
Abstract
Dengue virus (
Introduction
Dengue virus (DENV) has become the most important pathological virus transmitted by vector mosquitoes (76). Dengue outbreaks affect populations, health systems, and economies of many tropical countries around the world. Dengue incidence at a global scale has increased 30 times in the last 40 years (11). Emergence, re-emergence, and propagation of the four
The presence of the mosquito vector Aedes aegypti and favorable conditions for breeding its populations favor the DENV dispersion. A recent estimation showed that 390 million infections occur annually and that up to 50% of the world population lives in areas with a high risk of DENV infection (11).
Further predictions based on potential climate change and modification of mosquito population range indicate that up to a billion people could be at risk within the next century (39). Propagation of the vector mosquito Ae. aegypti within unregulated newly formed urbanizations have facilitated the re-emergence of Dengue outbreaks throughout tropical and subtropical regions (11).
Multiple efforts are being carried out to generate a Dengue vaccine for the protection of populations at risk of infection. Unfortunately, none of these efforts has produced the required success for a vaccine that would be applicable on a global scale (33). Recently, the most advanced vaccine, Dengvaxia, was authorized and applied to some populations (50). However, the application of this vaccine requires specific epidemiological context regarding DENV exposure prevalence within the target population. The potential effects of this vaccine on naive individuals have led to a halt on its application recommended by the World Health Organization (WHO) (77). Also, the WHO has recommended that in the case of a successful vaccine emerging, vector control strategies should complement the vaccination programs.
DENV transmission to human populations and the consequent generation of outbreaks of DENV-associated pathologies require the participation of a competent vector. Therefore, mosquito molecules and pathways that contribute to vector competence have the potential to impact DENV transmission. Identifying and understanding the components of this complex mosquito character should allow the design and maintenance of strategies directed to improve entomological management to block DENV transmission. Therefore, gaining knowledge of vector-virus interactions remains relevant to generate transmission-blocking strategies.
Although many studies are analyzing various aspects of vector biology, in this study, we concentrate mainly on scientific reports where
Vector Competence
Ae. ageypti vector competence for DENV transmission
Vector competence consists of the intrinsic characteristics of Ae. aegypti populations that allow DENV to infect, replicate, and disseminate through the tissues of the mosquito that can then transmit the virus to human hosts. Ae. aegypti mosquitoes acquire DENV through blood-feeding from infected individuals.
Once the virus has replicated within midgut cells and escapes the midgut barrier, it can reach other tissues and cells, including the hemocytes and the fat body. When DENV reaches the hemocoel, viruses can then infect the salivary gland, replicate within its cells, and reach the salivary gland lumen, where viruses can then be inoculated into a persońs bloodstream when the mosquito bites for feeding (Fig. 1). This replication cycle implies that the virus has to infect and replicate within a diversity of mosquito cell types, potentially affecting multiple molecules and pathways.

Vector Competence and Innate Immunity
DENV infection induces an immune response in Ae. aegypti
Since the mosquito midgut is the first tissue that becomes infected (58), it has been an important focus to study the vector response to DENV infection in search of the molecules and pathways involved in this process. Therefore, multiple efforts and a variety of approaches have been directed to characterize the DENV-infected mosquito midgut at a molecular level. The public release of the first Ae. aegypti genomic sequence draft (42) facilitated studies of vector-virus interactions. The initial microarray-based transcriptomic analyses, coupled with functional assays, revealed the participation of the Toll innate immunity pathway in mosquito antiviral response to DENV infection (79).
DENV infection downregulates Ae. aegypti immune response
The vector-DENV interaction does not only involve the activation of the mosquito innate immune response. The analysis of in vivo infection of Ae. aegypti with DENV has shown that at later stages of infection, the majority of affected genes are downregulated by the viral infection in the midgut, carcass, and salivary gland. DENV infection of Ae. aegypti induces downregulation of multiple transcripts in various strains, including the highly susceptible Rockefeller strain infected with DENV2 NGC (79), the salivary gland of Liverpool (LVP) strain infected with DENV2 Thailand 16681 (35), and the susceptible, recently colonized, Chetumal (CTM) strain infected with DENV2-Jam1409 (15).
An important observation of these studies is that the abundance of immunity transcripts in DENV-infected mosquitoes is lower than that in noninfected mosquitoes, showing that DENV infection also induces downregulation of mosquito immune response. Consistent with those findings, the successful infection of Ae. aegypti-derived cell line Aag2 with DENV also involves the downregulation of multiple genes, in particular, innate immunity genes (62). Therefore, although the mosquito generates a potent immune response upon DENV infection, the viral replication cycle induces downregulation of this response in various mosquito tissues. The involvement of mosquito innate immunity in this complex dynamic interaction suggests that it is a critical constituent of vector competence for DENV transmission.
The basal level of innate immunity correlates with the degree of mosquito susceptibility to DENV infection
It has been observed that mosquito populations may differ in their degree of susceptibility to DENV infection (16,63,66). Variations in the midgut expression levels of multiple transcripts among Ae. aegypti strains are readily detected in noninfected mosquitoes. For instance, there is a significant variation of transcript abundances between three Ae. aegypti strains: CTM, D-Puerto Rico (Rex-D), and LVP (16). These differences in basal gene expression profiles could be related to the differential susceptibilities of these strains to DENV infection.
Comparison of transcriptional midgut profiles among other susceptible refractory field and laboratory strains has shown that the pattern of basal expression groups them into refractory strains on one side and susceptible strains on the other. More importantly, it shows that heightened basal expression of innate immunity genes correlates with a higher level of refractoriness (63). This correlation of the basal level of expression of immunity genes with the mosquito susceptibility to DENV infection further supports that the mosquito innate immune response is a component of vector competence.
As a consequence, the role of each of the three canonical mosquito innate immunity pathways, Toll, immune-deficient (IMD), and JAK-STAT, in limiting infection by DENV has been the focus of further studies. Such studies have contributed to the identification of critical regulatory and effector components of the three innate immunity pathways in Ae. aegypti. In the following section, we briefly review the current knowledge about each of these three pathways (Fig. 2) in Ae. aegypti.
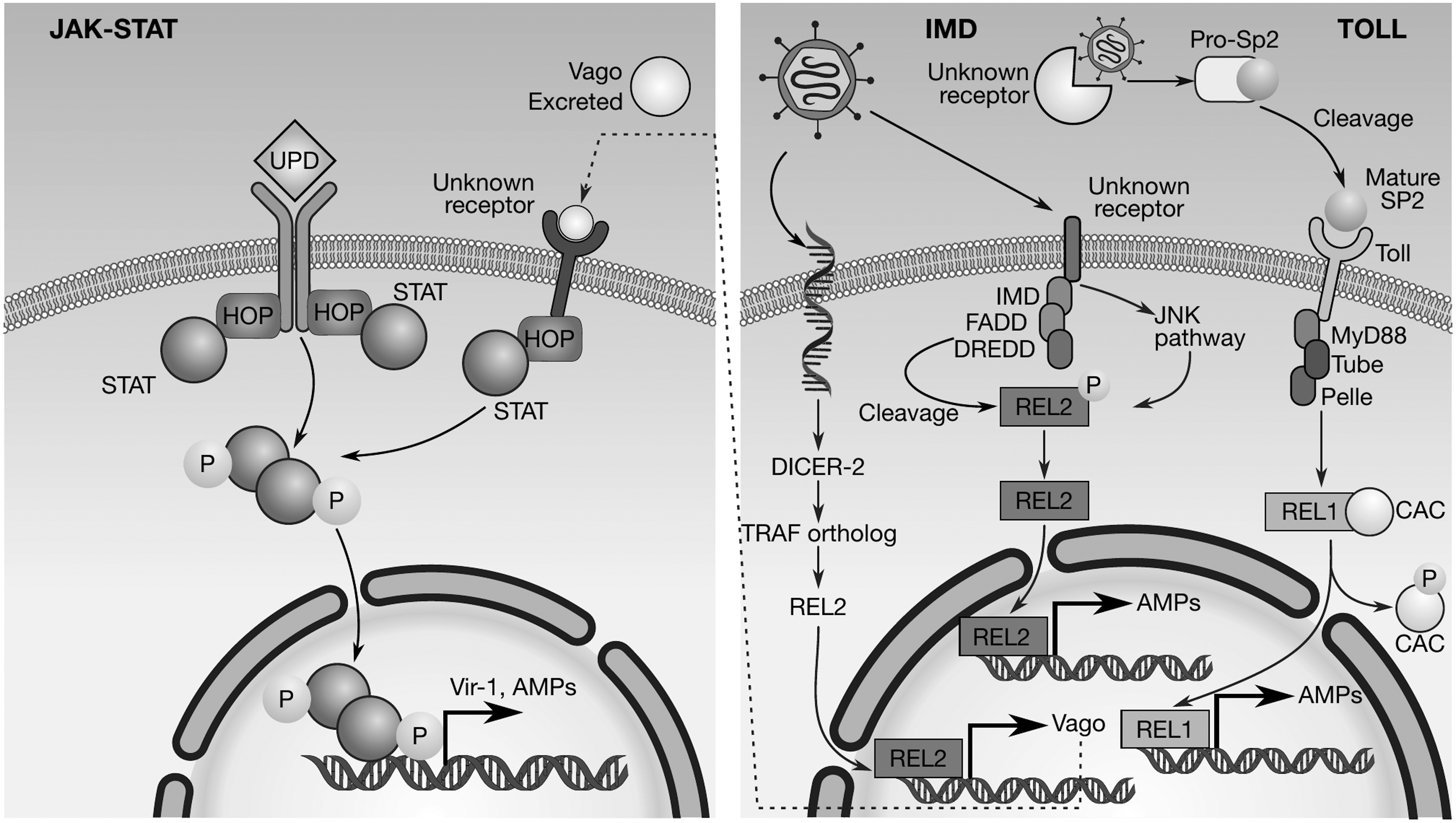
Innate immunity pathways in mosquitoes. Infection with
Innate Immunity Pathways
The toll pathway
The Toll pathway participates in Ae. aegypti anti-dengue defense. Infection with DENV-2 induces upregulation of genes belonging to the Toll pathway, suggesting that it plays a crucial role in regulating the mosquito's ability to limit its infection by DENV (79). These Toll pathway genes include Toll, Rel1A, and Spätzle (Spz); consistent with the activation of Toll, its negative regulator, Cactus, becomes downregulated (79). Subsequent studies of additional mosquito field strains and DENV serotypes (DENV-4) at an earlier stage of infection (3-day postinfectious blood meal) have confirmed the conserved role of the Toll pathway in mosquito anti-dengue defense (51). In this case, an impaired Toll pathway, achieved by attenuation of MyD88 gene expression, also increases DENV viral particles in the mosquito midgut, independent of mosquito strain.
Interestingly the midgut and carcass have shown differential responses to viral infection. For instance, at 10-day postinfection, the carcass shows that more transcripts are being upregulated or downregulated, as well as more significant fold changes in transcript levels, compared to the midgut (51). Nevertheless, the majority of transcripts that change in abundance correspond to immunity genes in both tissues. Functional analyses, by reverse genetics, have confirmed the participation of the Toll pathway in mosquito defense against DENV-2 infection (79).
Whole-body gene expression analyses have confirmed activation of the Toll pathway by DENV infection and provided evidence of Toll pathway differential response to DENV serotypes (65). In this case, the DENV infection induces increased expression of Spz and Rel1A, and transient upregulation of Cactus, which becomes downregulated after 7 days postinfection (65). Analysis of some field Ae. aegypti populations showed that, although they differ in the basal level of Rel1 expression, DENV infection does not induce differential expression of this gene (19). Such analysis was not carried out at earlier stages, where others have found DENV induces downregulation of the Toll pathway (20,51,62).
Since the salivary gland is directly involved in the process of virus transmission, efforts have been made to characterize virus-mosquito interactions in this organ. Blood feeding itself induces upregulation of a small number of immunity-related genes, including Defensin 1A (69). However, the salivary gland has a distinct pattern of the innate immune response to DENV infection, when compared to other mosquito organs.
An important feature of the salivary gland response is the downregulation of the Toll pathway, suggesting that DENV infection alters mosquito innate immune response (15,64). Some variants on the 3′UTR of DENV-2 produce subgenomic flaviviral RNA that inhibit the Toll pathway in mosquito salivary glands, enhancing the production of infectious saliva (49). This finding directly illustrates and adds support to the notion of the mosquito Toll pathway relevance in Dengue transmission.
The Toll pathway also participates in the DENV replication interference induced by the bacterial endosymbiont Wolbachia; in this case, there is evidence that the bacteria induce oxidative stress and this, in turn, activates the Toll pathway (46). However, others found that the blocking of DENV replication, induced by the Wolbachia endosymbiont, does not require the Toll pathway (55). This finding would suggest that there could be additional, redundant mechanisms that contribute to the inhibition of DENV replication. The evidence summarized in this section supports the notion that the Toll pathway plays a critical role in mosquito vector competence for DENV transmission.
The IMD pathway
The IMD pathway in mosquitoes is similar to the NF-κB signal transduction pathway of vertebrates. This pathway is known for its antibacterial and antiparasite activity in mosquitoes (80); more recently, it has also been shown to be involved in the antifungal response in Ae. aegypti, (52).
The IMD pathway in mosquitoes is activated when a microbe binds to a transmembrane receptor (Fig. 2), triggering the recruitment of some adapter proteins. The factors fas-associated dead domain (21) and the death related ced-3/Nedd2-like protein (DREDD) are recruited to then cut the phosphorylated C-terminal domain of Rel2 (Drosophila Relish ortholog in the mosquito). Activation of the JNK pathway, an additional branch of the IMD pathway, leads to Rel2 phosphorylation (26,61). Transcription factor Rel2 is subsequently translocated to the nucleus to activate transcription of effector genes.
Activation of the IMD pathway leads to transcriptional activation of effector genes for the production of antimicrobial peptides (AMPs) that counteract microbial invaders. In Ae. aegypti, 17 AMPs have been identified; they belong to five distinct families (72), and it has been shown that they are mainly regulated by the IMD pathway (81). The IMD pathway plays an important role in Ae. aegypti response against DENV; activation of Ae. aegypti IMD pathway by silencing its negative regulator Caspar does not modify DENV infection (79). However, in a DENV refractory strain where the IMD pathway was obliterated by gene silencing, the viral titers were increased in the midgut, thus suggesting that this pathway may participate in mosquito anti-dengue defense (63).
In the Ae. Aegypti-derived cell line Aag2, DENV infection-induced downregulation of immune response allows improved bacterial growth in co-culture, suggesting that the IMD pathway is suppressed by the virus and therefore this pathway could be implicated in anti-dengue defense (62). Also, the expression of a cecropin-type AMP gene (AAEL000598), regulated by the IMD pathway, becomes increased in the salivary gland by DENV infection; the cecropin-type peptide encoded by this gene displays antiviral activity against DENV (35).
Further evidence of IMD pathway participation in anti-dengue defense has been provided by blocking this pathway in DENV-refractory mosquito strains, which leads to an increase in viral replication (63). Some C-type lectin domain protein genes (2) are regulated by the IMD pathway and change their pattern of expression upon DENV infection in Ae. aegypti (5,15,20,64). The specific role of these proteins is complex and not yet fully understood (2), but their change in expression levels upon viral infection also implicates the IMD pathway in mosquito interactions with DENV.
The JAK-STAT pathway
Ae. aegypti infection with DENV-2 induces upregulation of genes belonging to the JAK-STAT immunity pathway (79). Functional studies (silencing of PIAS, a negative regulator of the JAK-STAT pathway) have shown that viral infection is reduced between twofold and fivefold at different times postinfection. Therefore, the expression of the mosquito JAK-STAT pathway genes can limit DENV infection. The function of this immunity pathway in the defense against viral infections is then conserved in mammals, Drosophila, and mosquitoes (67). It has been observed that genomic and phenotypic plasticity have led to the formation of distinct Ae. aegypti populations regarding their susceptibility to DENV infection.
The analysis of Moyo-S and Moyo-R, two closely related strains (both derived from the Moyo-in-Dry strain (44)) with contrasting degrees of susceptibility, has allowed the discovery of gene networks potentially involved in vector competence determination (10). In this study, the early response to viral infection (DENV-2 JAM1409) was characterized after an infected blood meal. Genes from the JAK-STAT pathway become upregulated in both susceptible and refractory strains (10), implying that this pathway constitutes a core element of the vector immune response to viral infection. Therefore, this suggests that the JAK-STAT response is independent of vector competence status.
To better understand the mechanistic dynamics of mosquito response to
RNA Pathways
One of the most effective antiviral responses in mosquitoes is the interfering RNA pathway that controls infection by the degradation of viral RNA (Fig. 3). A critical event in the interfering RNA pathway is the production of small RNA molecules from long double-stranded RNAs (dsRNA) (24). Three main types of small RNA molecules participate in regulatory and defense mechanisms: small interfering RNAs (siRNAs), micro RNAs (miRNAs), and PIWI-interacting RNAs (pi(RNAs).
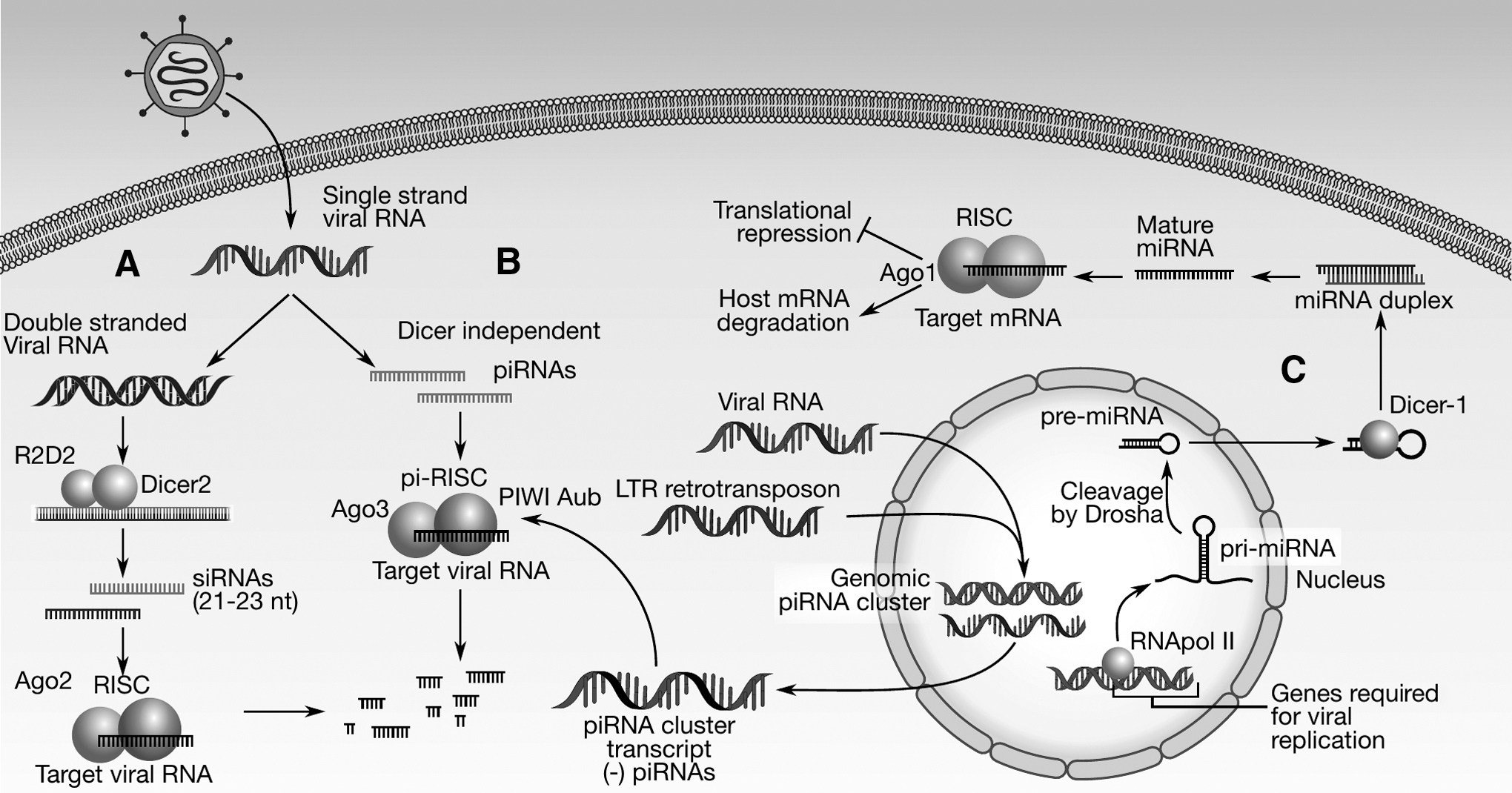
Antiviral RNA pathways in Aedes aegypti.
DENV-infected Ae. aegypti mosquitoes generate DENV-specific siRNAs and P-element-induced wimpy testis (PIWI)-associated RNAs (piRNAs) (27). The disruption of the RNAi pathway in DENV-infected mosquitoes by gene silencing induces an increase in viral replication, suggesting that this pathway is critical to control viral replication (59). siRNAs extracted from mosquito tissues have been found to match viral genomes. The exogenous siRNA pathway is mainly related to the antiviral RNAi. DENV-2 NS4B is an RNAi suppressor in mosquitoes (30).
The RNAi pathway is the most effective mechanism of antiviral defense in mosquitoes and other arthropod vectors. In this pathway, dsRNAs (generated by secondary structures of RNA virus genome or other dsRNA molecules generated or introduced by the virus) are detected and identified as a pathogen-associated molecular pattern by Dicer-2. The subsequent formation of a complex with pre-RNA-induced silencing complex (RISC) leads to the fragmentation of the target RNA producing 21 base fragments, which will repeat the cycle and amplify the effect of RNA degradation, thus interfering with viral RNA functions.
RNA molecules are involved in Ae. aegypti response to DENV infection; the RNA interference (RNAi) defense mechanism has been shown to become activated in Ae. aegypti against viral infection (25). The RNAi response is an important defense mechanism triggered by the presence of viral dsRNAs produced by the replication process (31,74). These long dsRNAs can form secondary structures at the UTRs. Strain-specific sequences within these regions can modify such secondary structures; a fragment of 400 nucleotides of DENV UTR is capable of downregulating the RNAi response (60).
Each DENV serotype is constituted by multiple genotypes, differing in the nucleotide sequence by 6%. These genotypes include multiple lineages that have been denominated strains, clades, lineages, or groups (57,73). Strains from a single serotype can participate in competitive strain displacement (37,38). An analysis of strains involved in displacement showed that a DENV-3-displaced strain is less effective in suppressing the RNAi response than a displacing strain of the same serotype (4). This mechanism shows how the interaction of dengue-specific strains with the mosquito RNAi response has a direct impact on viral transmission.
The RNAi pathway
The RNAi pathway participates in anti-dengue defense. This pathway constitutes the principal antiviral response in the mosquito (13). In this pathway, the RNAse enzyme Dicer-2 (Dcr) associates with the R2D2 protein (56). The RNase activity cuts dsRNA molecules produced by the viral RNA into fragments of 21–23 base pairs, the siRNAs. These siRNAs bind to the RISC that degrades one of the strands of the siRNA. The remaining single strand becomes a guide to detecting RNA molecules that contain a sequence complementary to the guide RNA. Molecules identified in this way become substrates for degradation by the endonuclease Argonaute-2 (ago2) (24).
In mosquitoes, the siRNA pathway participates in the control of DENV infection. DENV-infected Ae. aegypti mosquitoes have virus-specific siRNA (27). The expression profile of these virus-specific siRNAs changes throughout infection, affecting cellular processes such as mitochondrial function (27). In mosquito cell lines, the expression of dsRNA sequences derived from the DENV-2 genome activates the RNAi pathway and leads to resistance to DENV-2 infection (1). Also, in Ae. aegypti transgenic lines expressing dsRNA DENV sequences, virus-specific siRNA can be detected, which are produced by activation of the RNAi pathway (70).
The midgut specific expression of a dsRNA derived from the coding region of DENV-2 premembrane protein in a transgenic Ae. aegypti transgenic line diminishes its ability to support DENV replication (25). Gene silencing of the siRNA pathway critical components leads to an increase in viral replication and a reduction of the extrinsic incubation period (EIP) (59). Therefore, the efficiency with which the mosquito siRNA pathway counters DENV infection is a determinant of vector competence since it can modify the length of the EIP affecting DENV transmission.
The miRNA pathway
The miRNA is a class of noncoding RNA molecules that participate in the posttranscriptional regulation of gene expression (9,32). These RNA molecules play a critical role in regulating the expression of endogenous genes and the cell response to viral infections. In Ae. aegypti, expression of miRNAs involved in the regulation of immunity genes can be induced in the midgut by a blood meal (28).
DENV infection induces changes in the expression profile of miRNA molecules in Ae. aegypti (17). In this study at 9 days postinfection, 23 miRNAs change their expression profile. Each of these miRNAs has a target sequence in at least one of a group of 464 coding genes that include factors involved in transport, transcriptional regulation, mitochondrial function, chromatin modification, and signal transduction, which may support viral replication and dissemination to mosquito tissues. For instance, the aae-miR-375 (28,34) molecule can interact with target sequences on two immunity genes, Cactus and Rel1.
Mimic molecules of aae-miR-375 induce increased expression of Cactus in Ae. Aegypti-derived Aag2 cells; this negative regulator inhibits expression of the NF-kappa B transcription factor, and then the DENV replication increases in these cells (28,34). Similarly, in Aedes albopictus, and its derived cell line C6/36, DENV infection also induces differential expression of miRNAs (7,68). In the midgut, there is a pronounced differential expression of miRNAs compared with noninfected mosquitoes, especially at 7 days postinfection (68). The aae-miR-375 can also increase DENV-2 infection in the C6/36 cell line (68). Therefore, the changes induced by DENV infection on the miRNA expression profile can contribute to the mosquito vector competence for DENV transmission.
The piRNA pathway
Ae. aegypti genome has eight PIWI protein family coding genes. PIWI family genes are upregulated by blood-feeding in Ae. aegypti (14,16,22). There is evidence that piRNAs are involved in mosquito antiviral response. In the last decade, multiple research groups have reported the production of piRNAs upon viral infection in mosquitoes. Profiling noncoding RNAs by deep sequencing of DENV-2-infected Ae. aegypti uncovered the participation of the PIWI pathway in mosquito antiviral defense (27). When Ae. aegypti, Ae. albopictus, and cell lines derived from each of them become infected with viruses, infected cells generate piRNAs derived from such viruses (41). DENV infection can elicit the formation of dengue-2-derived piRNAs in mosquito midgut and carcass. However, the midgut was found to be deficient in virus-induced host endogenous piRNAs (71).
Cells can produce piRNAs from endogenous viral elements that derive from the insertion of sequences from nonretroviral viruses into the mosquito genome (40). Recently, the presence of endogenous viral sequences has been extensively investigated in the two most important DENV Aedes genus vectors and cell lines derived from their tissues.
Analysis of the genome of mosquitoes has revealed that Ae. aegypti genome contains more than 10-fold viral sequence integration compared to Anopheles and other Culex species (45). Multiple flavivirus genomic sequences have been found residing within the genome of Aag2 Ae. Aegypti-derived cells. The Ae. albopictus genome contains DENV derived endogenous sequences, but neither Ae. aegypti nor its derived Aag2 cell line genomes contain such kind of sequences. The integrated virus-derived sequences are located within piRNA clusters and can produce piRNAs. This mechanism constitutes an Ae. aegypti adaptative antiviral immunity system that relays on the integration of virus coding sequences into the mosquito genome (75).
The interferon-like antiviral response
The Vago cytokine-like factor coordinates an interferon-like immune response mechanism in Culex mosquitoes and participates in the antiviral response. Since Ae. aegypti genome contains a gene (AAEL000200) coding for the putative ortholog of the Vago cytokine-like factor; this mechanism could be relevant for anti-dengue defense. In Culex mosquitoes, the expression of the vago gene mRNA, induced by WNV infection, requires active viral replication and is dependent on the RNA pathway factor Dicer-2. This factor, in turn, activates the JAK-STAT innate immunity pathway upon viral infection in Culex-derived cell lines and mosquitoes. Also, complementation assays in Culex-derived cell lines demonstrated that the Vago ortholog in Aedes mosquitoes has a conserved functionality (48).
The TRAF-REL2 signaling pathway is also involved in the activation of Vago (47). The Wolbachia endosymbiont can upregulate the expression of Vago in Ae. aegypti; this upregulation may contribute to limit DENV replication (6), suggesting the participation of this pathway in mosquito anti-dengue defense. Therefore, the Vago cytokine-like factor pathway establishes a functional link between a canonical innate immunity pathway (JAK-STAT) and an antiviral immunity pathway based on interfering RNA molecules in mosquitoes.
Discussion
The general picture that emerges from current knowledge on the progression of DENV infection is one where the mosquito responds to the early phases of infection with the upregulation of multiple genes, some notably from the innate immunity system (20). The early immune response to DENV infection is similar to that against other flaviviruses. However, it is important to stress that in infections by other disease-causing viruses, Yellow fever Virus (YFV) and West Nile Virus (WNV), the early immunity response of Ae. aegypti seems to be similar, but the late response could be virus specific (20). As the infection process continues, the virus can induce a shutting down of mosquito defense mechanisms, including the Toll and JAK-STAT pathways, as illustrated by the downregulation of multiple genes participating in these innate immunity pathways (20,62,79).
The basal state of immunity is key to mosquito susceptibility to DENV infection. An extensive comparative transcriptomic study of a variety of field and laboratory refractory and susceptible strains provided evidence for a significant advance in the understanding of DENV-vector interactions (63). The analysis of the midgut global transcriptional profiles of these strains did not correlate with susceptibility to DENV infection. However, the refractory mosquito strains have a higher basal expression of innate immunity genes. This differential expression of immunity genes implies that regulation of the expression of innate immunity genes is a component of vector competence [15], and therefore, it may have an impact on DENV transmission.
The EIP is the time it takes for DENV from the moment it arrives into mosquito midgut to the moment when it infects and replicates within the salivary glands, and the mosquito can transmit DENV to human hosts. Some studies have provided evidence that mosquito innate immune response could have a determinant effect on the EIP (51). This evidence supports the notion that the mosquito innate immune system can be directly relevant for DENV transmission.
Mosquito microbiota can modify mosquito vector competence for viral transmission (29). In particular, infection with Beauveria bassiana increases Ae. aegypti refractoriness to DENV infection (23). The modification of vector competence by microbiota includes the modulation of mosquito innate immune response (23,54). For instance, the activation of the IMD pathway by microbiota in Ae. aegypti has been shown to limit infection by Sindbis virus (8). The introduction of bacterial endosymbionts into mosquitoes represents a successful modification of vector competence that blocks DENV transmission. Wolbachia is a genus of maternally inherited bacterial endosymbionts naturally occurring in many insects, including many mosquito species, but not Ae. aegypti.
However, Wolbachia strains have been introduced into Ae. aegypti (36,78) and the presence of these bacteria renders mosquitoes less prone to become infected and less likely to transmit DENV (12). Although mosquito midgut microbiota can directly affect an antiviral function on DENV (53), there is an important indirect effect through innate immunity modulation in the case of Wolbachia. The mechanism of Wolbachia-dependent mosquito refractoriness to DENV includes activation of the Toll pathway by the reactive oxygen species (ROS) induced by the presence of the endosymbiont in mosquito cells (46). The effector molecules regulated by the Toll pathway contribute to mosquito antiviral response (46), further supporting the role of mosquito innate immunity in vector competence.
Due to the demonstrated effectiveness of the presence of these endosymbionts in limiting DENV infection in Ae. aegypti, mosquito strains are being currently reared and experimentally released to the environment in locations where the toll of DENV transmission on public health is high (18,43). Some other promising experimental strategies to modify Ae. aegypti vector competence for DENV transmission target at least one pathway of the mosquito immunity system (25,79). It is worth noting that successful modifiers of mosquito susceptibility to DENV infection seem to enhance mosquito intrinsic antiviral immunity mechanisms.
Conclusions
The research results summarized in this review indicate that the innate immune response of Ae. aegypti is a critical element of vector competence for DENV transmission.
An important concept is the proposal of the existence of a basal level of immune response activation as a critical component of vector competence (63). This finding led to the conclusion that the genotype and activity level of immunity gene regulatory elements (cis-regulatory sequences) are determinant of Ae. aegypti vector competence (63).
A second important relatively new element is the discovery of an interferon-like pathway centered on the Vago factor. This Vago factor directly connects an RNAi pathway with the JAK-STAT pathway. This functional connection indicates that mosquito antiviral response is a well-coordinated and interconnected network of pathways. This network orchestrates the deployment of molecular effectors directed to limit viral replication in mosquito tissues and cells.
A third relevant element is the confirmation of the participation of the piRNA pathway in the mosquito antiviral response, now conceptualized as a mechanism of a genetically acquired adaptive immune memory. In this mechanism, mosquitoes acquire a viral reference in the form of viral sequences incorporated to their genomes that are later used to produce antiviral RNA molecules in response to viral infections.
These three recent discoveries have generated new questions whose answers will contribute to gain a more accurate picture of mosquito defense and evasion mechanisms against DENV. For example, it will be useful to identify cis-regulatory sequences involved in the basal expression of immunity genes. This identification will allow determining which transcription factor binding sites, and potential polymorphisms within them, contribute to vector competence or refractoriness. Also, it will be useful to know what the degree of variation is and what the allelic distribution of polymorphisms in specific Ae. aegypti populations in high and low transmission locations is.
For a better understanding of the interferon-like antiviral response, it will be useful to identify the specific receptor for the Vago cytokine-like factor as well as the target genes of this pathway. The potential role of the antiviral piRNA adaptive memory mechanism in Ae. aegypti vector competence for DENV requires the assessment of whether natural populations have incorporated DENV sequences in their active piRNA clusters.
In conclusion, Ae. aegypti innate immune response is a crucial component of vector competence for DENV transmission and modifiers that enhance immunity contribute to blocking DENV transmission. Further studies of mosquito innate immune response will provide additional strategies to generate refractory strains that can block DENV transmission in the mosquito.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the following: Consejo Nacional de Ciencia y Tecnología, México, Health Research (FOSISS) Grant 161237, and Basic Science Grant 257990. MC-M was supported by a Postdoctoral Fellowship from Consejo Nacional de Ciencia y Tecnología, México.