
Abstract
Dengue virus infection (DEN) is one of the most prevalent arbovirus diseases in the tropical and subtropical areas. Some human leukocyte antigen (HLA) alleles have been reported to be a protective or risk factor to DEN. Due to the limited sample sizes and regional limitations, the results of individual studies were various. This meta-analysis aimed at investigating the relationship between HLA alleles and dengue disease. Relevant studies of the relationship between HLA and dengue disease were searched through PubMed, Embase, Web of science, and Cochrane databases. Subgroups according to ethnicity or sub-alleles and sensitivity analysis were used to explore the potential source of heterogeneity, which was performed to confirm the findings. The relationships between HLA and dengue disease were defined by odds ratios (ORs) with a 95% confidence interval (CI). Fourteen studies were finally confirmed. Results indicated that A*0203 (OR = 2.19, 95% CI = 1.30–3.69) and A*24 in the Asian group (OR = 1.44, 95% CI = 1.21–1.71) were positively associated with an increased risk of DEN when compared with normal controls. A*33 (OR = 0.49, 95% CI = 0.34–0.69) in Southeast Asia was negatively associated with DEN when compared with normal controls, suggesting a protective role against DEN. In addition, DRB1*11 (OR = 4.10, 95% CI = 1.23–13.69) was positively associated with severe dengue (SD) when compared with dengue fever, whereas DRB1*03 (OR = 0.48, 95% CI = 0.28–0.82) and DRB1*09 (OR = 0.73, 95% CI = 0.55–0.96) were negatively associated with SD when compared with normal controls. The meta-analysis confirmed that HLA-A*0203, A*24, A*33, DRB1*03, DRB1*09, and DRB1*11 have significantly affected dengue disease, and the associations are related to race and regions.
Introduction
Dengue virus infection (DEN) has become one of the most common public health problems in the world. More than half of the global population is at risk for dengue infection, with 100 million symptomatic cases being reported every year (25,39). The virus belongs to the single-stranded RNA virus and the Flaviviridae family, which has four serotypes (DEN-1, -2, -3, and -4) (9,53); DEN is a self-limited illness, but it can manifest in a range of symptoms, from mild flu-like illness (25) and rash (dengue fever, DF) to severe dengue (SD) fever. SD, including dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS), can endanger the lives of patients, and it is characterized by plasma extravasation, thrombocytopenia, coagulation abnormalities and hypovolemic shock (54).
The pathogenesis of SD remains unclear, and there are several hypotheses, of which the secondary infection theory is the most impressive one, namely the antibody dependent enhancement (ADE) hypothesis (34). The ADE hypothesis implies that secondary infections may develop into SD, and SD disease is seen in only 1% of DEN cases; however, mortality in severe cases can have a rate of up to 20% (45). Some investigators have suggested that human genetic factors, such as human leukocyte antigen (HLA) alleles, may play an outstanding role in susceptibility or protection in dengue disease (37).
HLA is controlled by the major histocompatibility complex and is located on the short arm of chromosome 6. After decades of research, HLA has been considered the most important genome associated with infection and immunity (8). The immune mechanism of the HLA system and its polymorphism is highly correlated with the occurrence and development of many infectious diseases (28,47). HLA-A delivers virus-specific dengue to specific CD4+ and CD8+T cells (7,13,20), which is one of the most important targets in the development of dengue vaccines. Therefore, HLA might be related to the development mechanism of dengue disease and may be used as an early diagnosis or prognostic marker.
We found that the earliest study (12) that reported an association between HLA and dengue disease was published several decades ago, followed by some articles about the association between HLA polymorphism and dengue disease. However, the results of individual studies are still controversial, and the well-established information was limited. To further investigate the relationship between HLA and dengue disease, we intended to summarize the available independent research results of HLA loci and dengue disease by performing a meta-analysis approach.
Materials and Methods
Search strategy
We retrieved four databases: PubMed, Embase, Cochrane, and Web of science up to May 30, 2018. The terms used were “dengue,” “severe dengue,” and “HLA Antigens” as the medical subject heading (Mesh) and text. In addition, we manually retrieved relevant papers to expand the search scale. The same terms were used in four databases. No language or region or publication year restriction was adopted.
Selection criteria
Inclusion criteria: (i) molecular genotyping; (ii) case–control trials of random selection to study the relationship of HLA and dengue; (iii) no restriction of the region, age, or publication year; (iv) studies providing original data; (v) applicable to Hardy–Weinberg equilibrium; (vi) normal controls without the dengue infection symptoms or laboratory data of dengue infection. Exclusion criteria: (i) irrelevant studies, reviews, and case reports; (ii) no control group; (iii) in vitro or animal model; (iv) infections of recombinant proteins from dengue virus; (v) studies providing insufficient data; (vi) repeated publication; (vii) serological typing.
Data extraction
After excluding the duplicates, papers without full text, and irrelevant studies, the rest of the studies were read by two researchers independently. They screened the title and abstract, had a preliminary understanding of the article, and reached a consensus, extracting the following data from the article: author, time of publication, region, diagnosis citations of disease, the sample size of case group and control group, genetic testing methods, and type loci. The third researcher was involved when the two had different opinions.
Assessment of study quality
Two reviewers independently assessed the quality of each of the included studies. If there was a dispute over a study, the third reviewer was asked to join the discussion and arrived at a consistent result. The quality evaluation criteria were based on a case–control study of the Newcastale-Ottawa scale (NOS). The evaluation included three aspects of study selection, compatibility, and exposure. It was considered good quality if the study reached six points over eight points (21).
Statistical analysis
The meta-analysis was performed by using Revman 5.3. Odds ratios (ORs) and 95% confidence interval (CI) were used to assess the relationship between type loci and DEN. I2 statistic was used to evaluate the heterogeneity of the studies (31). Both the random-effect and fixed-effect models were used. A fixed-effect model was used without significant heterogeneity, whereas a random-effect model was used with significant heterogeneity. When heterogeneity was unacceptable, subgroup analysis of region or subsite and sensitivity analysis, as well as random-effect model were conducted. Subgroup analysis was used only when the number of included studies was more than four and that could be divided into two groups or more. We did not perform publication bias because of low quantitative amount when the number of the included studies was small (29).
Results
Study characteristics
There were 721 studies, including PubMed (n = 88), Embase (n = 218), Cochrane (n = 5), and Web of science (n = 410). Two hundred forty-seven duplicates were excluded, 436 irrelevant studies were excluded from screening titles and abstracts, 5 articles could not be obtained full-text, and 19 articles were excluded after reading the full text based on violating the inclusion criteria. Therefore, 14 studies were included and the selection process is shown in Figure 1. All 14 studies were case–control studies (2 –5,10,11,19,32,33,35,36,38,46,48), published from 2002 to 2015. The meta-analysis included 2,386 patients and 2,374 healthy controls. Alagarasu et al. published two articles in the same year (2013) (3,4), both of which were included in the study (numbers have been used to identify the author's different studies). The characteristics of the included studies are shown in Table 1.

Flow diagram of study inclusion.
Characteristics of Included Studies in the Meta-Analysis
NOS, Newcastale-Ottawa scale; PCR, polymerase chain reaction; RFLP, restriction fragment length polymorphism; SSO, sequence-specific oligonucleotide; SSP, sequence-specific primers.
Assessment of study quality
According to the quality evaluation of the studies, 5 scored 8 points, 5 scored 7 points, and 4 scored 6 points, so there was an average of 7.07 points (higher than 1.07 points in good quality). Therefore, they all were included. The detailed NOS scores are shown in Supplementary Table S1. The research areas were mostly in dengue infection prevalent areas. The comparative groups of the study were also different. To identify whether HLA was statistically significant between the dengue groups, we compared DEN (including DF and SD), DF, and SD with normal controls, respectively, as well as DF and SD.
Association of HLA-A with dengue
In the subgroup analysis based on the HLA-A*02, a positive association with the DEN group was detected in the A*0203, compared with the normal controls (two studies, OR = 2.19, 95% CI = 1.30–3.69, p = 0.003); whereas the association went by contraries in the other A*02 sites (seven studies, OR = 0.92, 95% CI = 0.77–1.09, p = 0.33), shown in Table 2 and Figure 2.
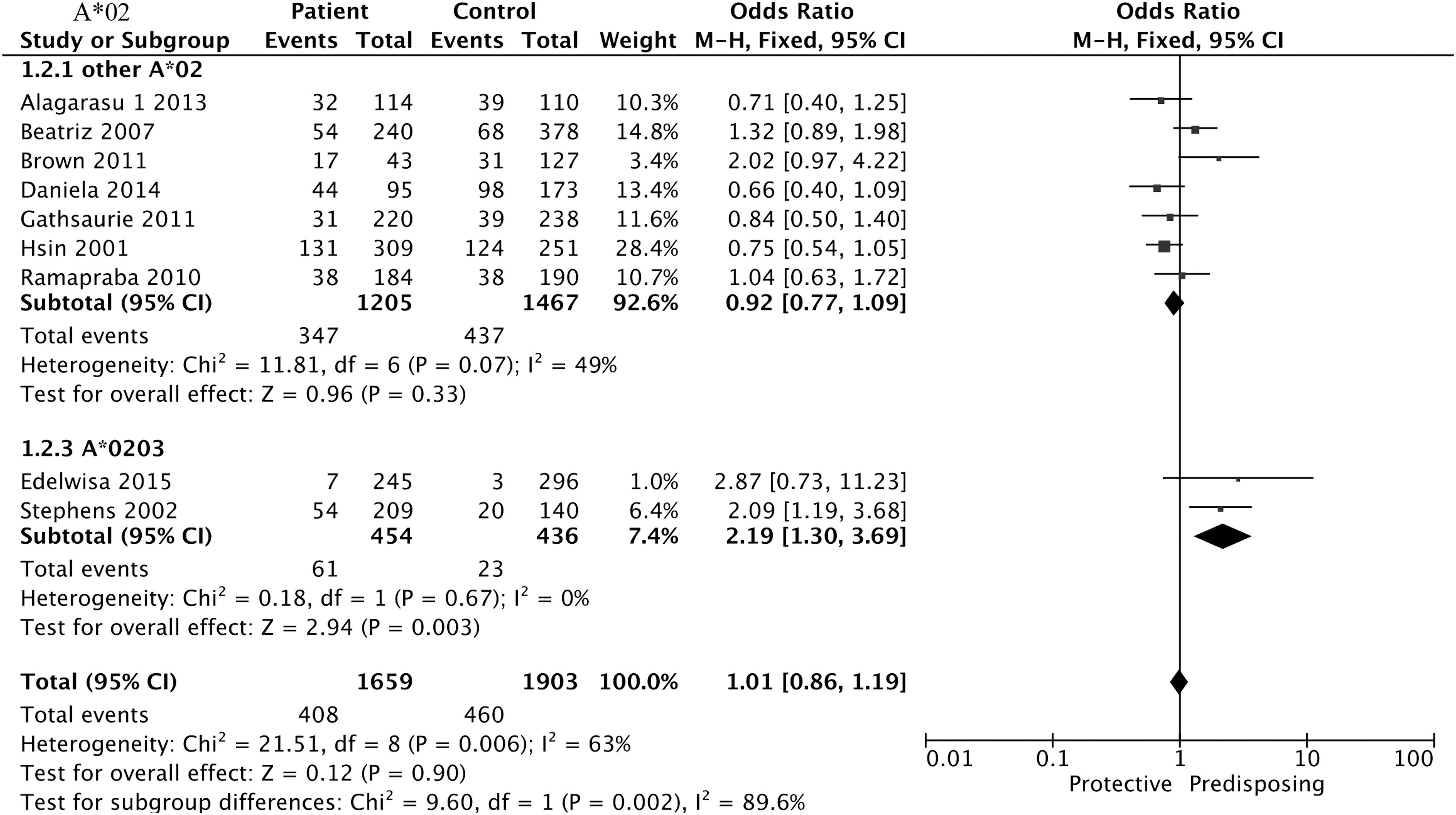
Forest plot for association between HLA-A*02 and DEN compared to normal controls. DEN, dengue virus infection; HLA, human leukocyte antigen.
Meta-Analysis and Subgroup Analysis of the Association Between Human Leukocyte Antigen and Dengue Virus Infection (Including Dengue Fever and Severe Dengue) Compared to Normal Controls
CI, confidence interval; DEN, dengue virus infection; F, fixed-effects model; OR, odds ratio; PRE, predisposing; PRO, protective; R, random-effects.
Eight studies of HLA-A*24 were analyzed with insufficient heterogeneity (I2 = 50%, p = 0.05) by using a fixed-effect model. Results showed that HLA-A*24 was positively associated with DEN compared with normal controls (OR = 1.41, 95% CI = 1.20–1.64, p < 0.001), depicted in Table 2 and Figure 3. The result of the Asian group (OR = 1.44, 95% CI = 1.21–1.71, p < 0.001) was similar to before grouping (OR = 1.41, 95% CI = 1.20–1.64, p < 0.001), supporting the strong positive association with DEN that was seen with A*24. Due to the limited numbers of studies, we did not compare A*24 between DF and normal controls as well as between SD and DF.
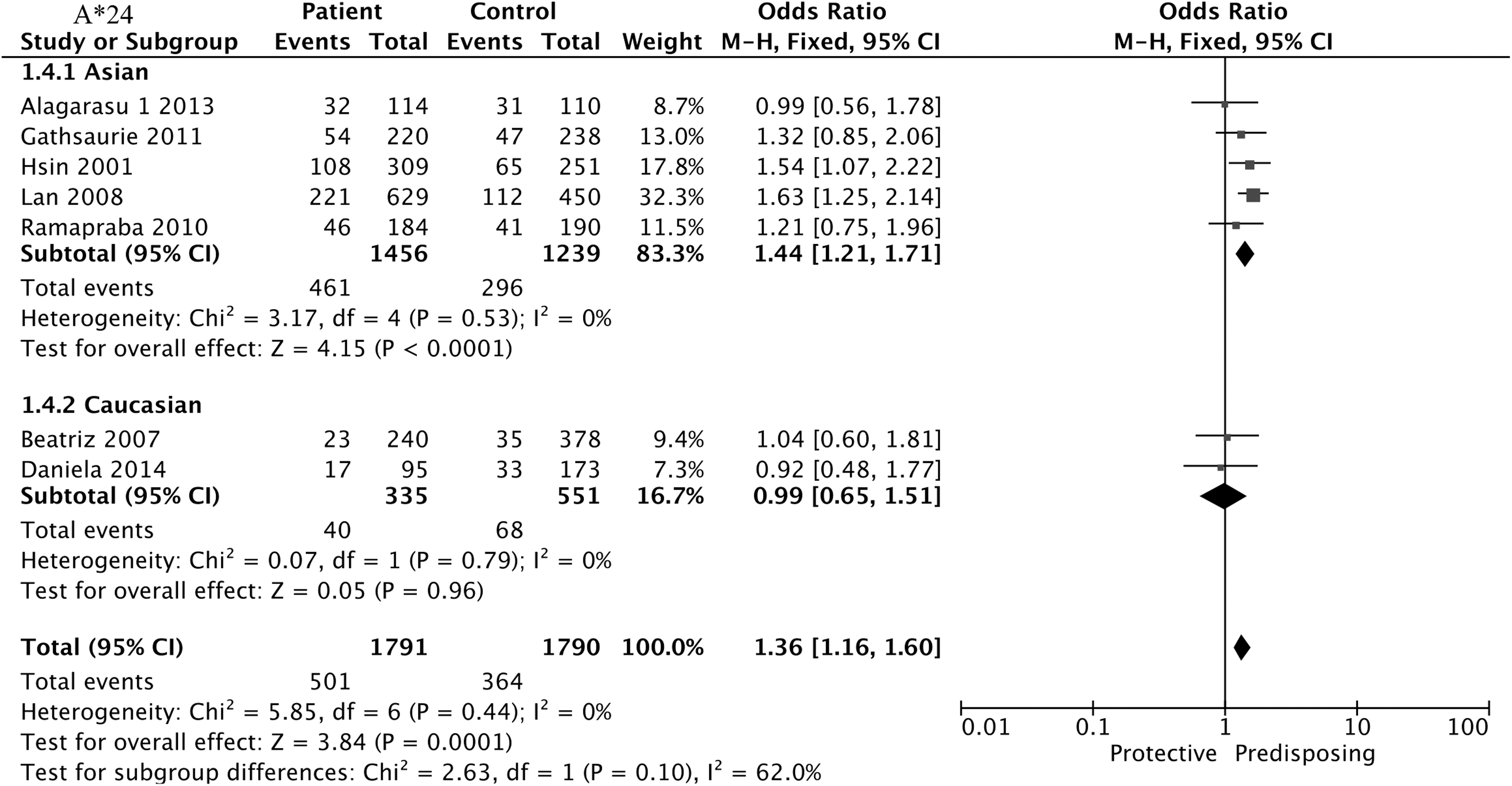
Forest plot for association between HLA-A*24 and DEN compared to normal controls.
The results of association between A*33 and DEN are summarized in Table 2 and Figure 4. Negative association was found between DEN and the normal controls in the subgroup of Southeast Asian A*33 (three studies, OR = 0.49, 95% CI = 0.34–0.69, p < 0.001), and the remaining subgroups could not mirror the result.
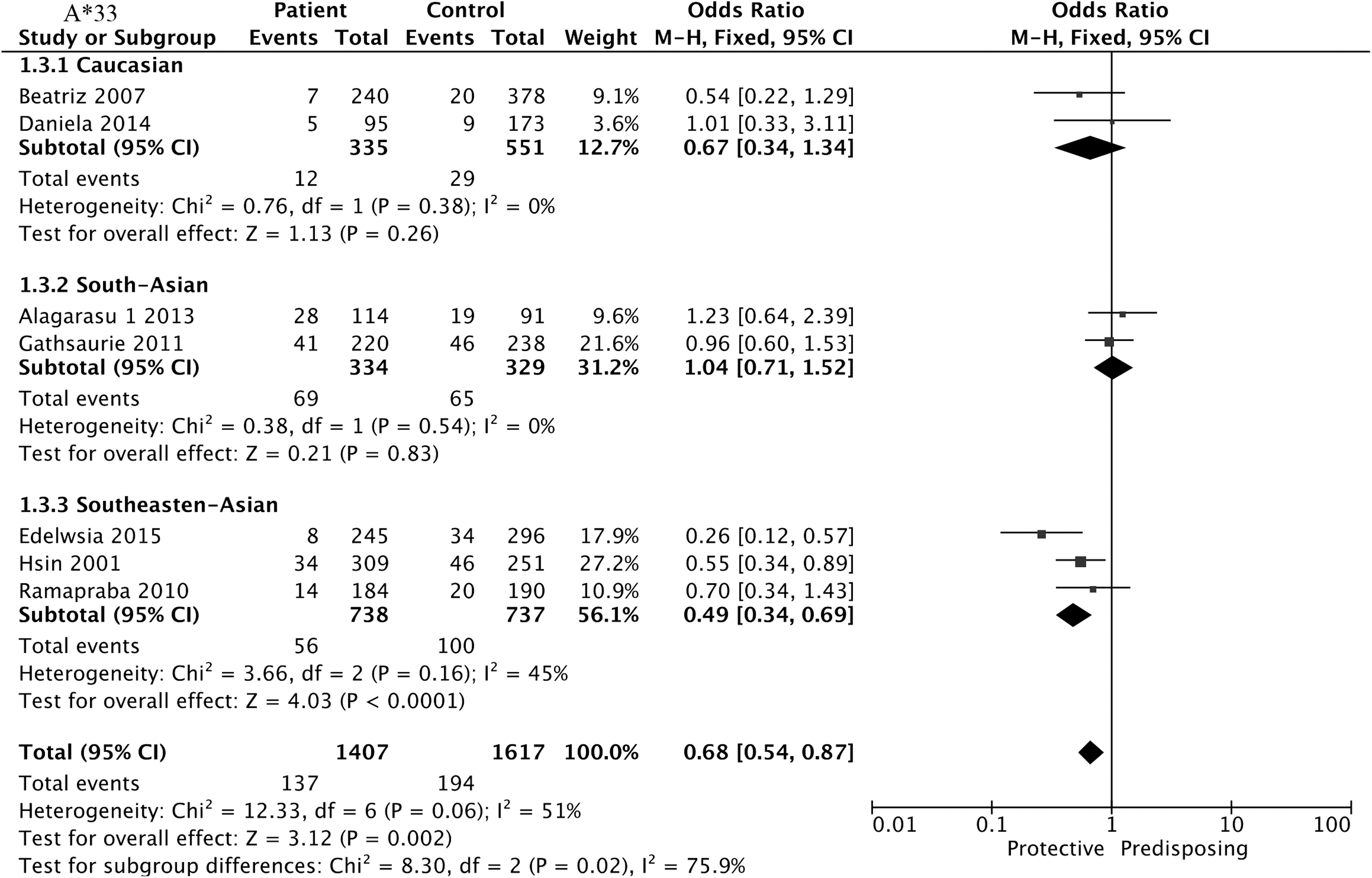
Forest plot for association between HLA-A*33 and DEN compared to normal controls.
Association of HLA-B with dengue
We included two HLA-B alleles in our meta-analysis, namely HLA-B*15 and HLA-B*18. As listed in Tables 2 –5, the two alleles showed no statistical significance in the four groups, presented in Supplementary Figures S1–S4.
Meta-Analysis of the Association Between Human Leukocyte Antigen and Severe Dengue Compared to Normal Controls
Meta-Analysis of the Association Between Human Leukocyte Antigen and Dengue Fever Compared to Normal Controls
Meta-Analysis of the Association Between Human Leukocyte Antigen and Severe Dengue Compared to Dengue Fever
Association of HLA-DRB1 with dengue
HLA-DRB1*03, HLA-DRB1*07, HLA-DRB1*09, and HLA-DRB1*11 alleles were included in the meta-analysis. The results found that DRB1*11 (OR = 4.10, 95% CI = 1.23–13.69, p = 0.02) was positively associated with an increased risk of SD when compared with DF (Fig. 5), whereas DRB1*03 (OR = 0.48, 95% CI = 0.28–0.82, p = 0.008) and DRB1*09 (OR = 0.73, 95% CI = 0.55–0.96, p = 0.03) were negatively associated with SD when compared with normal controls (Figs. 6 and 7). However, the three alleles showed no statistical significance in the remaining three comparisons, as did DRB1*07 in the four groups, shown in Tables 2 –5.

Forest plot for association between HLA-DRB1*11 and SD compared to DF. SD, severe dengue.
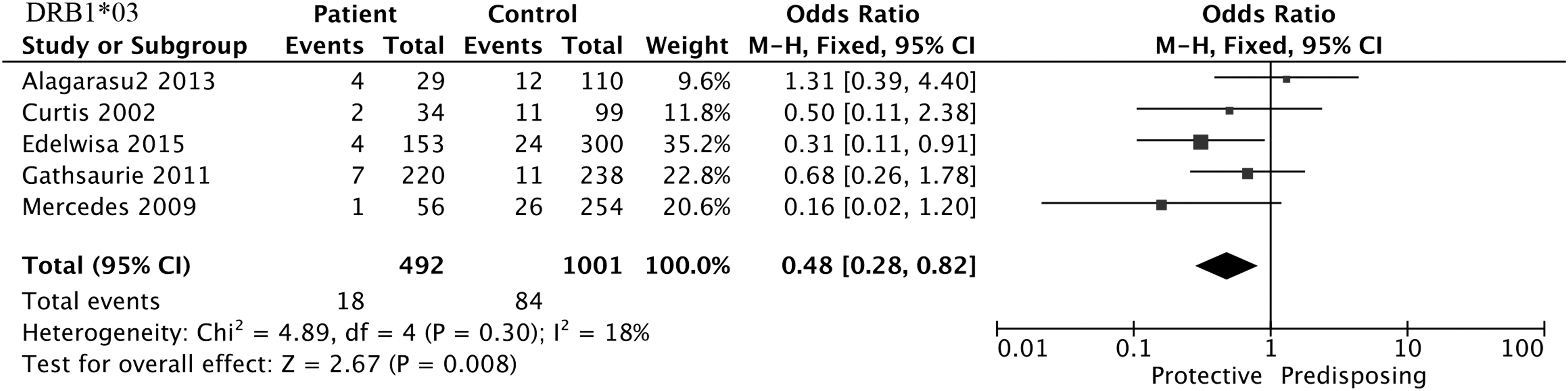
Forest plot for association between HLA-DRB1*03 and SD compared to normal controls.
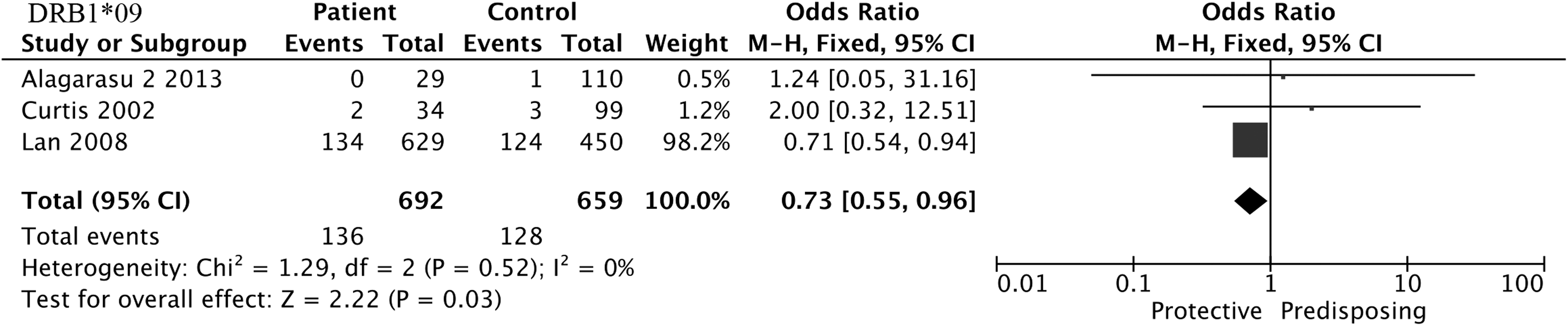
Forest plot for association between HLA-DRB1*09 and SD compared to normal controls.
Sensitivity analysis
Sensitivity analysis was performed to examine the effect of individual study on the whole quantitative synthesis of OR. We found that the new pooled OR indicated no significant change, ranging from 0.98 (95% CI = 0.75–1.28) to 1.08 (95% CI = 0.80–1.44) in the analysis of HLA-A*02 after omitting any of the individual studies. It was interesting that the heterogeneity in the two A*02 subgroups was insufficient (other A*02: I2 = 49%; A*0203: I2 = 0%) when we conducted a subgroup analysis of sub-allele, suggesting that the subsite may be the source of heterogeneity.
It was worth mentioning that I2 changed substantially (I2 = 0%) when the study of Brown et al. was eliminated, but there was no such change in the new pooled OR (OR = 1.36, 95% CI = 1.16–1.60, p < 0.001). We employed two subgroups to detect the substantial change in the new pooled OR of A*24 after removing Brown et al. (African descent). The Asian group matched the result to the previous group, whereas the Caucasian group did not show this association.
No apparent evidence revealed that the new pooled OR exhibited a distinct change after omitting the individual study. When focusing on whether the region was the possible source of heterogeneity of A*33, we grouped it by Caucasians, South Asians, and Southeast Asians after excluding Brown et al. (African descent). The new pooled OR of Southeast Asians (OR = 0.49, 95% CI = 0.34–0.69, p < 0.001) manifested significant change after grouping. It was found that ethnicity, region, and publication year could not explain the between-studies heterogeneity of the remaining characteristics. The different sex, age, study design, mean disease duration, and varied characteristics of participants could all be partly responsible for this heterogeneity (13,51).
Discussion
In recent years, several case–control studies have explored the association between HLA alleles and DEN but provided inconsistent results. This study aimed at summarizing whether the available HLA alleles have a protective or risk effect on different forms of dengue infection. Based on data from the 14 case–control studies, the main findings of the meta-analysis were that HLA-A*0203, HLA-A*24, HLA-A*33, HLA-DRB1*03, HLA-DRB1*09, and HLA-DRB1*11 were significantly associated with dengue disease. To our knowledge, it was the first meta-analysis to investigate the association between HLA and dengue disease.
We only included controversial alleles rather than all alleles with data, and we also performed subgroup analysis to explore the degree to which potential confounders might have influenced the findings. The A*0203 was positively associated with an increased risk of DEN when compared with controls. In the A*0203 subgroup, Stephens (48) considered that A*0203 was more frequent in secondary infections than in healthy controls, showed a strong promotion in the Thai population; whereas Mercado et al. (38) compared the DEN with the healthy control group, which indicated that the frequency of the DEN was higher than that of the healthy control group, and the promotion effect in the Filipinos was not strong in Thailand. More research was needed to support their argument.
HLA-A*24 was positively associated with DEN in Asians; this result was consistent with previous studies by Lan et al., Loke et al., and Malavige et al. (33,35,36). They had shown that HLA-A*24 was involved in the development of DHF or DSS, whereas Brown et al. (10) studied the relationship between HLA-A*24 and DF. Malavige et al. (36) discussed the primary and secondary infections of dengue virus, contributing to the mechanism of SD. All four studies believed that HLA-A*24 was positively associated with the development of DF or DHF.
The association between A*24 and dengue disease was most reported in Asian populations. When the A*24 included articles were removed one by one for sensitivity analysis, it was found that one article was different from other articles. The strength of HLA disease may differ in different races, regions, or genders (16,30). Therefore, we conducted regional grouping excluding Brown et al.'s study (African descent). Although the positive result could not be replicated in Caucasians, A*24 was statistically greatly associated with the Asian group. The different result could be explained by the difference in the presence or absence of the site in each population (22,27,43,49).
On the other hand, HLA-A*33 was negatively associated with DEN in Southeast Asians and had no sufficient effect in Caucasians and South Asians. There were two articles with very different conclusions: Mercado et al. (38) (Philippine) and Loke et al. (35) (Vietnam) believed that A*33 was negatively associated with SD, whereas Brown et al. (10) (Jamaica) and Alagarasu et al. (4) (India) believed that A*33 was positively associated with DEN.
These two conclusions could not be compared because their comparison groups were different. The participants in both Mercado et al.'s study and Loke et al.'s study were children, whereas the participants in both Brown et al.'s study and Alagarasu et al.'s study were not limited by age. However, Alagarasu et al. found that A*33 was observed with higher frequency in DEN compared with normal controls but without statistical significance. When DEN was classified into DF and SD cases, A*33 revealed a significantly positive increased risk associated with DF compared with normal controls. We conducted a subgroup analysis of A*33, which was divided into the Caucasian group, the Southeast Asia group, and the South Asia group. The A*33 subgroup analysis excluded Brown et al. (African descent). Heterogeneity disappears after grouping, suggesting that the region or race may be the source of heterogeneity.
In the meta-analysis, we confirmed the association of the three HLA-DRB1 alleles with dengue disease. DRB1*11 was positively associated with SD when compared with DF, whereas both DRB1*03 and DRB1*09 were negatively associated with SD when compared with normal controls. The class II locus, DRB1, is involved in the pathogenesis of several diseases (1,6), which has been predicted to be able to be joined by proteins of dengue virus (46). Besides, HLA class II alleles were associated with the phenotype of responding to cytotoxic CD4+ T cells, and they reported an increased resistance or susceptibility to SD (50). The functional correlation between HLA-DRB1 and the development of dengue disease has not been confirmed.
HLA-B*15 was found to exhibit increased frequency in SD compared with normal controls (Sierra et al.) (46) (Cuba), whereas Fernández-Mestre et al. (19) (Venezuela) found that B*15 was significantly reduced in frequency in DEN compared with healthy controls. The difference might be contributed to race and region or the different comparison. It was interesting to note that the result of combining all of the B*15 individual studies showed no statistical significance. This could be attributed to increased sample size. Alagarasu et al. (4) indicated that the frequency of HLA-B*18 was significantly higher in the dengue group compared with healthy controls (p = 0.047, pc = 0.846, OR = 3.53). Again, we unearthed no such association after combined overall studies.
Both A*24 and A*33 were associated with DEN; however, the association could not be proved until Brown et al.'s study was excluded. For the reason that Brown et al.'s study was the only one that studied African descent, the results of A*24 and A*33 showed difference compared with the researches in other ethnicities. It has been prospectively studied that SD is a rare consequence to Africans (15,26) and HLA frequencies play a role in it. The results still made sense, but more evidence was needed to support this claim.
The mechanism by which HLA is associated with dengue disease is still unclear. HLA-A has long been reported to have a promoting or protective effect on dengue patients (51). What is more, genome-wide association analysis also suggested that HLA-A alleles were closely associated with DF (24). This would indicate that HLA class-restricted cross-reactive T cell-driven immune responses.
Four HLA-A*02 and three HLA-DR1 restricted dengue viral epitopes have been recognized (7,23). Results highlighted the key role of the HLA, which demonstrated DEN responsiveness and defined phenotypic characteristics associated with the specificity of dominant T cell responses (14,52). In the absence of effective early diagnostic markers for dengue, HLA is, indeed, worth studying (44). HLA is an antigen-presenting molecule on the cell surface. Its polymorphism may alter immune regulation and is immunologically relevant to dengue disease, which is associated with weaker CD8+ responses (51). HLA also acts on cells infected with dengue virus through T cell-related functions, changing in epitopes and regulating T cellular interactions (17,40,42). These are the reasons that so many scholars study the relationship between HLA and dengue disease.
A comprehensive search and strict inclusion criteria were conducted. As a result, the articles included reached an average score that was higher than a good quality (7.07/6). This meta-analysis was based on generic data; SSO and SSP typing are the main approaches to the current criteria (18,41). The meta-analysis intended to divide dengue cases into three groups: DEN (included DF and SD), DF, and SD, compared with healthy controls, respectively. In addition, another group of SD was compared with DF. This grouping could make the results more credible and evaluate the association between HLA and different forms of dengue. Although the early prediction of SD is invaluable, distinguishing the people who have infected dengue virus carrying a high likelihood of developing SD is also worth exploring (24).
There are still limitations in this meta-analysis, which are reflected in the following aspects. First, due to the research limitations of the included articles, there are more potential molecular markers or other alleles of HLA that we did not include. Second, there were four different diagnostic criteria used in the studies included in the current meta-analysis, which may make the patients exhibit a little different heterogeneity. Third, most of the studies included did not address first or second infections. Although the pathological mechanism of DEN is not clear, many papers showed that secondary infections contribute to SD disease. However, only three articles included in this study have contributed to this discussion (33,36,48).
Conclusion
This meta-analysis was performed with low heterogeneity, indicating the robustness and reliability of the findings, demonstrating strong associations between HLA and dengue disease. The associations were related to race and region, which conferred susceptibility or protection against the DEN, ranging from DF to more SD hemorrhagic fever and DSS. In the case of dengue disease, considerate grouping allows us to understand more about the relationship between HLA and dengue disease. HLA-A*0203, A*24 and DRB1*11 were the risk factors of dengue disease, whereas HLA-A*33, DRB1*03, and DRB1*09 were the protective factors.
Footnotes
Acknowledgment
This work supported by Guangzhou Science Technology and Innovation Committee (No. 201607010163), Health and Family Planning Commission of Guangdong Province (No.A2016448),and Guangzhou Medical University (No.2014C24). The authors would like to thank PhD Shankun Zhao for his insightful feedback on the pre-submission of this article.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.