
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) still poses a threat to the swine industry worldwide. Currently, commercial vaccines against PRRSV, which consist of modified live or inactivated virus, reduce symptoms and viremia in immunized pigs, but efficacy against heterologous strains is variable. This has led to the development of subunit vaccines that contain viral antigens that show the highest variability. In this work, a chimeric protein comprising short amino acid sequences from glycoprotein 3 (GP3), glycoprotein 4 (GP4), glycoprotein 5 (GP5), and M (matrix protein) proteins of PRRSV was designed and expressed in Escherichia coli. This protein, designated as PRRSVchim, was purified by immobilized metal affinity chromatography and evaluated. PRRSVchim was identified by immunoglobulin G (IgG) presence in serum samples from PRRSV-positive pigs. Also, the protein probed to be antigenic in immunized mice and piglets and provided some degree of protection against challenge with a PRRSV field isolate. These results show the potential of PRRSVchim protein for both PRRSV diagnostic and immunoprophylaxis.
Introduction
Porcine reproductive and respiratory syndrome virus (PRRSV) is considered one of the first causes of infections in pig farming and the most important disease by the global swine industry, due to the annual economic losses ranging $560–663 million dollars (28). Previously known as the mystery disease, the porcine respiratory and reproductive syndrome (PRRS) was first described in 1987 in North America and in early 90's in Europe (51). The virus causes reproductive failures in sows, characterized by late abortions and births of weak or dead pigs (19). In piglets and young pigs, PRRSV causes respiratory disorders and delayed in the normal growth, predisposing to secondary microbial infections and contributing to the complex of porcine respiratory diseases in fattening pigs (10,44). The clinical manifestations of the disease depend on elements, such as the virus strain and host genetics. PRRSV is an enveloped, positive-strand RNA virus with icosahedral capsid, belonging to the Arteriviridae family into the Nidovirales order. The viral genome is about 15 kb and contains nine open reading frames (ORFs). The ORF1a and ORF1b constitute up to 75% of the size of the virus and the rest of the genome comprises seven smaller ORFs. The nonstructural polyproteins, pp1a and pp1ab, are translated from ORF1a and ORF1b and then proteolytically processed into 14 nonstructural functional proteins (nsps) (39). On the other hand, the low-abundant structural proteins, GP2a, GP2b, glycoprotein 3 (GP3), and glycoprotein 4 (GP4) are encoded by ORF2a, ORF2b, ORF3, and ORF4, whereas the three major structural proteins, glycoprotein 5 (GP5), M (matrix protein), and N, are derived from ORFs 5, 6, and 7, respectively (46). Two different genotypes of PRRSV have been reported to date: the genotype type 1 or “Lelystad” isolated in Europe, and the genotype type 2 or “VR2332” from North America (20,51). Currently, both genotypes are circulating worldwide and several strains of PRRSV have been found for both genotypes, showing differences in genome, sequence, antigenicity, and pathogenicity (20).
At present, there are two types of approved commercial vaccines against PRRSV: the attenuated virus vaccines (modified live vaccines) and the inactivated virus vaccines (killed vaccines). Although both kinds of vaccines decrease symptoms and viremia, they do not prevent from the infection, and crossprotection against heterologous viruses is variable (27). The attenuated virus vaccines against PRRSV have shown to be more effective than the inactivated vaccines (11), but the protection degree depends on the similarity in the genome sequences between the viral strain and the circulating or wild strain (11,22). Currently, several research groups have been working on the development of new vaccines against PRRSV with different strategies and outcomes.
DNA vaccines, resulting in symptoms reduction but without protection against the infection (6,31,43); subunit and peptide vaccines where most promising results came from subunit vaccines fused to antibodies specific to receptors or dendritic cells (41,42), chimeric and virus-vectored vaccines that contain proteins from bacteria or another viruses, for instance AHIN1, or Pseudomonas proteins, fused to PRRSV proteins (17,49), or even another viruses expressing PRRSV proteins (8); and finally plant-derived vaccines, where in some cases the administration route is by eating the transgenic plants expressing PRRSV antigens (1,3,32). However, generating an effective vaccine to prevent and control PRRSV infection has been challenging given the great diversity of viral subtypes generated for both PRRSV genotypes.
The most common transmission pathway is by oronasal route directly from another infected pig, or indirectly by fomites; but the infection in pigs also can be acquired during coitus, insemination, or even percutaneously during farming practices, including vaccination with reused needles (15,29). During oronasal exposure, infection begins in tonsils and the upper respiratory tract, where it has been observed that virus attenuates innate antiviral immune responses to the infection, resulting in a delayed or weak immunomodulatory response in the host (18).
Pigs may exhibit differences in the expression of proinflammatory cytokines depending on the PRRSV strain (23). Proinflammatory cytokines such as alpha interferon (IFN-α), tumor necrosis factor alpha (TNF-α), and interleukin-1 (IL-1) have been shown to play a key role in the severity of respiratory disease since the major target cells for virus replication are porcine alveolar macrophages (PAMs) (45).
Viral proteins have different roles both in infection and stimulating the immune system in the pig, resulting in the production of neutralizing and non-neutralizing antibodies. The GP5 and M proteins form a heterodimer that is essential in the formation of infectious viral particles. The GP5/M and GP2/GP3/GP4complexes are linked through noncovalent interactions between GP5 and GP4 (47). Several publications have shown that major neutralizing epitopes are found in the GP5 and GP4 proteins, which are also the proteins with the most variation among the different isolates (36,52), but more recent publications also have shown neutralizing epitopes to be found in M, GP3, and GP2 proteins (48). Finally, the N protein binds to the RNA viral genome to form the nucleocapsid. The N structural protein of PRRSV is also immunogenic, causing a strong antibody response. However, the antibodies do not neutralize the virus (5).
What we are presenting herein is the development of a novel chimeric protein comprising short sequences from PRRSV structural proteins. This chimeric protein designated as PRRSVchim, was expressed in Escherichia coli and further purified by immobilized metal affinity chromatography (IMAC). The PRRSVchim demonstrated during the activation of Raw-246.7 murine macrophages after 24 h an increased production of proinflammaroty cytokines, such as IL-6, TNF-α, anti-inflammatory cytokines as IL-10, and growth factor as GM-CSF compared with the isolated PRRSV. Also, this novel protein was assayed for its ability to be recognized by immunoglobulin G (IgG) from serum samples of PRRSV-infected pigs and for the IgG response that induces after being administered to piglets through the intramuscular route.
Materials and Methods
PRRSV chimeric protein design and plasmid construction
PRRSV chimeric protein was designed to include some of the sequences from structural proteins of the virus that are relevant for setting a protective immune response in pigs, as follows: two GP3 epitopes (gp3-epi1: QAATEIYEPGRS and gp3-epi2: LWCRIGYDRCEE), one GP4 epitope (gp4-epi: RDSASEAIR), and GP5 and matrix protein (M) ectodomains (gp5-ed: NASNDSSSHLQLIYNLTLCELNGTDWLANKFDWAV and m-ed: GSSLDDFCHDSTAPEKVL, respectively), each separated by a short linker of four alanines (Fig. 1). The sequences are based on the vaccine strain RespPRRS MLV genome (GenBank accession: AF066183.3), which belongs to the North American genotype, represented by the prototype virus isolate VR-2332.
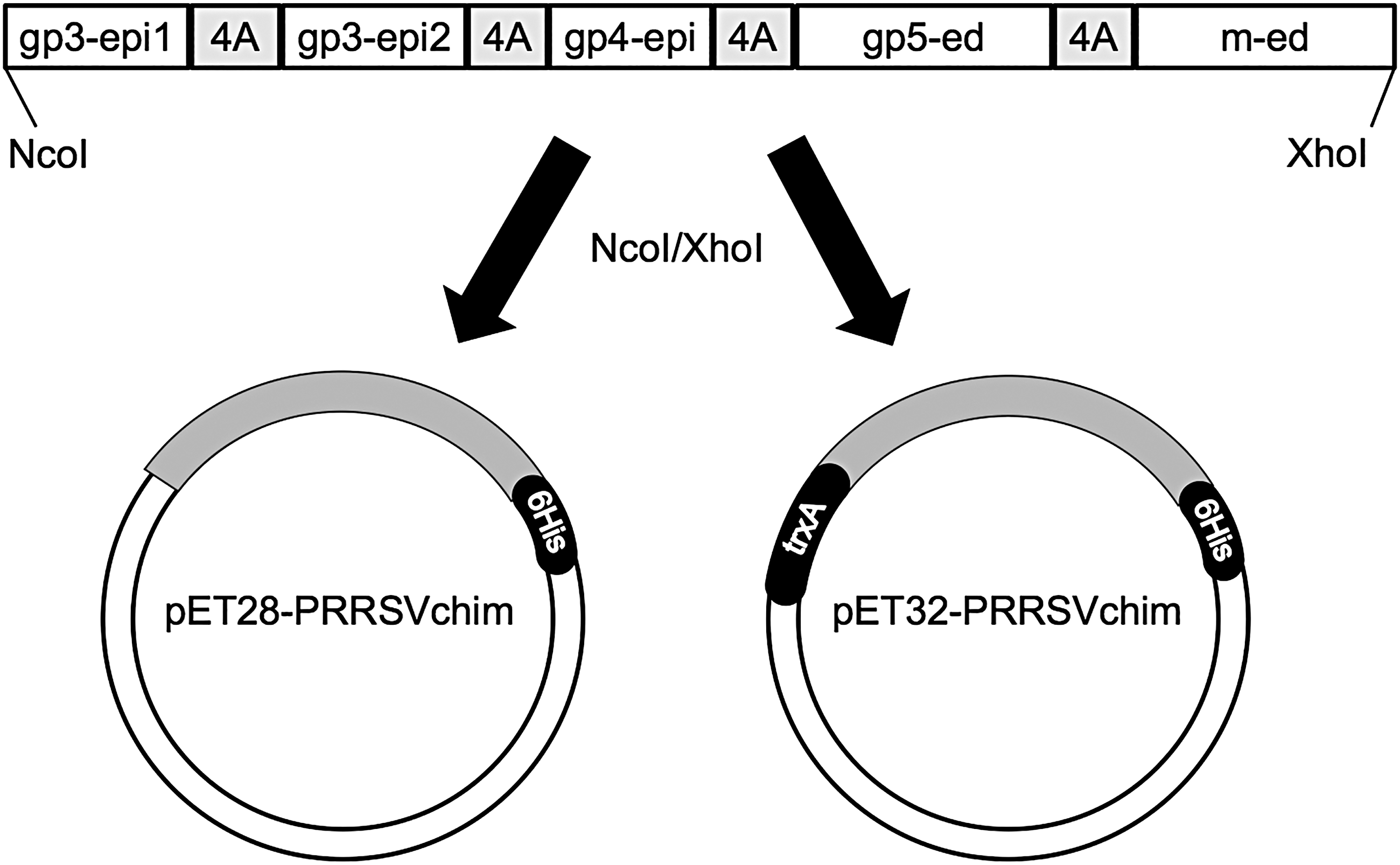
PRRSV chimeric protein design and cloning into bacterial expression plasmids. An optimized sequence comprising fragments from PRRSV structural proteins was synthesized and cloned into T7 bacteriophage RNA polymerase-based bacterial expression plasmids, pET28a+ and pET32a+, at NcoI and XhoI sites. 4A, four alanine spacer; 6His, hexahistidine tag; GP, glycoprotein; gp3 epi-1 and gp3 epi-2, GP3 protein epitopes; gp4 epi, GP4 protein epitope; gp5-ed, GP5 protein ectodomain; M-ed, M protein ectodomain; PRRSV, porcine reproductive and respiratory syndrome virus; trxA, thioredoxin fusion partner.
A synthetic sequence encoding PRRSV chimeric protein with NcoI and XhoI sites at 5′ and 3′ ends, respectively, was manufactured and inserted in pUC57 plasmid (Genscript). The synthetic sequence was excised with NcoI and XhoI restriction enzymes (New England Biolabs), electrophoresed in a Tris-acetate-EDTA agarose gel stained with SYBR-Safe (Thermo Fisher Scientific), cut, and purified with the MinElute Gel Extraction Kit (Qiagen) according to the provider's instructions. Later on, the synthetic sequence was ligated into NcoI/XhoI-digested pET28a+ and pET32a+ expression plasmids (Merck-Millipore) with T4 DNA ligase (Promega).
Ligation products were used to transform electrocompetent E. coli DH5-alpha cells by electroporation in a Gene Pulser II apparatus (Bio-Rad) and cells were plated onto LB/agar medium (Sigma-Aldrich) supplemented with 50 μg/mL kanamycin or 50 μg/mL ampicillin (Thermo Fisher Scientific) for selecting colonies harboring pET28a+ or pET32a+ constructs, respectively. Plasmid DNA from a few colonies was extracted with the GeneJet Plasmid Miniprep Kit (Thermo Fisher Scientific) and analyzed by restriction pattern with NcoI and XhoI enzymes with Gel Doc EZ System (Bio-Rad). New plasmid constructs were designated as pET28-PRRSVchim and pET32-PRRSVchim (Fig. 1).
PRRSV chimeric protein expression
The new plasmid constructs were first mobilized into E. coli BL21 Star (DE3) expression strain by electroporation, and a few colonies growing on LB/agar medium selective plates were checked for plasmid presence by restriction analysis. Single colonies were picked, inoculated into 15-mL polypropylene conical screw cap tubes (Neptune Scientific) containing 2 mL of LB liquid medium (Sigma-Aldrich) supplemented with the appropriate antibiotic, and overnight grown at 37°C/250 rpm. Then, 0.25 mL of each incubated cultures were transferred to 10 mL of fresh LB liquid medium in 50-mL polypropylene conical screw cap tubes (Neptune Scientific) and incubated at 37°C/250 rpm.
When cultures reached an OD600 between 0.6 and 0.8, they were split in half (5 mL) in 50-mL tubes, isopropyl-β-D-1-thiogalactopyranoside (Promega) was added to one of the halves at a final concentration of 1 mM, and cultures were incubated 37°C/250 rpm/4 h. Subsequently, cultures were spun 4,000 × g/4°C/30 min, supernatants discarded, and cell pellets resuspended in 0.5 mL of phosphate-buffered saline (PBS) at pH 7.4. A given volume of resuspended cells was mixed with same volume of protein sample buffer containing 10% 2-mercaptoethanol, samples were boiled for 5 min and subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) in 12% acrylamide gels, using Mini-PROTEAN Tetra System (Bio-Rad).
Gels were stained with Coomassie Brilliant Blue. All protein electrophoresis reagents were purchased from Bio-Rad. To determine whether the chimeric protein was accumulating in the soluble or insoluble fraction, cell pellets were lysed by short sonication pulses in PBS pH 7.4 containing 0.1% Triton X-100 (Sigma-Aldrich) using a Microson XL-2000 sonicator (Misonix), lysate was centrifuged 17,000 × g/room temperature/5 min, supernatant was transferred to a new tube, and both supernatant and pellet were mixed with protein loading buffer and electrophoresed.
Western blots were conducted to confirm the expression of chimeric PRRSV protein. For this, proteins were transferred to 0.2 μm nitrocellulose membrane (Bio-Rad) by semidry electrotransference. Membrane was blocked 1 h/room temperature with PBS pH 7.4 buffer containing 5% nonfat milk powder and 0.1% Tween-20. After that, membrane was briefly washed with PBS pH 7.4 buffer/0.1% Tween-20 (wash buffer) and incubated overnight/4°C with anti-His mouse monoclonal antibody (Roche) diluted 1:500 in PBS buffer/2.5% nonfat milk/0.05% Tween-20. Membrane was washed five times with wash buffer, anti-mouse IgG polyclonal antibody-horseradish peroxidase conjugate (Bio-Rad) diluted 1:1,000 in PBS buffer/0.05% Tween-20 was added and incubated 2 h/room temperature. Membrane was again washed five times and HRP color development reagent (Bio-Rad) was added according to the manufacturer's instructions. Finally, reaction was stopped with tap water and membrane was left to dry.
PRRSV chimeric protein purification
Based on the initial results of the chimeric protein expression, the following assays were performed only with the pET32-PRRSVchim plasmid/clone/culture. An overnight culture (3 mL) from a fresh colony bearing pET32-PRRSVchim plasmid was inoculated into a 1,000-mL Erlenmeyer flask containing 250 mL LB Lennox medium (Sigma-Aldrich) and incubated 30°C/250 rpm until OD600 reached 0.8. Protein expression was induced with 1 mM IPTG for 4 h and cells were pelleted by centrifugation 4,000 × g/4°C/30 min. Pellets were resuspended in 25 mL of lysis buffer (20 mM Tris pH 8.0, 500 mM sodium chloride, 0.1% Triton X-100) and sonicated with repeated medium strength 10 sec pulses. Lysate was cleared by filtering through a 0.45 μm Millex-HV syringe filter unit (EMD Millipore) and imidazole pH 8.0 solution was added to cleared lysate to a final concentration of 20 mM.
PRRSV chimeric protein purification was conducted by IMAC in HisTrap HP 1 mL column (GE Healthcare Life Sciences) as follows. Twenty-five column volumes of lysate were injected through the column; afterward, column was washed with 25 volumes of washing buffer (20 mM Tris pH 8.0, 500 mM sodium chloride, 50 mM imidazole) and protein was eluted with 5 column volumes of 20 mM Tris pH 8.0, 500 mM sodium chloride, and 500 mM imidazole solution in 1 mL fractions. Selected fractions from the purification steps were analyzed by SDS-PAGE. Finally, elution fractions with highest recombinant chimeric protein content were pooled, underwent buffer exchange to PBS pH 7.4 with Amicon Ultra-4 centrifugal filter unit (EMD Millipore), and finally the purified protein was stored at 4°C.
PRRSV chimeric protein crossreactivity assay
To assess crossreactivity against the PRRSV chimeric protein, 36 positive and four negative pig sera samples were used; these samples were from individuals located in pig farms from Western Mexico that had been previously screened for detection of IgG antibodies against PRRSV, using the Indirect Enzyme-Linked Immunosorbent Assay (ELISA) IDEXX PRRS X3 Kit (IDEXX Laboratories), according to the manufacturer's instructions,. MaxiSorp flat bottom 96-well plates were coated overnight/4°C with PRRSV chimeric protein (1 μg/well) using coating buffer (10 mM carbonate buffer pH 9.6) in a 100 μL volume. Afterward, plates were washed three times with 300 μL/well of wash buffer (10 mM phosphate buffer pH 7.4, 0.05% Tween-20).
Pig serum samples were diluted 1:100 in wash buffer, 100 μL added to the wells, and plates were incubated at 37°C/1 h. Positive and negative controls from the ELISA IDEXX PRRS X3 Kit were also included. Plates were again washed three times in 100 μL/well of conjugate solution containing anti-pig IgG horseradish peroxidase conjugate (Sigma-Aldrich), diluted 1:5,000 in 10 mM phosphate buffer pH 7.4, 2.5% nonfat dry milk and 0.05% Tween-20 were added, and plates were incubated at 37°C/1 h. A further wash step was performed, 3–3′, 5–5′ tetrametylbenzidine substrate solution (Sigma-Aldrich) was added to the wells (100 μL), plates were incubated 3 min in the dark, and color development was stopped by adding 100 μL of 1 N sulfuric acid solution. Absorbance was measured at 450 nm in a Xmark microplate absorbance spectrophotometer (Bio-Rad).
Murine macrophage cell line stimulation assay
The response of a murine macrophage cell line to PRRSV chimeric protein was assessed. RAW 264.7 cell line from American Type Culture Collection (ATCC) was propagated in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, 1 mM sodium pyruvate, 2 mM
An isolated virus (VR2332c) was used for comparison purposes and added to the cells at a multiplicity of infection of 0.1. Culture medium containing only macrophages without PRRSV or PRRSVchim protein was used as negative control. Supernatants were stored at −20°C for further analysis. Cytokines, chemokines, and growth factor levels (IL-6, TNF-α, IL-10, MCP-1, RANTES, and GM-CSF) from the supernatants were measured in a Milliplex mouse cytokine plate (Merck Millipore) according to the manufacturer's instructions. The plate was read in a Luminex® xMAP® Technology instrument using xPONENT software.
Mice and pig immunization with PRRSV chimeric protein
All applicable international, national, and institutional guidelines for the care and use of animals were followed. The protocol was approved by the Internal Committee for the Care and Use of Laboratory Animals of Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, with the reference number 2015-006, following federal standards such as NOM-033-ZOO-1995 and NOM-062-ZOO-1999.
Twelve 4–6-week-old female BALB/c mice were acquired from Harlan Laboratories and separated in three groups of four individuals each. They were injected by the subcutaneous route with 10 μg PRRSVchim, 10 μg PRRSVchim/Freund's incomplete adjuvant, or PBS in a total volume of 100 μL on days 1 and 14. Blood samples were collected from tail vein on days 0 and 28, incubated 37°C/1 h, centrifuged 17,000 × g/5 min/4°C, serum was collected in new tubes, and stored −80°C for further analysis.
Serum samples collected were tested for total IgG and isotypes IgG1, IgG2a, and IgG2b antibody response against PRRSVchim by ELISA as described above with a few modifications. After coating, plates were blocked with 250 μL of PBS, 0.05% Tween-20, and 5% nonfat dry milk for 3 h at room temperature. Samples from each group were pooled and diluted 1:20 followed by serial 1:5 dilutions in blocking buffer. One hundred microliters of the appropriate secondary antibody, HRP-donkey anti-mouse IgG (Bio-Techne) diluted 1:1,000 or HRP-goat anti-mouse IgG subclass 1, 2a, or 2b (Jackson ImmunoResearch Laboratories) diluted 1:5,000 using PBS, 0.05% Tween-20, and 2.5% nonfat dry milk, were added and incubated for 3 h at room temperature. Color development was monitored before saturation.
Twenty-five piglets with a mean weight of 6.48 ± 1.61 kg were weaned 21 days after birth, acclimated for 5 days, and randomly divided into five groups of five individuals each. Groups were as follows: (1) nonimmunized/nonchallenged, (2) nonimmunized/challenged, (3) Ingelvac PRRS MLV commercial vaccine (Boehringer Ingelheim), (4) 100 μg PRRSVchim adjuvanted with Montanide ISA 15 A VG (Seppic), (5) 100 μg PRRSVchim adjuvanted with polymeric gel 01 (Seppic) and (6) 100 μg PRRSVchim without adjuvant. All animals were injected intramuscularly with a 2 mL volume. Nonimmunized group received physiological saline solution. All but the commercial vaccine group received a boost 14 days after the first immunization. Blood samples were taken on days 0, 14, 21, 28, and 42 (2 weeks after virus challenge) after priming from external jugular vein and serum samples were assayed for PRRSV-specific IgG with the commercial PRRS Ab ELISA 4.0 (Bionote), following the manufacturer's instructions, and PRRSVchim-specific IgG as described below.
Ninety-six-well MaxiSorp plates (Nunc) were coated with PRRSVchim protein (2.5 μg/well) overnight/4°C in 100 μL of 10 mM carbonate buffer pH 9.6. Then, plates were blocked with 10 mM phosphate buffer pH 7.4, 0.05% Tween-20, and 5% bovine serum albumin (BSA; 300 μL/well), and incubated at 37°C/1 h. Plates were washed thrice with 300 μL/well of wash buffer (10 mM phosphate buffer pH 7.4, 0.1% Tween-20). Afterward, serum samples were diluted 1:20 in blocking buffer, added to the plates (100 μL well), and these were incubated at 37°C/2 h. Plates were washed thrice with wash buffer.
Anti-porcine IgG-horseradish peroxidase conjugate (Sigma-Aldrich) diluted 1:5,000 in 10 mM phosphate buffer pH 7.4, 0.05% Tween-20, and 2.5% BSA was added (100 μL/well) and plates incubated at 37°C/1 h. Plates were again washed as described above and 100 μL of 3–3′, 5–5′ tetramethylbenzidine substrate (Sigma-Aldrich) was added to each well. After a 5 min incubation in the dark, the reaction was stopped with 100 μL 1 N sulfuric acid. Absorbance was read at 450 nm in a xMark microplate spectrophotometer (Bio-Rad).
Pig challenge experiment
A PRRSV field isolate from La Piedad, Michoacán, México, was employed for challenging the pigs. This isolate belongs to the North American VR2332 lineage, as confirmed by ORF5 sequencing, and was grown in MARC-145 cells. Each specimen received a 1 mL intranasal and intramuscular dose of 1 × 107 TCID50 of the field isolate 29 days after the first immunization. Pigs were sacrificed humanitarily at day 42 after priming. The duplex reverse transcription (RT)-insulated isothermal polymerase chain reaction (RT-PCR) POCKIT PRRSV-NA test, which is a qualitative presence/absence test that targets the North American lineage of PRRSV ORF6 and ORF7 genes (limit of detection of 10 copies), was used for detecting virus in nasal swabs, sera, and organ pools (lungs, tonsils, lymph node, and spleen). Also, rectal temperature, ocular secretion, and weight were registered for all groups.
Results
PRRSV chimeric protein design and plasmid constructs
The coding sequence of PRRSVchim protein, comprising GP3 and GP4 epitopes and GP5 and M ectodomains of PRRSV, based on the sequence of vaccine strain RespPRRS MLV, was in vitro synthesized and cloned into T7-based pET28a+ and pET32a+ plasmids. The hypothetical chimeric protein generated from pET28a+ plasmid, named pET28-PRRSVchim, would have a histidine tag at its carboxy terminus, whereas that generated from pET32a+ plasmid, designated as pET32-PRRSVchim, would have a thioredoxin fusion partner and a histidine tag at its amino and carboxy termini, respectively (Fig. 1).
PRRSV chimeric protein expression and purification
For PRRSVchim protein expression from pET28a+ and pET32a+ plasmids, E. coli BL21(DE3) strain was used. Lysates from both uninduced and IPTG-induced bacterial cultures were analyzed by SDS-PAGE (Fig. 2A). No differences were observed in the pattern of protein bands from uninduced and induced cultures carrying pET28-PRRSVchim. In contrast, a protein band corresponding to the estimated size of PRRSVchim generated from pET32-PRRSVchim plasmid (29.4 kDa), was clearly observed in both uninduced and induced lysates, but its accumulation was higher in induced than in uninduced lysates. PRRSVchim protein expression was confirmed by western blot, using a monoclonal antibody that binds the histidine tag of recombinant proteins (Fig. 2B). PRRSVchim protein accumulated at equal levels in soluble and insoluble protein fractions and this accumulation took place to a higher extent in IPTG-induced cultures than in uninduced ones (Fig. 2C). After this finding, PRRSVchim protein derived from pET32-PRRSVchim plasmid was expressed to a 250-mL culture medium scale in Erlenmeyer flasks for its further purification through IMAC. The protein eluted almost completely in the first two fractions; however, some contaminant proteins were present in these fractions (Fig. 2D). This way, a second round of IMAC purification of the protein was performed, using the same method as in the first round, which helped in obtaining a purer chimeric protein that was utilized for subsequent experiments. The estimated yield of this protein was about 15 mg/L of culture medium.

PRRSV chimeric protein expression in Escherichia coli and purification.
PRRSV chimeric protein interaction with IgG from serum samples of PRRSV-infected pigs
To determine whether PRRSVchim protein preserves the antigenic properties of its short amino acid fragments that originate from GP3, GP4, GP5, and M proteins of PRRSV, the interaction of this protein with IgG from serum samples of PRRSV-infected specimens located in pig farms in Western Mexico was assessed by ELISA, coating the microplate with PRRSVchim protein. The 36 samples that had been tested positive for PRRSV-specific IgG using a commercial ELISA Kit also reacted with PRRSVchim protein, with a 0.3 absorbance cutoff value (Fig. 3). Additionally, negative pig serum samples were below this cutoff value.
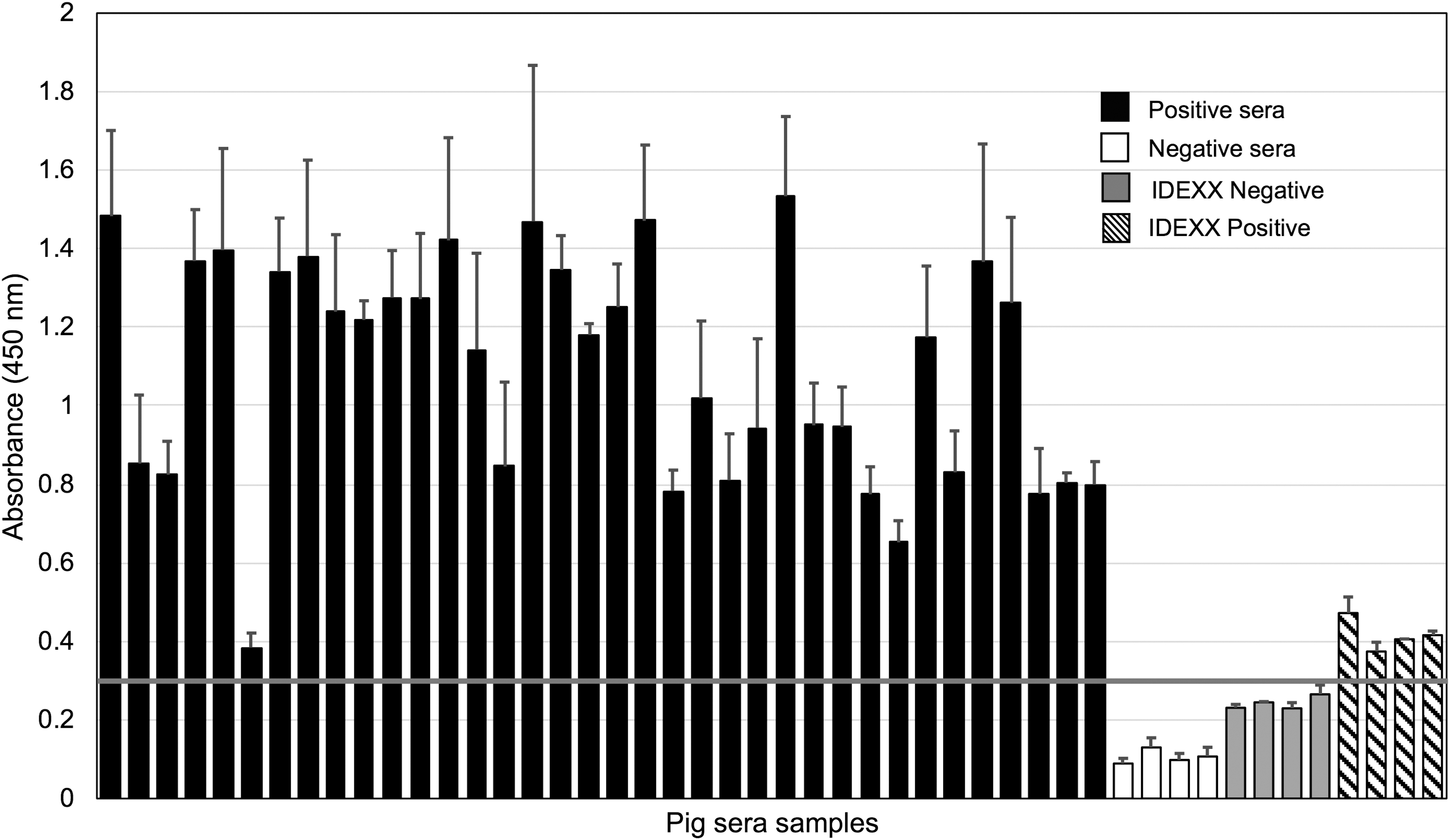
Interaction of serum IgG from PRRSV-infected pigs with PRRSV chimeric protein assessed by ELISA. PRRSV-positive (positive) and negative (negative) pig field serum samples, as well as positive (positive IDEXX) and negative (negative IDEXX) controls from a commercial kit that detects PRRSV-specific IgG were assayed for their interaction with PRRSV chimeric protein. The horizontal line indicates the cutoff value. ELISA, enzyme-linked immunosorbent assay; IgG, immunoglobulin G.
Cytokine secretion profile of a murine macrophage cell line exposed with PRRSV chimeric protein
To obtain a global perspective of the cytokine and chemokine profiles that PRRSVchim protein could stimulate, the murine macrophage cell line Raw 264.7 was used for in vitro assays. Also, a PRRSV isolate was included as treatment on this cell line. The analysis showed that PRRSVchim protein stimulated murine macrophages to secrete IL-6 and TNF-α cytokines at levels that were as twice as much as the ones measured with the PRRSV isolate treatment (Fig. 4A, B). In addition, PRRSVchim and PRRSV isolate induced similar levels of MCP-1 chemokine in the murine macrophages (Fig. 4C). Furthermore, GM-CSF was expressed 24 h after incubation with PRRSVchim, independently of the concentration of protein used (Fig. 4D). The chemokine RANTES was overexpressed in the cells 24 h after the addition of the virus isolate, in contrast to the PRRSVchim protein treatment (Fig. 4E). Finally, both PRRSVchim and PRRSV virus induced the production of IL-10 (Fig. 4F).
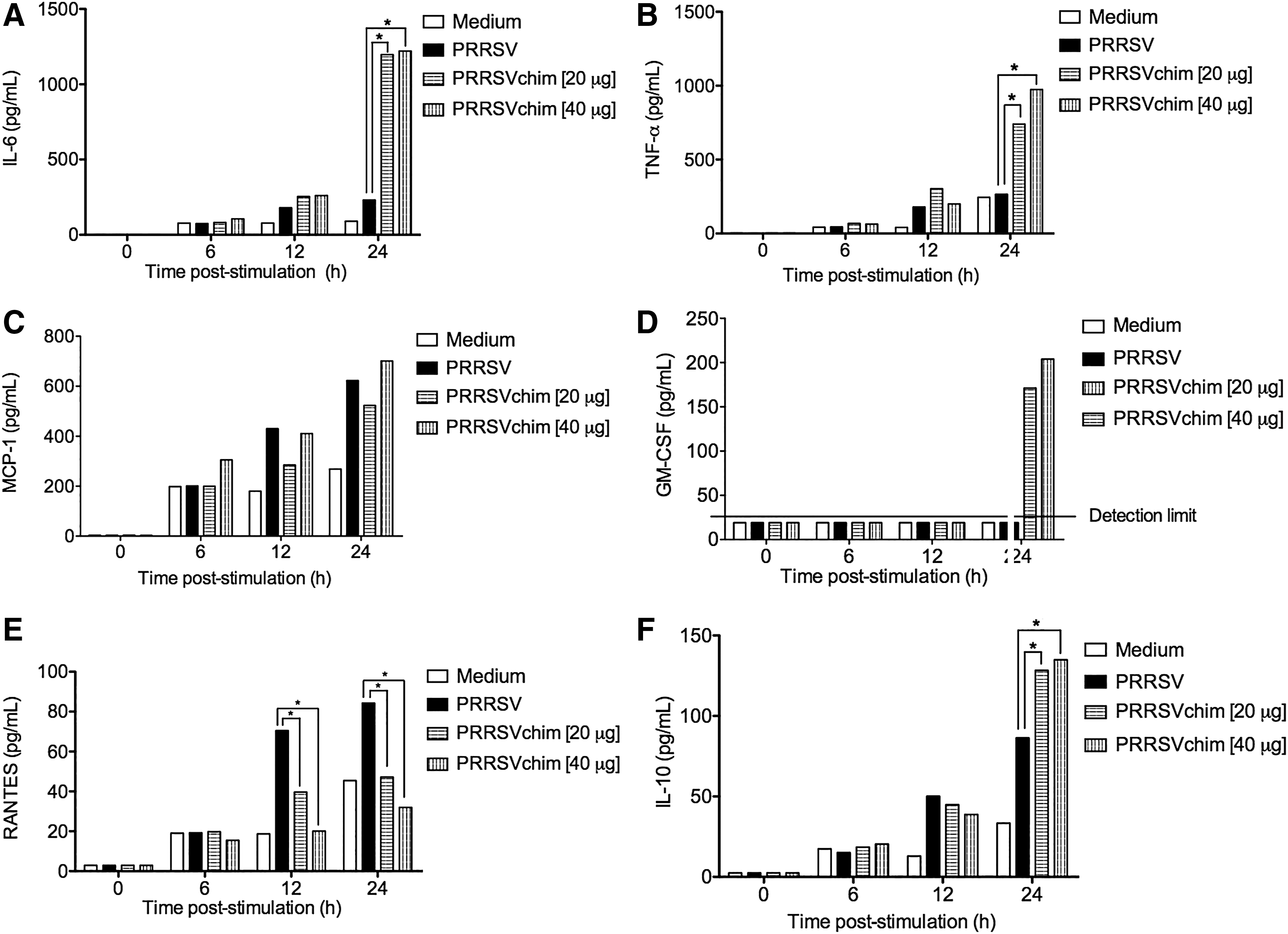
Murine macrophage cell line stimulation assay with PRRSV chimeric protein. RAW 264.7 cells were incubated with two different amounts of PRRSV chimeric protein (PRRSVchim [20 μg] and PRRSV [40 μg]), a PRRSV mexican isolate (multiplicity of infection of 0.1) or medium. Supernatants were taken at different time points for measuring cytokine levels [
PRRSV chimeric protein antigenicity in mice and piglets
In a first instance, PRRSVchim protein antigenicity was assessed in BALB/c mice by subcutaneous administration, with or without an adjuvant, in a prime-boost scheme. Sera taken 28 days after the first immunization were analyzed for PRRSVchim-specific IgG, IgG1, IgG2a, and IgG2b. When administered along with Freund's incomplete adjuvant, the protein induced IgG, IgG1, IgG2a, and IgG2b titers of 312,500, 1,562,500, 62,500, and 12,500, respectively. Additionally, the protein alone was able to induce IgG and IgG1 titers of 62,500 (Fig. 5).
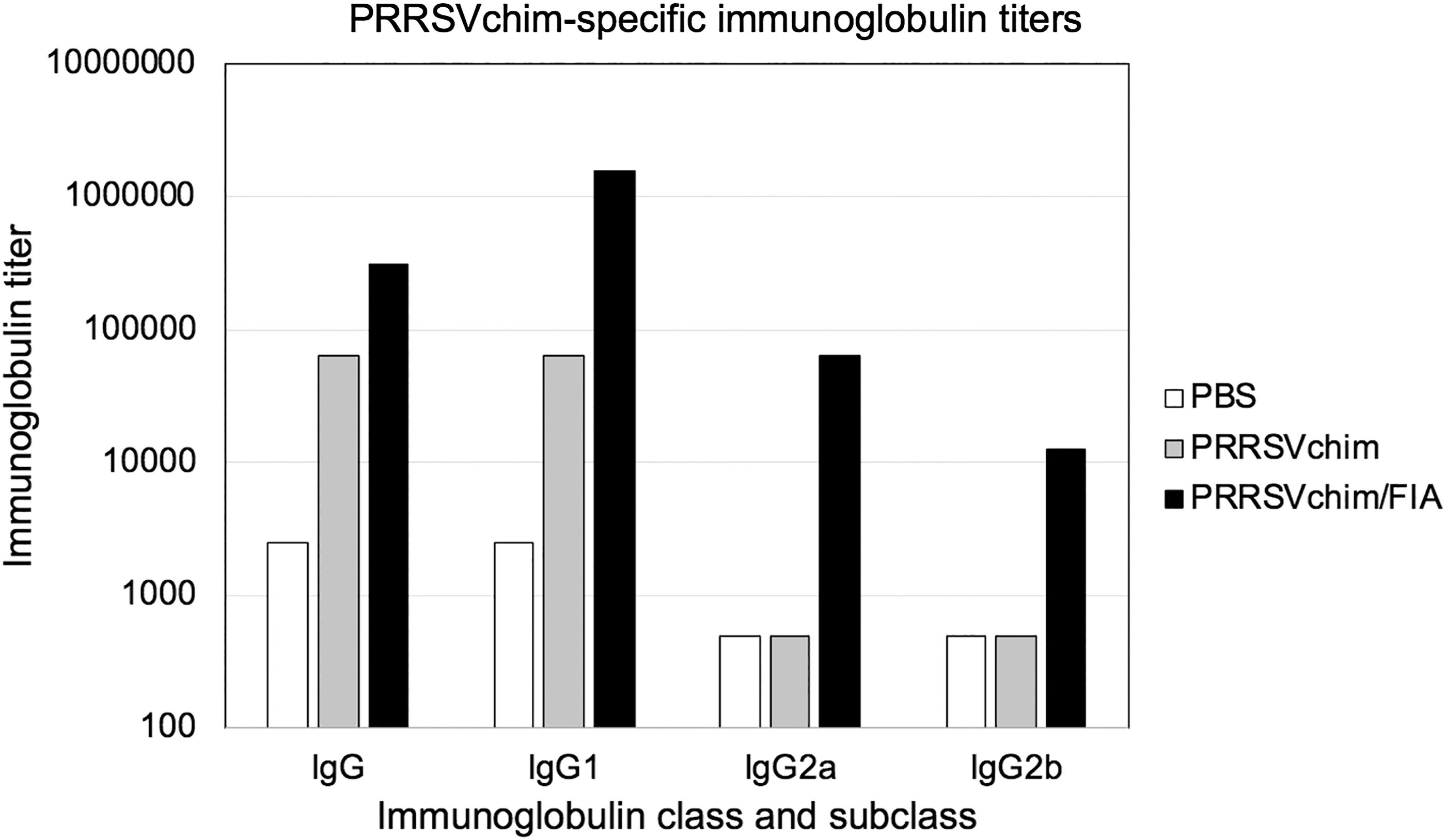
IgG response of mice immunized with PRRSV chimeric protein. IgG, IgG1, IgG2a, and IgG2b antibody titers against PRRSV chimeric protein were measured in serum samples from mice injected subcutaneously in a prime/boost regime with PBS, 10 μg of PRRSV chimeric protein (PRRSVchim), or 10 μg of PRRSV chimeric protein with Freund's incomplete adjuvant (PRRSVchim/FIA). The titers correspond to serum samples taken 28 days after priming. PBS, phosphate-buffered saline.
The antigenicity of PRRSVchim protein was evaluated in piglets. PRRSV seronegative piglets were immunized intramuscularly at days 0 and 14 with 100 μg of PRRSVchim protein alone, or adjuvanted with either a mineral oil-based adjuvant (Montanide ISA 15 A VG) or a polymeric adjuvant (polymeric gel 01). A commercial vaccine, Ingelvac PRRS MLV (Boehringer Ingelheim), was also included as positive control in the experiment. Piglet seroconversion was assessed by ELISA with either a commercial test or a PRRSVchim protein-coated microplate assay. The IgG response against PRRSVchim protein was observed 14 days after priming with the protein in combination with either adjuvant (Fig. 6A). This response was faster than that observed after immunization with the commercial vaccine, which took an additional week to reach similar IgG levels. In addition, PRRSVchim protein was able to induce an IgG response without the aid of an adjuvant, however, this response could be clearly distinguished 28 days after priming, indicating that boosting was necessary for this increase. PRRSV seroconversion using a commercial test was only observed in the group that was immunized with the commercial vaccine (Fig. 6B).
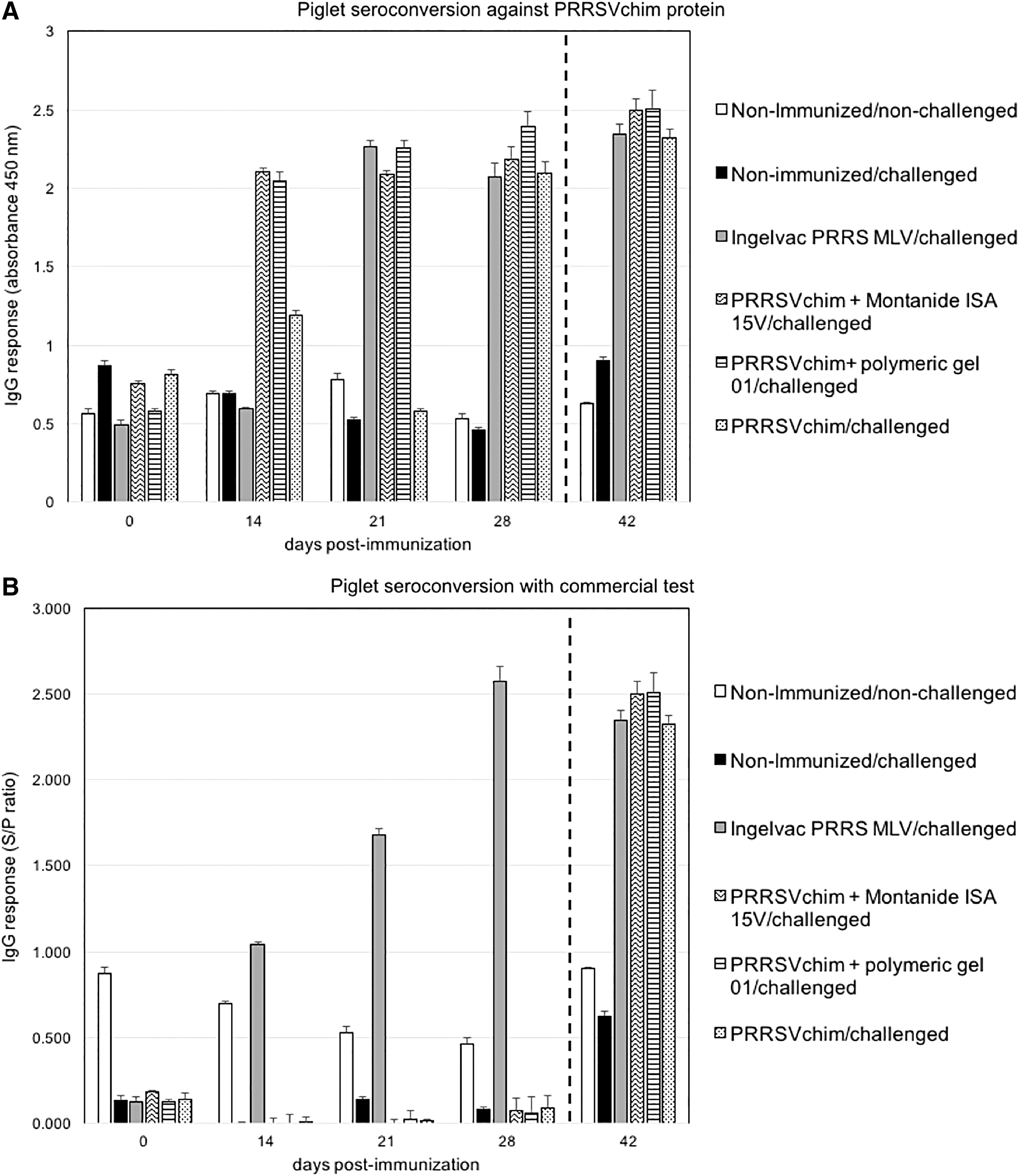
Seroconversion of piglets immunized with PRRSV chimeric protein. Seroconversion of piglet groups that received no treatment and no challenge (nonimmunized/nonchallenged), no treatment and challenge (nonimmunized/challenged), Ingelvav PRRS MLV commercial vaccine and challenge (Ingelvac PRRS MLV/challenged), PRRSVchimeric protein adjuvanted with Montanide ISA 15 A VG and challenge (PRRSVchim+Montanide ISA 15V/challenged), PRRSV chimeric protein in combination with polymeric gel 01 and challenge (PRRSVchim+polymeric gel 01/challenged) or PRRSV chimeric protein alone and challenge (PRRSVchim/challenged), was followed at different timepoints. Timepoint 42 (separated by a dashed line) corresponds to seroconversion 2 weeks after challenge with PRRSV field isolate. Seroconversion against PRRSV chimeric protein
Pig challenge with Mexican PRRSV field isolate
Pigs were challenged with a Mexican PRRSV field isolate belonging to the American genotype 29 days after the first immunization. All but one nonimmunized group, received a 1 mL intranasal and intramuscular dose of 1 × 107 TCID50 of the field isolate. Fourteen days after PRRSV inoculation, the presence of viral genomes in different samples from each individual was detected by isothermal reverse transcription-polymerase chain reaction (RT-PCR). The nonvaccinated/challenged group was 100% positive in sera, nasal swabs, and organ pools. As opposed to the nonvaccinated/vaccinated group, the group immunized with the commercial modified live vaccine resulted negative in sera and nasal swabs, whereas 60% positive in organs pool. From the treatments that included PRRSVchim protein, the one that elicited the best protection was the polymeric gel-adjuvanted formulation, with 0%, 80%, and 100% PRRSV positives in nasal swabs, organs, and sera, respectively (Fig. 7). Seroconversion after challenge was also assessed (Fig. 6A, B). The most important change was observed in the PRRSVchim-immunized groups as assessed with the commercial test, with an increase in PRRSV-specific IgG levels. No differences were detected in rectal temperature, ocular secretion, and weight gain among groups (data not shown).
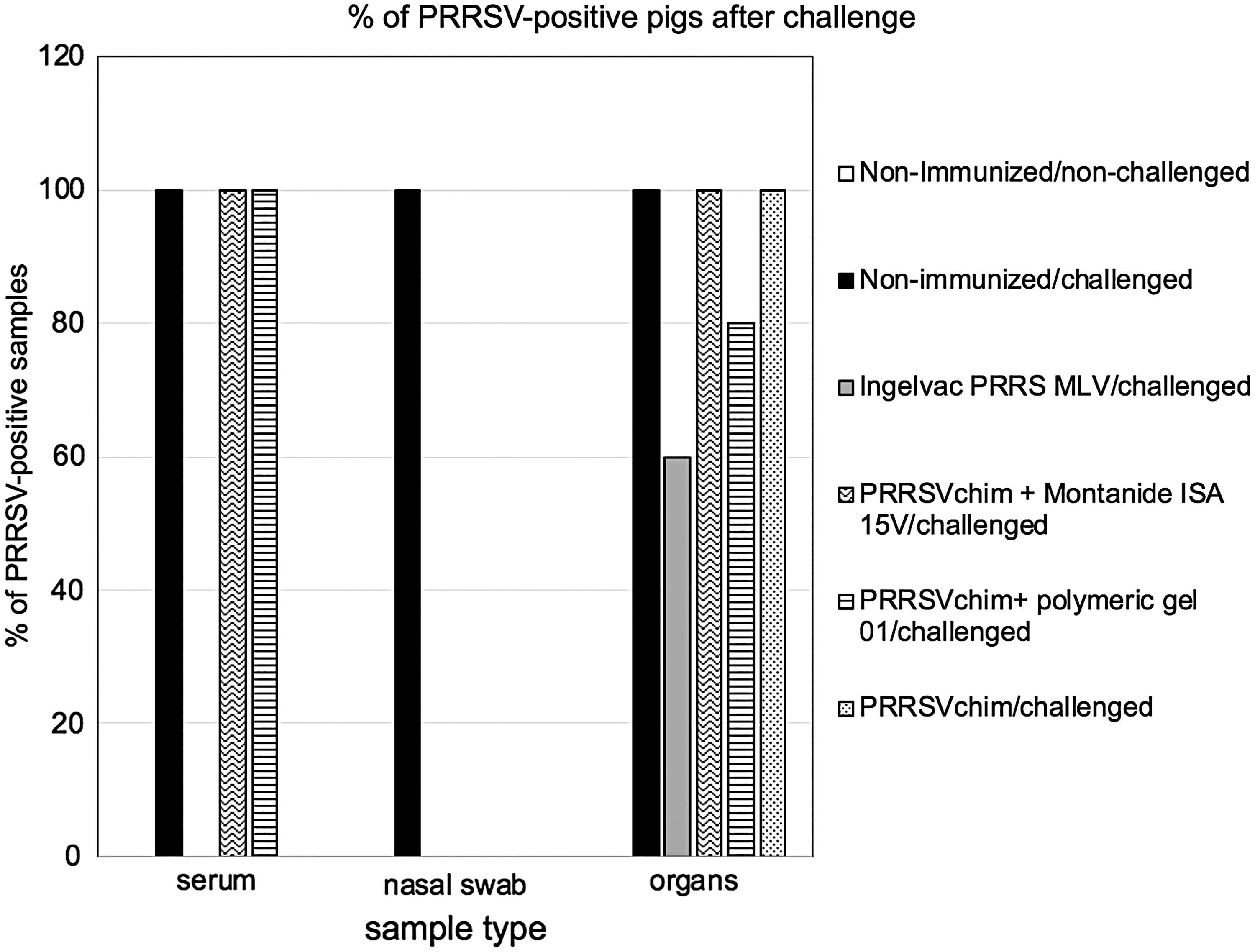
PRRSV detection in sera, nasal swabs and organs of pigs challenged with a Mexican virus isolate by isothermal RT-PCR. The presence of viral PRRSV genomes in sera, nasal swabs and organs (lungs, tonsils, lymph node and spleen) of individuals from all groups was assessed by an isothermal RT-PCR kit that detects North American PRRSV genotypes 2 weeks after inoculation with a Mexican virus isolate. The groups are the same as in Figure 6. Data are expressed as percentage of positive samples in each group. RT-PCR, reverse transcription polymerase chain reaction.
Discussion
At present, the commercial vaccines against PRRSV are mainly based on modified/live or inactivated viral particles. The antigenic complexity of PRRSV due to the several structural proteins it contains and the arising of new virus variants, affects the protective efficacy of these vaccines, pushing forward the development of upgraded vaccines that incorporate the new variants. For example, a high genetic diversity in the PRRSV strains, all belonged to the type 2, located in semitechnified and technified swine farms in seven of the eight swine productive regions of Mexico was found, mostly in GP5 and N proteins (26). Also, some PRRSV strains are difficult to grow in mammalian cell lines that are routinely employed for PRRSV vaccine production. In line with this, the main goal of this work was to develop a flexible platform that allows for the fast development of subunit vaccines against PRRSV based on short PRRSV sequences from GP3, GP4, GP5, and M proteins, reducing antigenic complexity. This way, a PRRSV chimeric protein, named PRRSVchim, was designed and it included two epitopes from GP3, one epitope from GP4, as well as GP5 and M ectodomains, each separated by a four alanine spacer sequence. The short sequences that comprise PRRSVchim were selected according to previous reports that demonstrate the importance of these in the onset of neutralizing antibody production against PRRSV (9,33,48,54). The protein was thought to be expressed in E. coli, because new versions that include the most recent amino acid changes in PRRSV variants can be generated in a few months using this expression model.
PRRSVchim protein was successfully expressed in the soluble fraction only when fused to thioredoxin. This result highlights the importance of this fusion partner for achieving expression of the chimeric protein. This fusion partner has been extensively utilized in previous works for increasing solubility of a variety of proteins of interest and served in this case for providing stability to the chimeric protein, making its accumulation possible in E. coli cultures. For instance, thioredoxin 2 (trx2p) helped in the stable expression of a viral antigen in Saccharomyces cerevisiae (30). Expressing PRRSVchim protein in the soluble fraction would aid in avoiding some steps during the downstream process, like protein solubilization with high concentrations of chaotropic agents and further protein refolding. Previous works closely related to the present study, where no fusion partner was used, reported that GP5 without the original secretion peptide and the transmembrane sequence and a chimeric protein comprising GP5 and M ectodomains accumulated in E. coli inclusion bodies (16,34). Unexpectedly, the fusion of glutathione S-transferase, another fusion partner that is commonly used, to truncated GP5 did not avoid its accumulation in inclusion bodies, but fusing flagellin B from Salmonella typhimurium to the same protein contributed to its soluble expression (40). PRRSVchim protein was purified by two sequential rounds of IMAC and its recognition by IgG from PRRSV-positive serum samples was assessed. All PRRSV-positive samples recognized PRRSVchim protein. This result suggests that PRRSVchim protein keeps its antigenic properties and also that it could be useful as reagent in an immunodiagnostic test for detecting antibodies against PRRSV in serum. An additional experiment that is able to differentiate the individual interaction of serum IgG from PRRSV-infected animals with each of the sequences that comprise the chimeric protein would be desirable.
To have a quick glance of the innate immune response that PRRSVchim protein alone can induce, the nonpermissive murine macrophage cell line Raw 264.7 was incubated with the protein and cytokine profile was measured in culture supernatants. Nonetheless, it is important to mention that the results derived from this in vitro assay would hardly reflect what happens in an animal immunization experiment. As reference, the murine macrophage cell line was also incubated with a PRRSV virus Mexican isolate. It has been documented that PRRSV shows tropism to (PAMs) (7,57), and generally permissive macrophages are used to verify virus infection and the response of these infected macrophages. Since the PRRSVchim protein does not have the ability to infect macrophages by itself, we considered that using a permissive cell line would not be appropriate for assessing its activation after being in contact with a protein that only displays immunogenic epitopes of the virus. Therefore, a murine macrophage cell line (nonpermissive) was more suitable for this experiment. Previous in vitro and in vivo analyses have demonstrated that when PRRSV infects PAMs, proinflammatory cytokines, such as IL-1β, TNF-α, IL-6, IL-8, and IL-12, are overexpressed (4,37,38,56). In this study, the analysis of cytokine profile production showed that PRRSVchim stimulated a two-fold increase of M1 cytokines (IL-6 and TNF-α) in the murine macrophage cell line in comparison to the viral treatment. It has been reported previously that induction of these cytokines in permissive cell lines is strain dependent (2,14). Interestingly, a higher level of M2 cytokine IL-10 was produced for the virus isolate and the PRRSVchim (Fig. 4F). An extensive analysis using a total of 39 PRRSV strains tested in PAM, peripheral blood mononuclear cells, bone marrow-derived dendritic cells, and peripheral blood SwC3+ cells, showed that TNF-α and IL-10 induction is strain dependent (12). The PRRSV strain used in this experiment was able to induce both cytokines. RANTES has been considered as a pro inflammatory chemokine (39,50) and PRRSV infection activates RANTES messenger RNA (mRNA) at a dose-dependent ratio with a maximum expression at 48 h in permissive MARC-145 cells (50). There is some evidence that indicates that RANTES contributes to the infection of PAMs and lymphoid tissues (13,53). In this study, RANTES was overexpressed in Raw 246.7 cells 24 h after their incubation with the PRRSV isolate, in contrast to the PRRSVchim protein treatment, where RANTES induction was low (Fig. 4E). This may represent a substantial advantage of using the PRRSVchim protein instead of a live attenuated vaccine, but more detailed studies should be conducted to clarify this discovery in the perspective of a potential synthetic regulator. GM-CSF was expressed 24 h after incubation with PRRSVchim, independently of the concentration of chimeric protein (Fig. 4D). The function of GM-CSF is related to the maturation of macrophages; this was demonstrated when a recombinant PRRSV expressing GM-CSF was used to induce a DC cellular response (55). Finally, the stimulated macrophages produce MCP-1 in response to both PRRSV isolate and PRRSVchim protein (Fig. 4C). This chemokine was described for granulomatous lesions in piglet lungs (21).
The last part of this study consisted in the evaluation of the immune response induced by PRRSVchim protein in mice and piglets. PRRSVchim administration to mice induced high IgG titers, even in the absence of an adjuvant. This could be a consequence of the intrinsic property of the chimeric protein to form aggregates, which are readily detected by the immune system (25). In the piglet immunization experiment, PRRSVchim protein elicited a fast IgG response in combination with two different adjuvants. It seems that an important feature that a good PRRSV vaccine candidate must possess is its ability to trigger a rapid immune protection, derived from both humoral and cellular immunity (24). As reviewed by Renukaradhya et al. (35), there are several experimental subunit vaccines, most of them based on GP5 that have demonstrated their efficacy in the induction of neutralizing antibodies against PRRSV after their administration to pigs. Hence, subunit vaccines carrying one or several antigens or protective epitopes from PRRSV are becoming an alternative strategy to enhance immunoprophylaxis against this virus on a global scale. A lack of seroconversion in the PRRSVchim-immunized animals was observed when a commercial test was employed. A possible explanation for this finding is that the commercial test might be coated with an antigen other than PRRSV GP3, GP4, GP5, or M proteins. It should be noted that the main target of the widely used serologic PRRSV test from IDEXX Labs is N protein. So, it would not be surprising that the commercial test employed in this study to assess seroconversion against PRRSV detects mainly N protein also. Finally, the challenge of the pigs against a PRRSV isolate that belongs to the American prototype, as confirmed by ORF5 sequencing, revealed that the RespPRRS MLV vaccine provided the highest protection efficacy, in terms of PRRSV positivity, followed by PRRSVchim protein adjuvanted with polymeric gel. It is noteworthy to mention that RespPRRS MLV vaccine and the three PRRSVchim protein formulations were able to reduce viral presence in nasal swabs to nondetectable levels with the test used in this study, which could reduce in turn viral shedding in field swine exploitations. Taken together, the results presented in this study show that PRRSVchim protein could be evaluated as reagent for immunodiagnostic tests. On the other hand, before considering this protein for evaluation as immunogen in field trials for the control of PRRSV, it is essential to address two questions: what are the neutralization titers that are elicited after PRRSVchim protein administration and what are the viral loads in individual organs, nasal swabs, and sera after PRRSV field isolate challenge? A virus neutralization experiment employing PRRSV-permissive cells and PRRSV field isolates will give a better idea of the antibody protective immunity for preventing viral entry and, subsequently, viral infection. Finally, assessing viral titers in individual organs, nasal swabs, and sera by means of quantitative RT-PCR or TCID50 after challenge, will contribute to define the efficacy of the protein in preventing virus multiplication in immunized pigs.
Authors' Contributions
D.E.-Q. helped in designing the experiments, analyzing and discussing results, and drafted the article. L.Z.-C. carried out protein expression and purification. J.A.U.-F. helped in carrying out protein expression and purification and assessed the interaction of PRRSVchim protein with sera from PRRSV-infected pigs. J.G.-B. designed and carried out the murine macrophage stimulation assay with PRRSVchim protein. T.A.C.-V. helped in designing the murine macrophage stimulation assay with PRRSVchim protein and article drafting. C.A.M.-C. performed the antigenicity of PRRSVchim protein in mice. M.E.T.-O. provided the PRRSV isolate used in the murine macrophage stimulation assay with PRRSVchim and helped in designing the experiment. G.R.H., M.A.H.-L. and M.C.M.-G. designed and performed the piglet immunization/challenge experiment and the antigenicity of PRRSVchim protein. A.G.O. conceived the project, helped in experiment design and article drafting, and supervised the whole study. All authors read and approved the final article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Fondo Sectorial de Innovación Secretaría de Economía-CONACYT (FINNOVA) project 238667 and by Programa de Estímulos a la Innovación (CONACYT-PEI) project 244089.