. 1). Results showed that 14 of 20 peptides were observed as positive reactivity (S/CO ≥2.1), 9 of which showed strong reactivity (S/CO ≥5.0), which are SP03, SP08, SP09, SP10, SP14, SP15, SP16, SP17, and SP20. In addition, none of these synthetic peptides can react with anti-EV-A71 rabbit serum (data not shown).
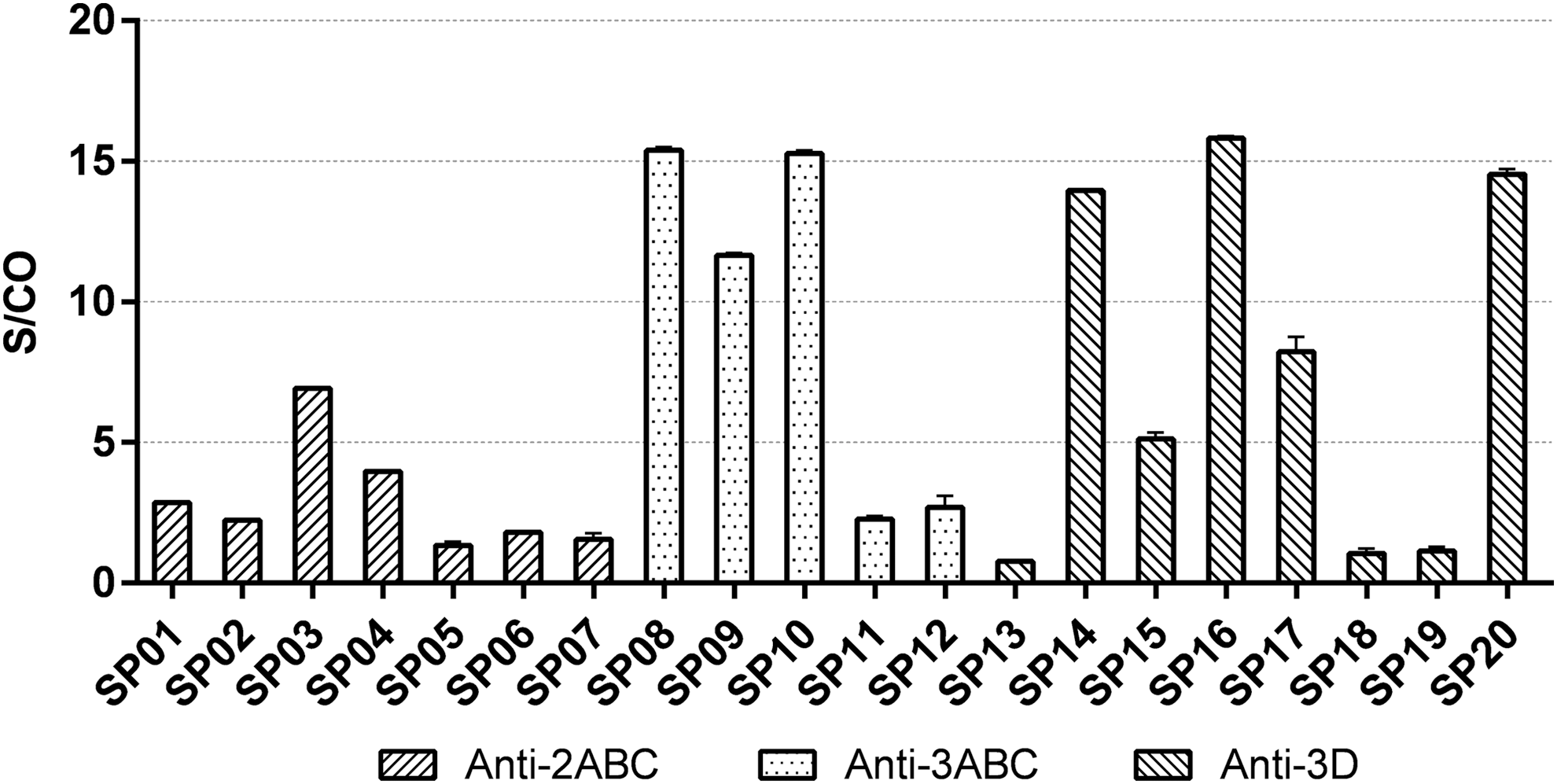
ELISA scanning of synthetic peptides. The entire set of purified peptides was subjected to epitope scanning by ELISA with corresponding rabbit antisera against EV-A71-2ABC, 3ABC, and 3D, respectively. Samples with an S/CO value ≥2.1 were considered positive, and those with S/CO value ≥5.0 were considered strong positive. ELISA, enzyme-linked immunosorbent assay; EV-A71, Enterovirus 71.
Comparison of immunoreactivity between candidate peptides and EV-A71 VP1
To evaluate immunoreactivity of nine candidate peptides already mentioned with human sera, anti-EV-A71 IgG antibodies were detected using indirect ELISA based on both synthetic peptides and EV-A71 VP1 (Fig. 2). Human serum specimens were collected from 80 healthy children under the age of 5 years. The results show that all of nine peptides can react with human sera to a certain extent, and among which SP09 had the closest immunoreactivity compared with EV-A71 VP1.
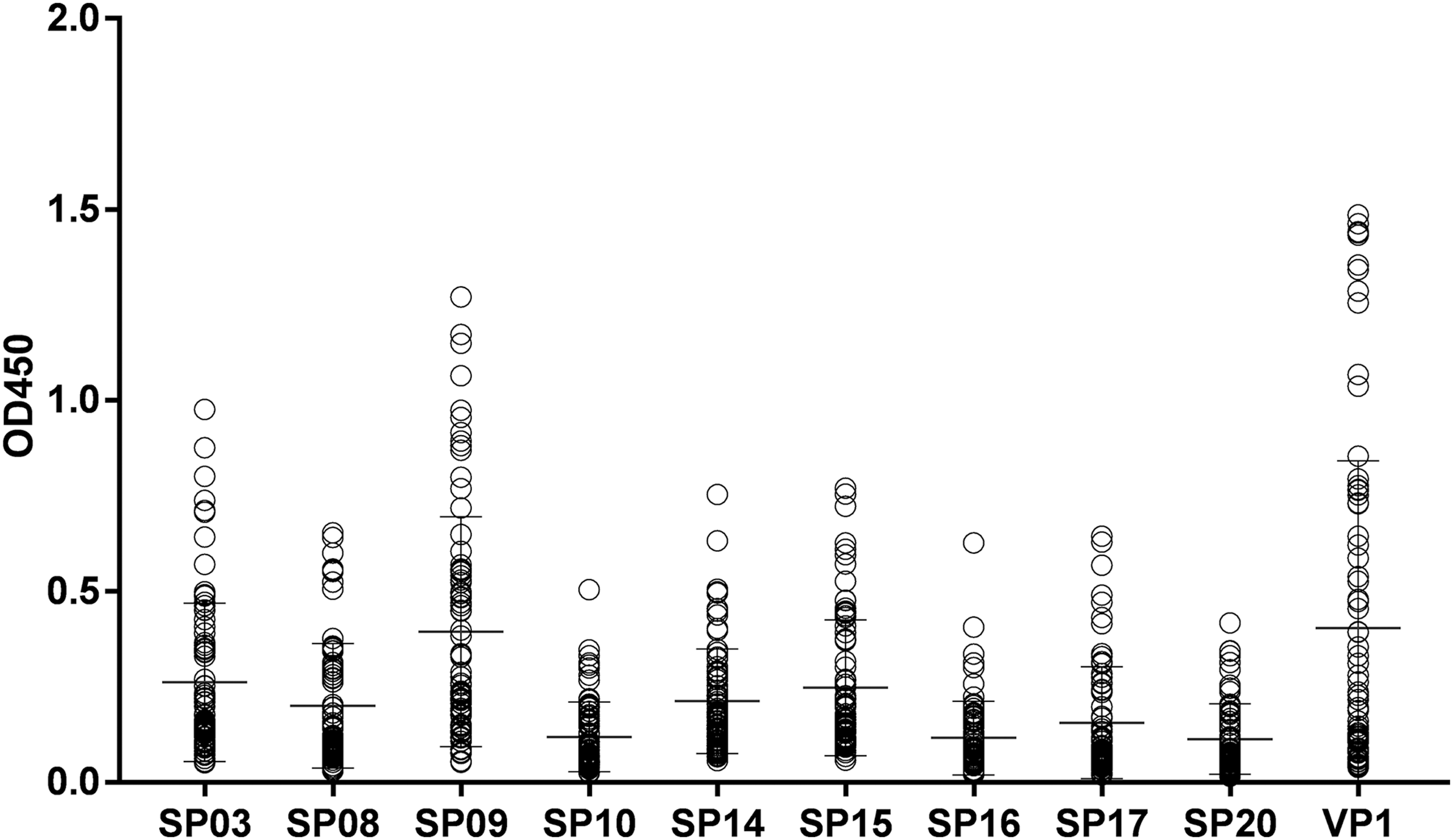
Serum reactivity profiles of candidate peptides in healthy children. Heterotypic IgG antibody responses against 9 candidate peptides and EV-A71 VP1 was shown in 80 healthy children's sera as measured by ELISA at an OD of 450 nm.
Discussion
Previous studies have shown that the nonstructural proteins of EV-A71 can elicit IgG antibody response against EV-A71, which is associated with viral infection or replication and not with vaccine immunization (3). However, the location of the immunodominant B cell epitopes within the nonstructural proteins has not yet been determined.
In this study, first, we used the bioinformatics methods to predict the potential linear B cell epitopes within the nonstructural proteins of EV-A71 (Table 1). Of the many online prediction platforms, five methods have been used to predict B cell epitopes, including BepiPred (9), ABCpred (11), AAP (2), BCPred (4), and FBCPred (5). These methods were developed based on the machine-learning technique, and have different sensitivity, specificity, and positive prediction values due to different algorithms, including hidden Markov model, artificial neural network, and support vector machine. In general, consensus predictions of multiple methods are more reliable than individual predictions. So, to ensure the accuracy of predictions, five methods were adopted in this study and overlapping peptide fragments from three or more methods were selected as the potential B cell epitopes. The result showed that a total of 20 epitopes were predicted: 7 from 2ABC, 5 from 2ABC, and 8 from 3D protein (Table 2).
Then, 20 epitope-containing peptides were synthesized and used to analyze its immunoreactivity using peptide ELISA. The result shows that of 20 epitope-containing synthetic peptides, 14 were positive by ELISA (S/CO >2.1) and 9 had higher reactivity (S/CO >5.0) (Fig. 1). So, the prediction accuracy based on combination strategy seems quite satisfied. In addition, consistent with previous studies, none of these epitopes can react with rabbit antisera against EV-A71 viral particles (data not shown).
Finally, considering the possibility that these epitopes may be used in seroepidemiological surveys of enterovirus infection in child population, we used healthy child sera instead of the HFMD patients' sera to evaluate the immunoreactivity analysis of the candidate peptides. Eighty healthy child sera (<5 years old) were included in ELISA. The result showed that synthetic peptide SP09 (3ABC26–41) had highest positive rate, and its immunoreactivity is closest to that of VP1 protein of EV-A71.
In addition, protein BLAST results (data not shown) showed that the amino acid sequences of SP09 were highly identical (almost 100%) to other members of human Enterovirus species A, such as CV-A16, CV-A10, and CV-A6. These viruses are also common pathogens of HFMD, and have been prevalent in mainland China over the past decade (16). These results indicate that the epitope located at position 26–41 amino acid of 3ABC is immunodominant in human population, and most likely the common epitope of human Enterovirus species A.
Some studies have sought to find common epitopes of enteroviruses for early diagnosis, but usually focused on the structural proteins of the virus (1,14). A recent study has shown that nine peptide segments can be recognized by IgG antibodies from HFMD patients, seven of which are located at the nonstructural proteins of EV-A71 (20). Of the seven peptide segments, the amino acid sequence of P102 and P147 almost overlaps with that of SP03 and SP09, which is almost consistent with our results. In particular, P147 (similar to SP09) located at the N-terminal portion of 3A protein can react with both IgG and IgM antibodies against EV-A71, which indicates that this peptide may be used as an antigen to detect HFMD.
In conclusion, we have found nine linear B cell epitopes within nonstructural proteins of EV-A71, of which an epitope located at position 26–41 amino acid of N-terminal part of 3ABC is immunodominant in human population. These findings may be useful in the development serological tests for the diagnosis of HFMD caused by a broad range of human Enterovirus species A.
Footnotes
Acknowledgments
The authors are grateful to Dr. Longfei Mao (center for disease control and prevention of Yuncheng district, Shaoxing city) for his help in collecting healthy children sera samples. This work was supported by grants from the Natural Science Foundation of Zhejiang Province (grant no. LY16H190005).
Author Disclosure Statement
No competing financial interests exist.