
Abstract
Matrix (M) protein of Newcastle disease virus (NDV) is an abundant protein that can induce a robust humoral immune response. However, its antigenic epitopes remain unknown. In this study, we used a pepscan approach to map linear B cell immunodominant epitopes (IDEs) of M protein with NDV-specific chicken antisera. The six epitopes with the highest reactivity by peptide scanning were obtained as IDE candidates. Among them, aa71–85 and aa349–363 were identified by immunological assays with NDV-specific or IDE-specific antisera. The minimal antigenic epitopes of the two IDEs were further characterized as 77MIDDKP82 and 354HTLAKYNPFK363. Moreover, an amino acid sequence alignment and immunoblot analysis revealed the conservation of the two IDEs in the M protein of strains of different genotypes. These two IDEs of M protein could be genetically eliminated as negative markers in recombinant NDV for serologically differential diagnosis in the development of marker vaccines.
Introduction
Newcastle disease is a highly contagious disease caused by the virulent strain of Newcastle disease virus (NDV) in avian species; it causes economic losses in the poultry industry (24). NDV, also named avian paramyxovirus serotype 1 (APMV-1), belongs to the genus Avulavirus, family Paramyxoviridae, and order Mononegavirales (37). It has a nonsegmented, single-stranded, negative-sense RNA genome that encodes six major structural proteins (from 3′ to 5′): nucleoprotein (NP), phosphoprotein (P), matrix protein (M), fusion protein (F), hemagglutinin-neuraminidase (HN), and RNA-dependent RNA polymerase (L) (7,16).
M protein has 364 amino acids (aa) with a molecular weight of ∼40 kDa (3,10). It is an abundant protein that interacts with the host cell membrane, viral RNP, and glycoprotein in the inner cytomembrane to promote virus particle assembly and budding (27,38). It is also virulence related and inhibits host cell RNA transcription and protein translation (1,26). In addition, M protein completes import and export from the nucleus through one nuclear localization signal (NLS) and three nuclear explore signals (NES) to assist with the reproduction of virus particles (11,13,14).
M protein elicits considerable serum antibody titers after the immunization of chickens with NDV (17), and anti-NDV chicken serum can recognize M protein in virus-like particles (29). These reports indicate that viral M protein is able to induce strong humoral immunity during NDV infection in animals, even though the M protein-specific antibody has no neutralization activity against NDV (30). However, the antigenic determinants of M protein remain elusive.
Epitope characterization is crucial for the understanding of antibody functions in recognition and binding (5,25). Detailed analyses of epitopes are essential for understanding immunological events and for the development of diagnostic tools for various diseases and epitope-oriented vaccines (21,33). Furthermore, the M epitopes would be useful as potential markers that can be modified in recombinant virus for the differential immunodiagnosis in differentiating infected from vaccinated animals (DIVA) vaccine.
The pepscan technique, also known as overlapping peptide scan or array-based oligopeptide scanning (12), has facilitated the identification of a linear epitope in the fusion (F) protein of bovine respiratory syncytial virus (BRSV) characterized by neutralization activity, which made it feasible to develop an epitope-based vaccine (19). A pepscan analysis has also been used to identify linear B cell epitopes on the NP protein of NDV to develop a marker vaccine candidate (23). This epitope mapping technology is fast and low cost for practical applications (28,34). Briefly, it involves the use of an array of unique, overlapping synthetic peptides based on the primary amino acid sequence of an antigen bound to individual pins to detect serologically accurate epitopes.
In this study, two accurate linear B cell epitopes, 77MIDDKP82 and 354HTLAKYNPFK363, of NDV M protein were identified by a pepscan analysis and site-directed mutagenesis. Our results clarify antigenic properties of the M protein and could genetically eliminate these immunodominant epitopes (IDEs) in recombinant virus that serve as negative markers for the development of DIVA or marker vaccines for NDV.
Materials and Methods
Cells and virus
HEp-2, DF-1, and BHK21 lines were obtained from the ATCC (Manassas, VA) and cultured in Dulbecco's modified Eagle's medium (DMEM)/high glucose (Hyclone, Thermo Scientific, Logan, UT) in a humidified 5% CO2 atmosphere at 37°C. All culture media were supplemented with 10% inactivated fetal bovine serum (GIBCO, Invitrogen, Gaithersburg, MD) and antibiotics (100 U/mL penicillin and 100 mg/mL streptomycin). The F48E9 strain of NDV was obtained from the China Institute of Veterinary Drug Control. The inactivated and attenuated LaSota vaccines were purchased from Yebio (Qingdao, China). The strains SX10, JS17, and Pigeon/QH were isolated from China in our laboratory.
Preparation of anti-LaSota chicken hyperimmune serum and anti-IDE peptide mouse antisera
Four 4-week-old specific pathogen-free (SPF) white leghorn chickens (SAIS Poultry) were raised in animal isolators. They were inoculated through the intraocular–nasal route with 0.2 mL per chicken of LaSota live vaccine on day 1. Then, the chickens were immunized with 0.2 mL per chicken of inactivated vaccine through subcutaneous injection on days 14 and 28. Blood samples were collected at day 35, and the antisera were prepared.
Three 6-week-old SPF BALB/c mice were immunized by subcutaneous injection with 100 μg per mouse of each synthetic IDE peptide emulsified by complete Freund's adjuvant (Sigma, St. Louis, MO) in phosphate-buffered saline (PBS) at day 1. Then, the mice were boosted with each peptide emulsified by incomplete Freund's adjuvant (Sigma) at days 14 and 28. Finally, the blood samples were collected at day 38. The antisera of selected IDEs (IDE1, IDE2, IDE5, and IDE6) were prepared. All experimental procedures were approved by the Animal Care and Use Committee of the Northwest A&F University, China (Approval No. 2015ZX0512008-012 to chicken, No. 2016XN0928010-068 to mouse).
Pepscan analysis
The pepscan analysis was performed by the Novasnow Science & Technology China with anti-LaSota hyperimmune chicken serum. The matrix protein sequences were linked and elongated with neutral GSGSGSG linkers at the C- and N-termini to avoid truncated peptides. The elongated antigen sequences were translated into 15 amino acid peptides with a peptide–peptide overlap of 14 amino acids. The resulting PEPperCHIP® Peptide Microarrays contained a series of different peptides printed in duplicate and framed by additional HA (YPYDVPDYAG, 100 spots) and c-myc (EQKLISEEDL, 100 spots) control peptides.
After 15 min of preswelling in washing buffer and 30 min of incubation in blocking buffer, peptide microarrays were initially incubated with the secondary antibody goat anti-chicken IgG (H+L) DyLight680 (1:5,000) and with control antibody mouse monoclonal anti-HA (12CA5) DyLight800 (1:2,000) for 45 min at 37°C to analyze background interactions with the antigen-derived peptides. Then, the peptide microarray analysis was performed with anti-LaSota chicken hyperimmune serum diluted at 1:500 in incubation buffer for 16 h at 4°C and shaking at 140 rpm. The LI-COR Odyssey Imaging System was used to scan the preprocessed peptide microarrays at scanning intensities of 7/7 (red = 700 nm/green = 800 nm).
Construction of the truncated M protein, IDE-RFP, and IDE mutants
To confirm whether NDV-specific antisera can recognize viral M protein, we constructed the M protein and its truncations. M protein was divided into two parts, aa1–177 (M1) and aa178–365 (M2), based on B cell epitope prediction using the Immune Epitope Database and DNAstar-protean (Fig. 1A).Viral RNA was extracted from the purified NDV using TRIzol according to the manufacturer's protocol (TaKaRa, Kusatsu, Japan). The extracted RNA was reverse transcribed with random primers to generate complementary DNA (cDNA) of LaSota. The fragments of the M, M1, and M2 genes were amplified by polymerase chain reaction (PCR) (GenStar, Beijing, China) with the primers 1 and 2, primers 1 and 3, and primers 4 and 2, respectively (Supplementary Table S1). The reverse transcription-polymerase chain reaction (RT-PCR) products were cloned into the pMD19-T vector (TaKaRa) and sequenced (GENEWIZ, South Plainfield, NJ). The gene fragments in the pMD-19T vectors were digested with the restriction enzymes EcoRI and BglII (Thermo, Waltham, MA). Then, they were cloned into the eukaryotic expression vector pCAGGS with T4 ligase (TaKaRa) to obtain the constructs pCAGGS-M, pCAGGS-M1, and pCAGGS-M2.

Pepscan analysis of M protein in the LaSota strain.
Six IDE fusions with red fluorescent protein (RFP) were constructed by overlapping PCR with primers for IDE-RFP (Supplementary Table S2). Upstream primers were combined with IDE sequences and the 3′ terminus of the RFP gene. The downstream primer was located at the 5′ terminus of the RFP gene. Each PCR product was cloned into pMD-19T for sequencing. Then, the IDE-RFP fragments were digested by restriction enzymes, XhoI and BglII, and ligated into pCAGGS vectors.
MIDE2 and MIDE6 mutants were generated by overlapping PCR (Supplementary Tables S3 and S4). Amino acids at the mutated sites were replaced with alanine. The fragments of MIDE2 and MIDE6 mutants fused with RFP were cloned into pCAGGS with the restriction enzymes XholI and BglII.
Immunofluorescence assay
Cells in 12-well plates were fixed with 4% paraformaldehyde for 10 min at 4°C. Then, they were washed three times with PBS and incubated for 1 h at 37°C with primary antibodies. After washing three times with PBS, the cells were incubated with fluorescein isothiocyanate (FITC)-conjugated anti-chick antibody (Abcam, Cambridge, United Kingdom) for 1 h at 37°C and then further washed with PBS. The cells were observed using an inverted fluorescence microscope (Olympus, Tokyo, Japan).
Western blotting
The 293T cells were transfected by TurboFect (Thermo Scientific) with expression plasmids. After 48 h posttransfection, the cell lysates were prepared with 2 × sodium dodecyl sulfate (SDS)-loading buffer and subjected to 12% SDS–polyacrylamide gel electrophoresis (SDS-PAGE) after denaturation at 100°C for about 10 min. After gel electrophoresis, the proteins were transferred to nitrocellulose membranes (Millipore, Billerica, MA), and blocked in 5% skim milk in PBS at 4°C overnight. After washing with PBST (0.5% Tween-20 in PBS), the membrane was incubated with primary antibodies for 2 h at 37°C. After washing, the protein bands were detected with an horseradish peroxidase (HRP)-conjugated IgG antibody (Abcam).
Dot blot
The IDE peptides (15 amino acids) were synthesized by Ontores Biotechnology (Shanghai, China). Each peptide (2 μg) was dropped onto the nitrocellulose membrane and incubated at 37°C for 15 min. The membrane was blocked with 5% skim milk at 37°C for 2 h. After washing three times with TBS, the membrane was incubated with the LaSota antiserum or anti-IDE antibody at 37°C for 1 h. After washing with TBST, the membrane was incubated with the corresponding HRP-conjugated secondary antibody. The ECL peroxidase substrate (Millipore) was used for detection.
Enzyme-linked immunosorbent assay
A 96-well plate was coated with synthesized epitopes in 0.1 M carbonate buffer (pH 9.6) and incubated at 4°C overnight. After washing three times with PBST, plates were blocked with 5% skim milk for 2 h at 37°C. After washing three times with PBST, a rabbit anti-mouse HRP-conjugated polyclonal serum diluted 1:5,000 with 5% skim milk (Abcam) was added and incubated for 1 h at 37°C. After washing, TMB (Solarbio, Beijing, China) was added to the wells as a substrate for HRP and incubated in the dark for 15 min. Then, 2 M H2SO4 was added to stop the reaction and absorbance was read at 450 nm using a Microplate Absorbance Reader (Bio-Rad, Hercules, CA). Data are presented as the average of triplicate assays.
Conservation analysis of IDEs
To determine whether IDE amino acid sequences are conserved in the M protein of NDV strains, the sequences of NDV strains belonging to different genotypes were downloaded from GenBank, and sequence alignments among them were generated using DNAstar MegAlign.
Results
Epitope mapping by peptide scanning
NDV infection or vaccination can induce a humoral immune response in chickens. First, to test whether the M protein of NDV can be recognized by NDV-specific chicken antiserum, the 293T cell lysates transfected with the M constructs were analyzed by western blotting using chicken hyperimmune NDV-specific antiserum prepared by LaSota vaccination. The NDV-specific antisera was able to recognize the expressed M, M1, and M2 proteins (Fig. 1B). This result was confirmed by an immunofluorescence assay (IFA) using transfected BHK21 cells (Fig. 1C). Subsequently, the NDV-specific antiserum was used to scan the linear B cell epitopes of M protein by the pepscan approach. Six epitopes with high reactivity in the intensity plot were selected as IDE candidates, marked MIDE1 to MIDE6 from the N-terminus to C-terminus of the M protein (Fig. 1D). Each IDE contained 15 amino acids (Table 1).
Six Potential Epitopes Identified by Pepscan
aa, Amino acid.
Identification of IDEs
To express the IDEs in mammalian cells, the six candidates were each fused to RFP. MIDE2-RPF and MIDE6-RFP were clearly detected by IFA with NDV-specific antisera in the transfected BHK21 cells, but the others were not (Fig. 2A). Similarly, MIDE2-RFP and MIDE6-RFP were positively detected by western blotting (Fig. 2B). These results indicated that the epitopes MIDE2 and MIDE6 could be recognized by NDV-specific antisera.
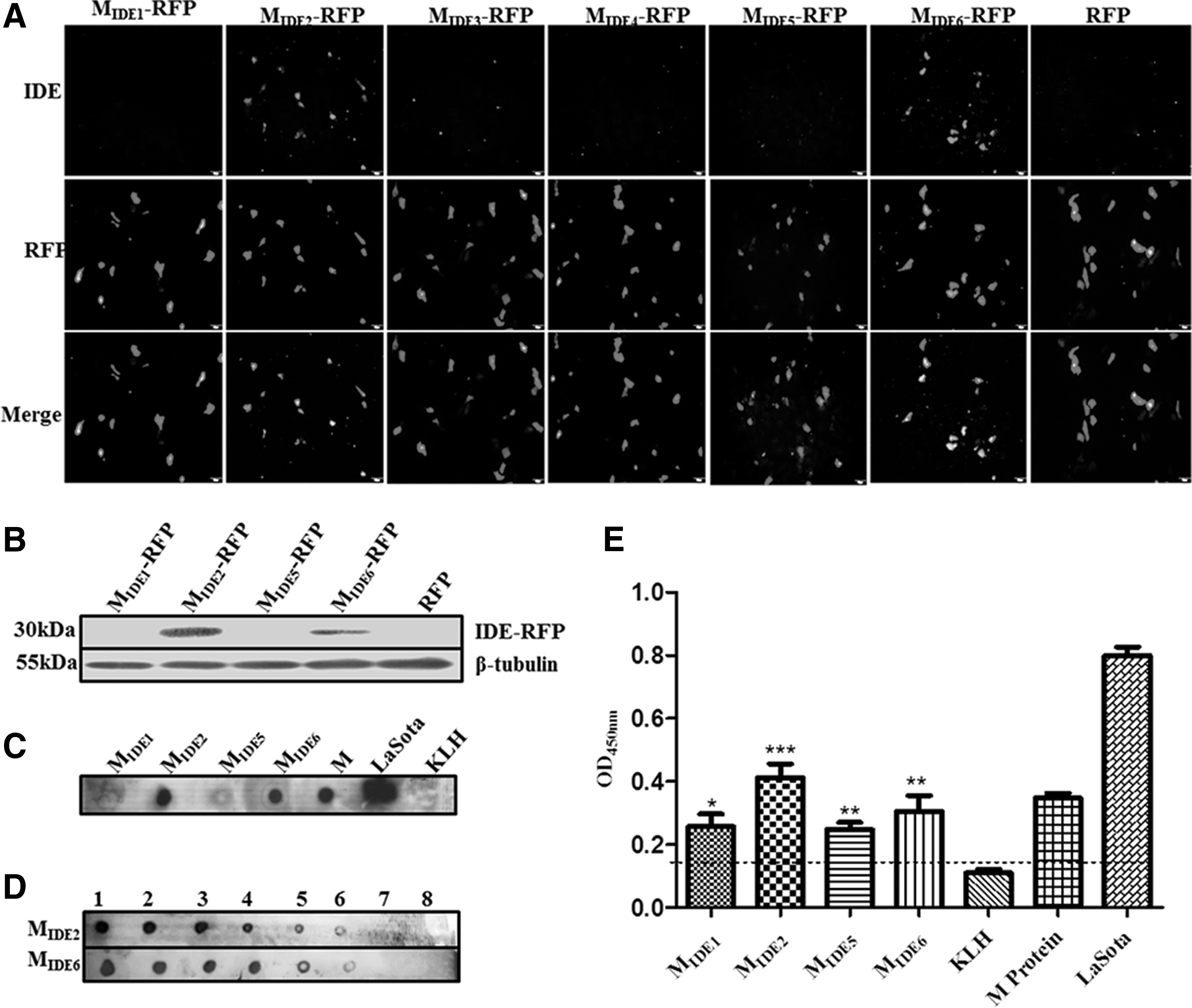
Identification of IDEs.
To confirm the IDEs, the peptides of four epitope candidates conjugated to KLH were synthesized. Using a dot-blot analysis, MIDE2 and MIDE6 were obviously detected with NDV-specific antisera and MIDE5 was slightly detected (Fig. 2C). The serial dilution of synthetic MIDE2 and MIDE6 showed a reduction in reactivity (Fig. 2D), indicating that both IDEs were highly sensitive to NDV-specific antisera. In addition, enzyme-linked immunosorbent assay further revealed that MIDE2 and MIDE6 exhibit high reactivity, even though MIDE1 and MIDE5 were also positively detected (Fig. 2E). Taken together, these results suggested that the MIDE2 at residues 71–85 (M71–85) and MIDE6 at residues 349–363 (M349–363) are IDEs of the M protein.
Immunogenicity of the IDE peptides
To examine the immunogenicity of the IDEs, anti-IDE mouse antisera were prepared. Using a dot-blot assay, antisera against MIDE1, MIDE2, MIDE5, and MIDE6 were able to recognize the LaSota virus and M protein. The antisera of MIDE2 and MIDE6 strongly reacted with their own peptides MIDE2 and MIDE6, respectively. However, the antisera against MIDE1 and MIDE5 were unable to recognize their own peptides (Fig. 3A). Antisera to the IDE peptides did not exhibit crossreactivity with each other. Moreover, these data were confirmed by western blotting (Fig. 3B). These results indicated that MIDE2 and MIDE6 could specifically elicit a humoral response.
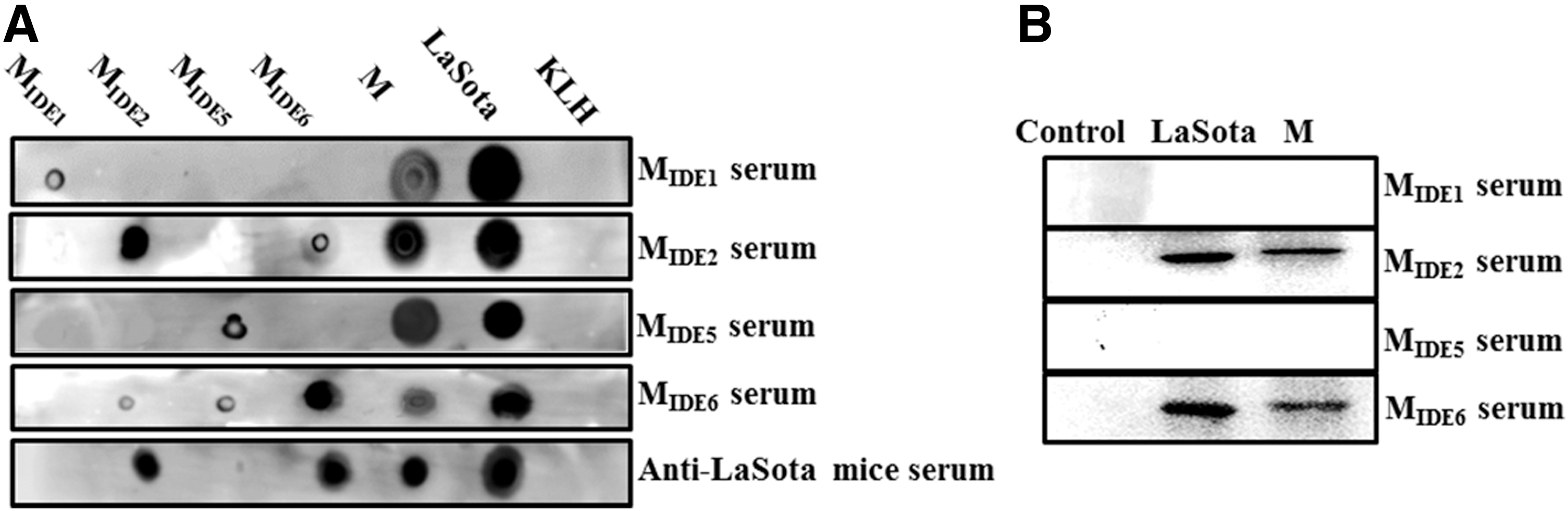
Immunogenicity of the IDE peptides.
Minimization of MIDE2 and MIDE6
To examine whether all 15 amino acids of MIDE2 71EEATVGMIDDKPKRE85 and MIDE6 349KLEKGHTLAKYNPFK363 are required (i.e., to identify minimal epitopes), a series of amino acid mutations of MIDE2 and MIDE6 fused to RFP were constructed and expressed in transfected 293T cells. Based on detailed data from a pepscan analysis (Supplement 2), the residues 71–74 and 83–85 of MIDE2 as well as 349–351 of MIDE6 were not important for the two epitopes. Therefore, MIDE2 75VGMIDDKP82 and MIDE6 352KGHTLAKYNPFK363 were analyzed by mutagenesis. The MIDE2 N-terminal mutants AAAIDDKP (V75A, G76A, M77A) and AAAADDKP (V75A, G76A, M77A, I78A) exhibited significantly decreased or a lack of recognition by anti-MIDE2 mouse antiserum (Fig. 4A, left). The MIDE2 C-terminal mutants VGMIDDKA (K81A), VGMIDDAA (K81A, P82A), and VGMIDAAA (K81A, P82A, K81A) failed to be recognized by the anti-MIDE2 mouse serum (Fig. 4A, right). These data indicated that 77MIDDK82 was a minimal epitope of MIDE2 responsible for recognition by the antiserum. Similarly, the MIDE6 N-terminal mutants AAAALAKYNPFK and AAAAAAKYNPFK (Fig. 4B, left) as well as the C-terminal mutants KGHTLAKYNPFA, AAHTLAKYNPFA, and KGHTLAKYNPAA (Fig. 4B, right) exhibited dramatically reduced or failed reactions with anti-MIDE6 serum, suggesting that 354HTLAKYNPFK363 was a minimal epitope of MIDE6.

Minimization of MIDE2 and MIDE6.
Conservation of IDEs among strains
Among various strains with different genotypes, the linear epitope of MIDE2 71EEATVGMIDDKPKRE85 was mostly conserved. In the minimal epitope 77MIDDKP82, the residues 78I, 80D, and 82P were particularly highly conserved. MIDE6 was also conserved, and the region 358KYNPF362 had the highest conservation among selected 14 strains (Supplementary Tables S5 and S6). Furthermore, the M protein of the strains LaSota (genotype II), SX10 (genotype VI), JS17 (genotype VII), and F48E9 (genotype IX) of Class II as well as Pigeon/QH (Class I) were positively detected by western blotting with anti-MIDE2 (Fig. 5A) and anti-MIDE6 (Fig. 5B) mouse antisera in the infected DF-1 cell lysates, indicating that the MIDE2 and MIDE6 peptide antisera were able to recognize the M protein of multiple NDV strains. These results suggested that MIDE2 and MIDE6 of NDV were not only conserved with respect to amino acid sequences but also with respect to serological characteristics.
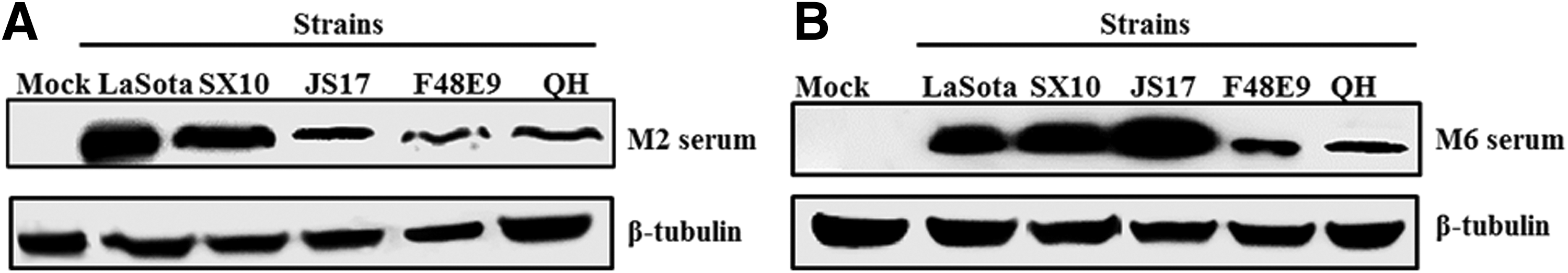
Serological recognition of the M protein in multiple NDV strains by anti-MIDE2 and anti-MIDE6 mouse antisera. DF-1 cells were infected with NDV strains. The cells were collected at 36 h for a western blot analysis with anti-MIDE2
Spatial location of the minimal MIDE2 and MIDE6 in M protein
We established the crystal model of LaSota M protein using SWISS-MODEL and performed a three-dimensional structure analysis using PyMOL (2), as shown in Figure 6. MIDE2 (aa77–82) and MIDE6 (aa354–363) were located at the random coil of the surface and were not in close proximity (Fig. 6). These characteristics suggest that they could feasibly form linear antigenic epitopes and have little interaction effects (18). These data further suggested that the refined motifs 77MIDDKP82 and 354HTLAKYNPFK363 are potential epitopes.

Three-dimensional localization of the minimal MIDE2 and MIDE6 in the M protein structure. The spatial structure of M protein was visualized using PyMol (Schrodinger, Inc.). 77MIDDKP82 is MIDE2 and 354HTLAKYNPFK363 is MIDE6.
Discussion
The identification of epitopes, as antigenic determinants, is critical for understanding antibody recognition and binding (20,31). As an abundant viral protein of NDV, M protein elicits high antibody titers after vaccination or infection in poultry (17,29). Although the antibody of M protein has no neutralization activity against NDV (30), it would be useful for the differential immunodiagnosis of various strains. To identify the epitopes of M protein, we used pepscan technology to screen immunodominant linear B cell epitopes. This approach has been widely applied to many viral proteins (4,35,39). It has various advantages over other methods, for example, it has high-throughput systematic scanning ability, is rapid, and is inexpensive (22). Its disadvantages are that it fails to identify previously confirmed epitopes and short linear epitopes are often not recognized with high affinity by antibodies, along with low accuracy for scanned data (15). In our study, MIDE1, MIDE3, MIDE4, and MIDE5 failed to react highly with NDV-specific chicken serum, despite significant signal peaks in the pepscan analysis (Supplement 2). Only MIDE2 and MIDE6 were positively confirmed among six candidates by multiple methods with NDV-specific chicken serum.
The M protein plays important roles in the infection, assembly, and budding of NDV (2,6,26). The NLS and NES signals of M protein are required for nuclear shuttling in cells (13,14). The L-domain is critical for NDV budding (8). The arginine residue at position 42 (42R) of M protein contributes to NDV replication and infection (9). MIDE2 and MIDE6 identified in our study were distant from those critical regions or sites of M protein (Fig. 1A). Hence, these two epitopes may have no effect on the main functions of M protein.
The M protein is highly conserved in NDV strains (32,36). MIDE2 and MIDE6 were highly conserved in the sequence alignment of 14 NDV strains, especially 78–82 aa and 358–362 aa in M protein. Furthermore, mouse anti-MIDE2 and anti-MIDE6 sera have distinct reactivity with different strains. These data further confirm the conservation of M protein 71–85 aa and 349–363 aa with respect to serological characteristics. Interestingly, 79D, 81K, and 356L in the minimal epitopes were not conserved among the NDV strains. These residues likely do not affect M protein antigenicity. Moreover, 77M and 82P of MIDE2 and 354H, 355T, 362F, and 363K of MIDE6 were important for IDE2 antigenicity.
Conclusion
In this study, we identified two immunodominant linear B cell epitopes, MIDE2 77MIDDKP82 and MIDE6 354HTLAKYNPFK363, of NDV M protein using a pepscan approach with vaccinated chicken antisera. All peptides of six IDE candidates (Table 1) exhibited effective immunogenicity in animals. These data provide fundamental information about the antigenic features of M protein and these IDEs could be genetically eliminated as negative markers in recombinant virus for serologically differential diagnosis in the development of DIVA or marker vaccines for NDV.
Footnotes
Acknowledgments
This work was supported by the Faculty Support Fund of the Northwest A&F University (grant No. Z111021401 to S.X.) and National Key Research and Development Program (grant No. 2017YFD0500702 to H.C.).
Authors' Contributions
Y.B., and S.X. designed the experiments. Y.B., Z.J., Y.W., and S.L. performed experiments. Y.B., S.M., W.W., Q.W., N.H., X.W., and Z.Y. analyzed the data. Y.B., S.X., and H.C. wrote and edited the article. All authors read and approved the final article.
Author Disclosure Statement
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work, there is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, the article entitled.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.