
Abstract
We have coined the term “alacrity” to describe the extraordinary diversity of B cell activation potentials, even among cells in a single B cell clone responding to a single antigen. The discovery of methodologies for B cell culture in limiting dilution allowed scientists to identify the source of cellular heterogeneity among cells of the immune system. Analyses of individual B cells set the stage for more detailed descriptions of the factors that diversify B cell functions, some of which will be expanded upon by partner articles in this B cell issue.
The Central Paradigm
A
Immature B lymphocytes have successfully recombined receptor genes, display their antigen receptors on the cell surface, and are selected by antigen for activation, inactivation, or elimination. The portion of lymphocytes that ends up with “unfavorable” receptors may undergo a secondary receptor gene recombination which replaces an inappropriate receptor with a new receptor, a mechanism called receptor editing. T cells also experience successive rearrangements and receptor modifications (19,22,43,62,79), and many immature T cells will perish in the thymus when their receptors respond unfavorably to negative or positive selection signals (83).
In the processes of generating functional B cells and T cells, there is an overwhelming biological apoptotic waste, but this helps the generation of a functionally competent, nonautoimmune repertoire. Several checkpoints exist along the developmental pathway, each examining the competence of new receptors to ensure the generation of functionally responsive, mature B cells expressing nonself receptors.
B cell immunology encompasses such a vast field of studied processes that experts in B cell and T cell fields may find themselves isolated from one another. The situation differs from the past when the two “immunologies” were solid partners, particularly when “T cell help” for B cells was first discovered. In fact, when one goes back to those days of first discoveries involving B and T cell cooperation, one finds the first discussions of functional cell heterogeneity, repertoire, memory, tolerance, suppression, clone size, allelic exclusion, and many other features.
The work presented below is neither a review article nor an unbiased report, it is rather a compilation of laboratory quests performed in an environment of academic freedom addressing issues, which were considered most important at that time. Projection from the past gives us assurance that once we understand where we are coming from, we know where we are heading.
Did It Start with Niels Jerne, or Perhaps with Sir Macfarlane Burnet?
Niels Jerne's “Natural selection theory of antibody formation” (26) became one of the most influential articles of the late 1950s of the last century. Jerne postulated that a full repertoire of antibody is present in the organism before any antigen contact, and antigen instead of serving as a template for antibody formation merely selects the fitting preexisting antibody. Although Jim Watson browsing through the draft of the article commented, “it stinks,” it was Sir Macfarlane Burnet who recognized that Jerne's idea of “preexisting” antibody repertoire provides an ingenious turn in understanding the basis of the immune system.
Burnet was ready to pave the road of modern immunology by extending Jerne's theory to conclude that the antigen selects the cells expressing that given antibody rather than selecting free antibody (6) [republished (8)]. This gave birth to the clonal selection theory (6) [republished (8)] (7). The cells are monospecific and are selected by antigen and are induced to expand clonally to produce antibody of a single specificity.
It was predicted that there were ∼1020 different antibody molecules that could be made in man [1017 in mouse; today, theoretical and actual repertoire sizes remain topics of continued debate (18)]. A postulate was coined: 1 cell = 1 antibody. The dichotomy of the immune system was that it could be considered as a collection of independently operating lymphocytes (28), but at the same time there were interactive associations among all immune relevant organs—spleen, lymph nodes, blood, skin, mucose, Peyer's patches, bone marrow, and thymus—in which immune functions were taking place.
This was obviously an undue oversimplification, but it enabled the quest for a system in which the complexity of the body could be disregarded. Therefore, methodology based on the use of lymphocytes (and some accessory cells) taken out of the context of the organism formed the proper starting point to study the basics of the immune system.
This reductionist view paved the path to the in vitro technique of Mishell and Dutton (68) for inducing primary antibody responses, where the organs harboring the lymphocytes could be disregarded. Evidence that the response was initiated from a small pool of cells precommitted to respond to the inducing antigen was obtained by a fluctuation test—developed by Luria and Delbruck (56) for studying mutations in bacteria and adopted for studying immune cells (45). The test also accommodated the notion that the response was ensued by clonal expansion of precursor cells (74). Several additional strategies for B cell analyses followed (12,33,34,37–40,61, and 73).
Upon supplementing B cells with T helper (Th) cells or with soluble factors from T cells, it was established that the induction of the response occurred upon cooperation of the two cell types, that is, of B cells and T cells (67). Quantitative aspects of the response, especially estimates of the frequency of cells involved in the response were established by in vitro limiting dilution analysis (LDA) (76). Single hit kinetics was observed when responses were induced in many small culture-aliquots, under such conditions one cell population was kept limiting and other cells required for the response were present in abundance.
In vitro responses were worked out in some details for the mouse system, but soon followed attempts to induce the response using rabbit lymphocytes, chicken, frog and finally human cells. Although “all-or none responses” under limiting dilution conditions provided a satisfactory evidence of the clonal homogeneity (of cells and their products), the actual proof of homogeneity came from determining the allotype of antibody (58).
Alacrity
Although this work is on antibody, it is also on a special feature of cells involved in the immune system, which we name “alacrity” (50). The basic notion is that in terms of molecular composition, no two cells are identical—neither within the organism nor across the entire species. Cells with “equal function” and “equal specificity” still differ in many components of their cellular inventory (49). Even cells with seemingly identical receptors and surface markers differ one from the other in molecular terms. The differences might be small or large; nevertheless, they translate into distinct eagerness and preparedness of cells to respond to a given stimulus. For this eagerness and preparedness of cells to respond, we coin the term “alacrity.”
Materials and Methods
Methodologies for B cell cultures are given in the original article of Mishell and Dutton (68), while the procedures for the LDA are described in the initial article by Lefkovits and Quintans (45,74) and then in all details in the monographs by Lefkovits and Waldmann (53,54). The use of plaque-forming cell (PFC) methodology is described in the original article of Jerne and Nordin (31) and in the detailed review by Jerne et al. (30).
Large scale 2D gel electrophoresis of proteins from lymphocyte populations was adopted from Anderson and Anderson (1) and evaluated by Kepler image analysis system (55).
Results
In vitro antibody response
A new platform for exploration of the immune response was launched by a method for inducing primary antibody responses in vitro (68), through which it became possible to dissect the immune response into discrete steps while disregarding the complexity of the entire organism. The methodology was worked out for studying mouse spleen cells, using sheep red blood cells (SRBCs) as antigen. The cultures were performed in 1 mL vessels with an input of several million cells. The actual response allowed scientists to identify cells involved in the response by enumerating antibody-forming cells using Jerne's plaque method, which was enthusiastically embraced by the scientific community just a few years earlier (31).
Upon incubation of cells (under delicately balanced culture conditions) for 4 days, typically several 100 antibody-forming cells were detected in each antigen-stimulated culture, while in the absence of added antigen, only a few background antibody-forming cells were found. That the antibody-forming cells were derived from B cells was already established in several articles (15), but the requirement for T cells in the response was substantiated later (69).
Whether the resulting response was due to direct conversion of precursor cells into antibody-forming cells or alternatively due to clonal proliferation of cells from a small number of precursor cells was by no means certain at the time.
Luria-Delbrück fluctuation test
The clonal origin of the antibody response has been well established for decades, but it was by no means the case in the 1960s (e.g., when B cell experiments were conducted at the 1967 EMBO (European Molecular Biology Organization) course in Frankfurt (68), where the “premiere” of the Mishell-Dutton culture system occurred). In early days of modern immunology, upon Jerne's natural selection article (26) and Burnet's clonal theory article (6) [republished (8)] (7), the issue was still under debate. If the cell is specific to synthesize antibody of a single specificity, then it must be a “rare cell,” since each of many specificities has to be represented at least by a few cells of the population. Also, if enough cells are supposed to be available for antibody production, then clonal expansion of stimulated cells is indeed required. Therefore, the question of the frequency of specific cells became of foremost importance.
The Mishell-Dutton system became widely used, and the well-known Luria-Delbrück fluctuation test offered itself to be adapted to study the cells of the immune system, to answer two questions: (a) whether or not the response is initiated by a few rare cells and (b) whether or not clonal proliferation occurred.
Spleen cell suspensions (from mice) were partitioned into many small aliquots (microcultures) under previously defined Mishell-Dutton system conditions, and the input numbers were adjusted such that it was possible to score all-or-none responses in the microcultures. Indeed, once the “partitioning” was tuned (narrowed down) to a cell input of 105 cells or less, a certain portion of cultures responded while others failed to do so. The Luria-Delbrück fluctuation test is meaningless in situations when “all cultures” are positive; therefore “inadequate” partitioning, as with 106 cells/culture well, yields no information. Experiments were performed in so-called Terasaki plates, with 60 microcultures of 10 μL volume each.
Both the number of responding cultures and the number of antibody-forming cells in each microculture were determined. As was the case in the conventional cultures, responses occurred only in cultures with added antigen, and remained nil in the absence of added antigen (thus confirming that the presence of antigen is a prerequisite for a response).
By finding substantial numbers of antibody-forming cells in some culture wells and none in others, the rare cell issue seemed to be established, at least in principle. Nevertheless a direct proof was still missing. The objection could be raised that cultures that failed to respond, failed due to (uncontrolled) inadequate culture conditions in certain culture vessels rather than due to the absence of specific cells. This argument was resolved by using a mixture of two antigens (SRBC and goat red blood cell); the responses were independently assorted such that the negative cultures to one antigen still yielded positive responses to another antigen—thus the conditions were amiable to the response, and negativity must have been due to the absence of a responding cell (45). The responding cultures contained as a rule many antibody-forming cells. Furthermore, as expected for random assortment, some cultures were double positive and others were double negative (Fig. 1).
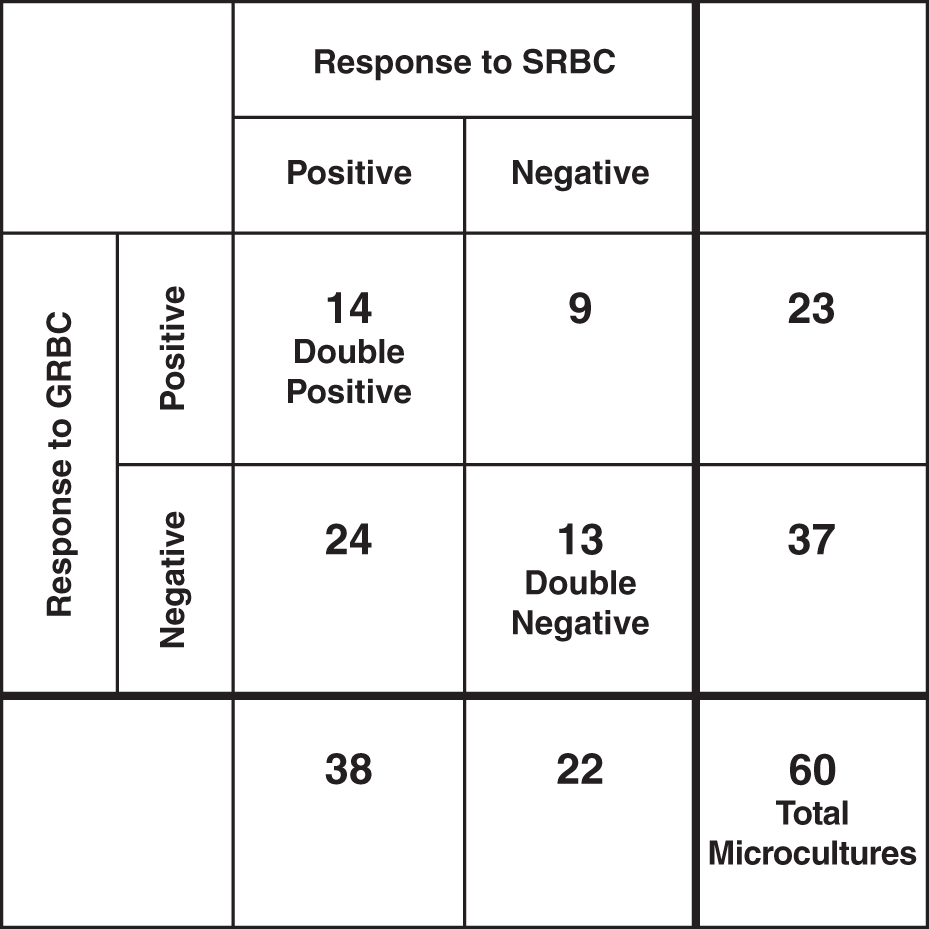
Primary in vitro stimulation of spleen cells with GRBC and SRBC. Sixty microcultures were tested separately for the response to GRBC and SRBC (45). A two by two table is shown (SRBC, GRBC): (a) Twenty three cultures responded to GRBC and 38 to SRBC. (b) In several cultures negative for GRBC (37 wells), a response to SRBC did occur (24 wells). (c) In several cultures negative for SRBC (22 wells), a response to GRBC did occur (9 wells). GRBC, goat red blood cell; SRBC, sheep red blood cell.
Requirements for cell cooperation
Above arguments did not take into account requirements for cell cooperation. The requirement for T cells to cooperate with B cells for eliciting an antibody response was foretold by the work of Miller (65) and indicated by several other in vivo and in vitro experiments (66). It was observed that whenever T cells were depleted from a population of lymphocytes, the response was seriously impaired. Such a T cell depletion was achievable by a treatment of a cell populations with anti-Thy1 serum and complement (77). This allowed scientists to perform in vitro studies using “treated” populations as a source of B cells. Similarly cells from thymusless (nude) mice were convenient sources of B cells. The studies were performed either in bulk cultures (46) or in microcultures (74). The experimental protocol for B cell responses had to be planned such as to ensure that those wells that scored negatively were indeed lacking precursor B cells, rather than an accessory component.
LDA: single-hit kinetics
Once the microculture experiments were refined to provide frequency estimates of specific B cells, three experimental conditions had to be ensured: (a) adequate numbers of T cell help (or adequate amounts of soluble T cell-replacing activity) had to be present in each culture (dubbed not entirely correctly as “saturated conditions”); (b) graded numbers of B cells had to be chosen such that a satisfactory spread of cell input was preserved—preferentially such that at low input at least some culture wells missed the putative specific precursor cell (i.e., they will score truly negative); (c) the resulting titration curve exhibited single-hit kinetics which was the prerequisite for accurate Poissonian scoring.
The experiments had to be composed of a large enough number of culture wells, such that for each experimental point a meaningful confidence interval (95% confidence limits) would permit distinguishing single-hit from multihit events. Results of an experiment for estimation of the frequency of antigen-specific precursor B cells in nude mice are depicted in Figure 2. This particular experiment was aimed at establishing the response (all-or-none) for six B cell inputs (0, 2 × 104, 4 × 104, 6 × 104, 8 × 104, and 1 × 105 cells/well). Sixty microcultures of 10 μL in volume were used per experimental point, thus 360 microcultures in toto. Negative controls were performed as well—they were nonresponding. The Poissonian plot was based on the portion of negative cultures. Increasing graded numbers yielded a decreased portion of negative wells.

Estimation of the frequency of antigen-specific precursor cells in nude mice. This figure is reproduced with permission [from Quintans and Lefkovits (74) Copyright Wiley-VCH Verlag GmbH & Co.].
To reiterate, it was ensured that the lower input was indeed at the fluctuation level (limiting), and accessory cells as well as antigen were at a saturating level. The intercept of the titration line with the level of 37% nonresponding cultures indicated the number of cells, which on average contained one precursor cell. The intercept was at 47,000 cells (frequency estimate 1/47,000 or 2.1 × 10−5). The theory behind these measurements is given by Lefkovits and Waldmann (53,54).
The frequency of specific cells for a number of antigens tested was of the order of magnitude 1/100,000 in cell populations from nonimmunized mice. These frequencies increased dramatically upon mouse immunization. These figures are useful for predicting the size of the repertoire. This issue, particularly as it relates to memory cells, is taken up again in the Discussion section.
LDA: multihit and leveling off
Although we have always considered the presented work as a procedure for a single-cell analysis, the notion of “a single cell” was only a conceptual one, since in reality, the studies were performed in “partitioned” cultures containing a multitude of cells or were designed to observe cells in a defined relationship with other cells (filler cells, bystanders). We learned how to tune the experimental conditions such that the response would be limited by a single component. We knew what was missing, and we supplied exactly what was missing. The “gold standard” was a single-hit kinetics (straight line in a Poissonian plot) and when we achieved it, firm conclusions were drawn.
But what about all those curves which displayed multihit or multitarget titrations or a leveling off? The common feature of all titration curves that deviated from linearity was that a portion of specific cells failed to respond. To reiterate, the cells were there, but they remained idle. This is shown in two semilog plots, one related to a multihit titration curve (Fig. 3A) and one to a leveling off (Fig. 3B). Although we use the term “deviation from linearity,” the common feature is that graded numbers do not yield graded responses, at least not in Poissonian terms.
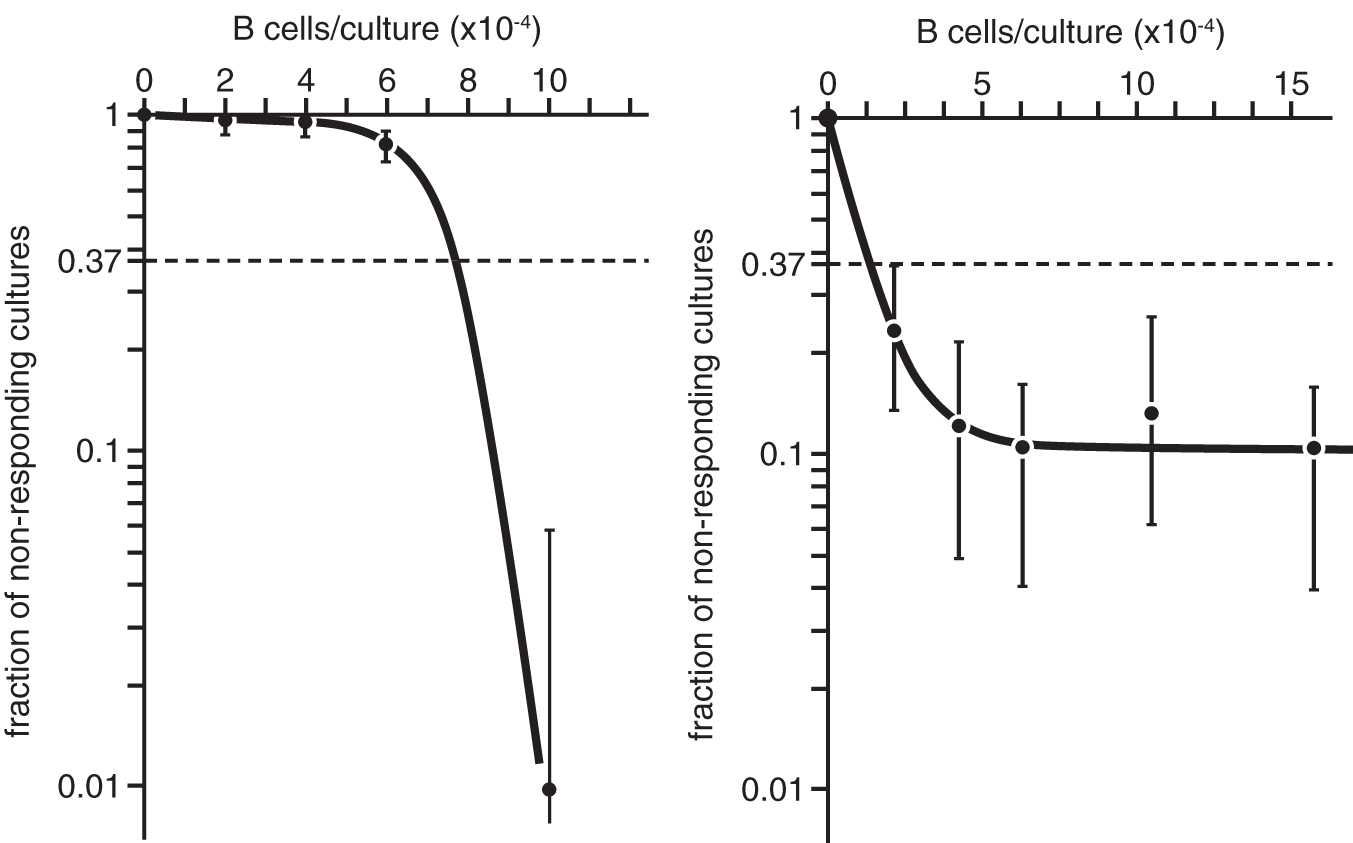
Titration curves deviating from linearity.
The “multihit” event and the “leveling off” are two cases, where some components are not adequate either at low or at high cell input. If we detect a good response at high cell input, there must be a reason why at low input the cells remain idle. If we detect a good response at low and intermediate response and it stagnates at high input, there must be a reason for it as well. In most of our work, we aimed at supplementing the “system” with cells that allowed for full potentiality of the response.
Only decades later we realized that behind these deviations from linearity were not only inadequacies of third party cells (usually help or macrophage function) but also inadequacies in B cell activation potentials. This will be considered below in some detail.
Syngeneic and allogeneic cell interactions
Introduction of the Mishell-Dutton technique enabled a novel type of experimentation with both primary and secondary immune responses, and it also enabled scientists to contemplate new questions—(a) Are T cells required for the initiation of the response? (b) Are T cells needed to induce clonal expansion of B cells? (c) Is the course of the B cell response (time point of the peak) dependent on the amount of helper activity? (d) Is there any correlation between the amount of help and the magnitude of the response?
Experiments described below were based on the classical Mishell-Dutton protocol for bulk cultures (1 mL), such that a constant number of nude spleen cells was complemented with allogeneic cells (nude mice were at various stages of backcross to C57BL/6 background, while normal NMRI mice [an outbred mouse strain] were used as a source of allogeneic cells; NMRI mice are rarely used today). An explanation related to “allogeneic cells” is needed: admixing T cells from unrelated strains of mice provided a considerably more vigorous help than admixing syngeneic T cells. This observation was made decades before the T cell receptor issue was resolved, and in most laboratories, the use of allogeneic T cells was simply for convenience. It was more opportune to supply “saturating” numbers of T cells with allogeneic cells than with syngeneic cells.
Figure 4 depicts results of such a complementation experiment demonstrating that the dose of allogeneic T cells (added to the nude spleen cells) dictates the time point at which the peak response occurs. We chose a wide range of allogeneic cells covering four orders of magnitude and measured the responses in terms of antibody-forming cells on days 4 and 5. The addition of 105 allogeneic cells per culture yielded ∼3,000 PFC on day 4 (black circles) and 5,000 PFC on day 5 (open circles). The addition of 5 × 105 allogeneic cells yielded optimal responses on day 4, and declining responses on day 5.
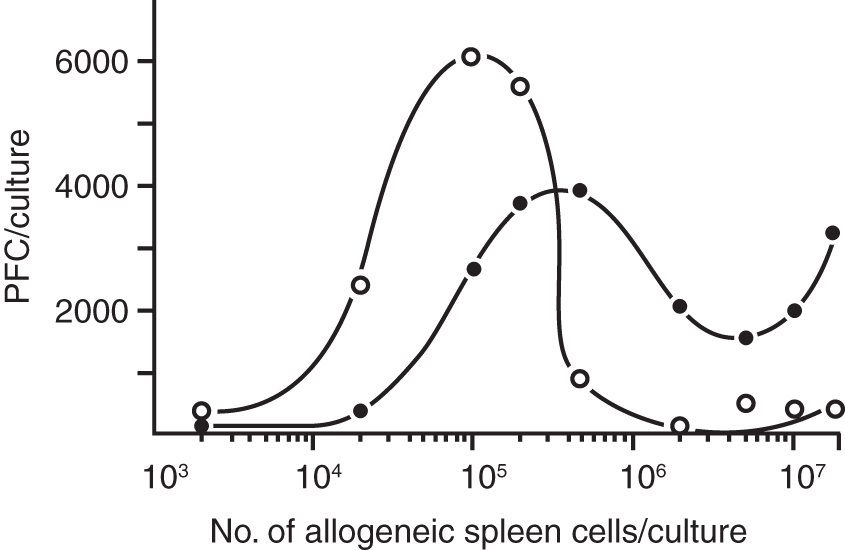
Reconstitution of antibody responses of nude spleen cells by complementation with a range of allogeneic cells, closed circles (day 4), open circles (day 5). This figure is reproduced with permission [from Lefkovits (46) Copyright Wiley-VCH Verlag GmbH & Co.].
Since the low input of added allogeneic cells turned out to be favorable for building up a vigorous response, we asked the question of whether admixing allogeneic T cells either at time zero, or with a delay of 24 or 48 h altered the time of the peak response. Figure 5 indicates that immediate addition or a 24 h delayed addition of T cells yielded a normal response, while additions beyond 24 h delayed the peak response.
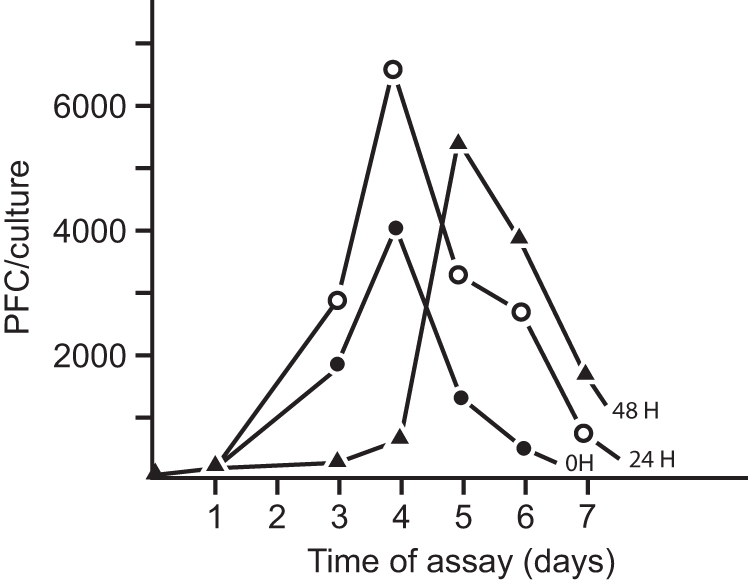
Antibody response induced through delayed addition of allogeneic cells, closed circles (addition 0 h), open circles (addition 24 h), losed triangles (addition 48 h). This figure is reproduced with permission [from Lefkovits (46) Copyright Wiley-VCH Verlag GmbH & Co.].
Although there was a positive correlation between the number of added T cells and the magnitude of the response, there was a limit beyond which an increased dose of allogeneic cells displayed inhibitory activity (we used a neutral description of “inhibitory activity,” since at that time regulatory T cells were not in parlance).
From the above experiments, we conclude that T cells are not required at the initial stage of the response, although are needed to ensure clonal expansion (48). Since the early days of modern immunology, we said “the clone breeds true,” and assumed that during clonal proliferation, each daughter cell was a true copy of the parental cell [except for somatic mutations which can modify/improve antigen binding (63,64,88)]. This assumption concerned the cell's specificity, but not the cell's activation potential. We shall elaborate on this issue when discussing the portion of the clones called “vegetative pool.”
Signals for clonal expansion
The finding that T cells were not required at the onset of the response called for further enquiry into the succession of signals governing B cell activation. Mitogens-like lipopolysaccharide (LPS) and Pokeweed mitogen offered themselves as convenient tools to study this issue (75). Summarily, we found that signals delivered to B cells by mitogens provided a nonspecific signal that “turned on” the cells to enter proliferation. Such proliferation, if not followed by further signals, ceased after a few cell divisions. A portion of these cells (probably blast cells) had the capability to produce antibody and were readily detected as PFCs. Early claims (2) that mitogens could support an indefinite growth of cells turned out to be unsubstantiated.
In the Mishell-Dutton experimental setup, LPS-stimulated B cells yielded early peaking responses (at day 2 or 3) of small clone sizes, while upon addition of helper activity (soluble or via allogeneic cells) a prolonged lag phase was observed, followed by a significantly augmented response peaking on day 4 or 5. Our interpretation was that during the lag phase, a “vegetative pool” built up (48,75). These were proliferating cells that had not yet secreted antibodies, but that could ultimately become plasma cells (antibody-forming cells [AFCs]). We concluded that cells receiving only the LPS-signal entered proliferation and then became early AFCs and thereafter decayed. Cells that received an LPS-signal plus adequate T cell help remained longer in the pool of proliferating cells (the cryptic—”vegetative pool”), and became AFCs after several rounds of proliferation, thus yielding substantially larger clone sizes.
There were many interesting propositions as to how B cell and T cell populations should be manipulated in vitro, separately and independently. As given in the review article on “Precommitment in the immune system” (47) the methodology for obtaining discrete cell populations was based either on the physical separation of cells (antigen-binding columns, nylon wool, and velocity sedimentation) or by killing subsets of cells by radioactive tagging or alternatively by treatment with anti-θ and complement—and using the noneliminated population for experimentation. Not surprisingly, interpretations of these results were often conflicting, since protocols (and reagents) differed from laboratory to laboratory.
Mouse, rabbit, chicken, frog, fish, man
Once Dutton's laboratory was successful in developing the in vitro antibody response for the mouse species, attempts in laboratories around the globe followed, using cells from other species. It turned out that a straight forward implementation of the “master protocols” was not possible. The Mishell-Dutton system utilized fetal calf serum, and not surprisingly, only some batches “worked.” None of the “good-for-mouse system” calf serum worked for inducing antibody responses in lymphocytes of other species. The first truly successful protocol for culturing cells from peripheral blood lymphocytes (PBL) of rabbits was worked out by Luzzati et al. (58). Work with other species followed and finally protocols for the culture of human PBL were established (57,60). Human tonsils (3), chicken cells (71,72), and frog cells (14,85) were used. Just for the sake of objective reporting, cultures using organ fragments (38 –40) were developed as well, and they were often more successful than cultures using cell suspensions. A major breakthrough came when Norman Iscove established serum-free culture conditions (24). Induction of primary antibody responses of fish lymphocytes was not successful, but propagation of populations producing antibodies was reported (44).
Allotype homogeneity in individual culture wells as evidence of clonal products
As mentioned above, the microculture system was originally developed for mouse spleen cells. Adaptation of the methodology for cells from other species was of some importance, since several topics, which could not be addressed using mouse species, could be addressed using other species. Luzzati and Pernis focused on the rabbit system for several reasons. First, this species supported experimentation with peripheral blood cells and allowed long-term, longitudinal sampling from each individual. Second, the laboratory of Luzzati possessed expertise on the study of b4/b6 allotypes of the heterozygous rabbit strain, which provided a clue toward solving the issue of antibody homogeneity in the microculture system (59).
The most important finding was obtained from microcultures of PBL of red blood cell (RBC)-primed rabbits heterozygous for b4/b6 allotypes (Fig. 6). For each test culture, an agar-coated microscope slide was prepared with three holes made as assay wells. Anti-b4 antisera and anti-b6 antisera were placed in top and bottom assay wells, respectively, and the tested supernatant was placed in the middle well. When RBCs were overlaid, the pattern of lysis indicated allotype (for a b4-producing B cell, RBCs were lysed on the top half of the gel and for a b6-producing B cell, RBCs were lysed on the lower half of the gel). Results demonstrated that (a) B cells usually expressed no more than one antibody, and (b) B cells generally used either one chromosome or the other, but not both, for antibody expression (allelic exclusion).
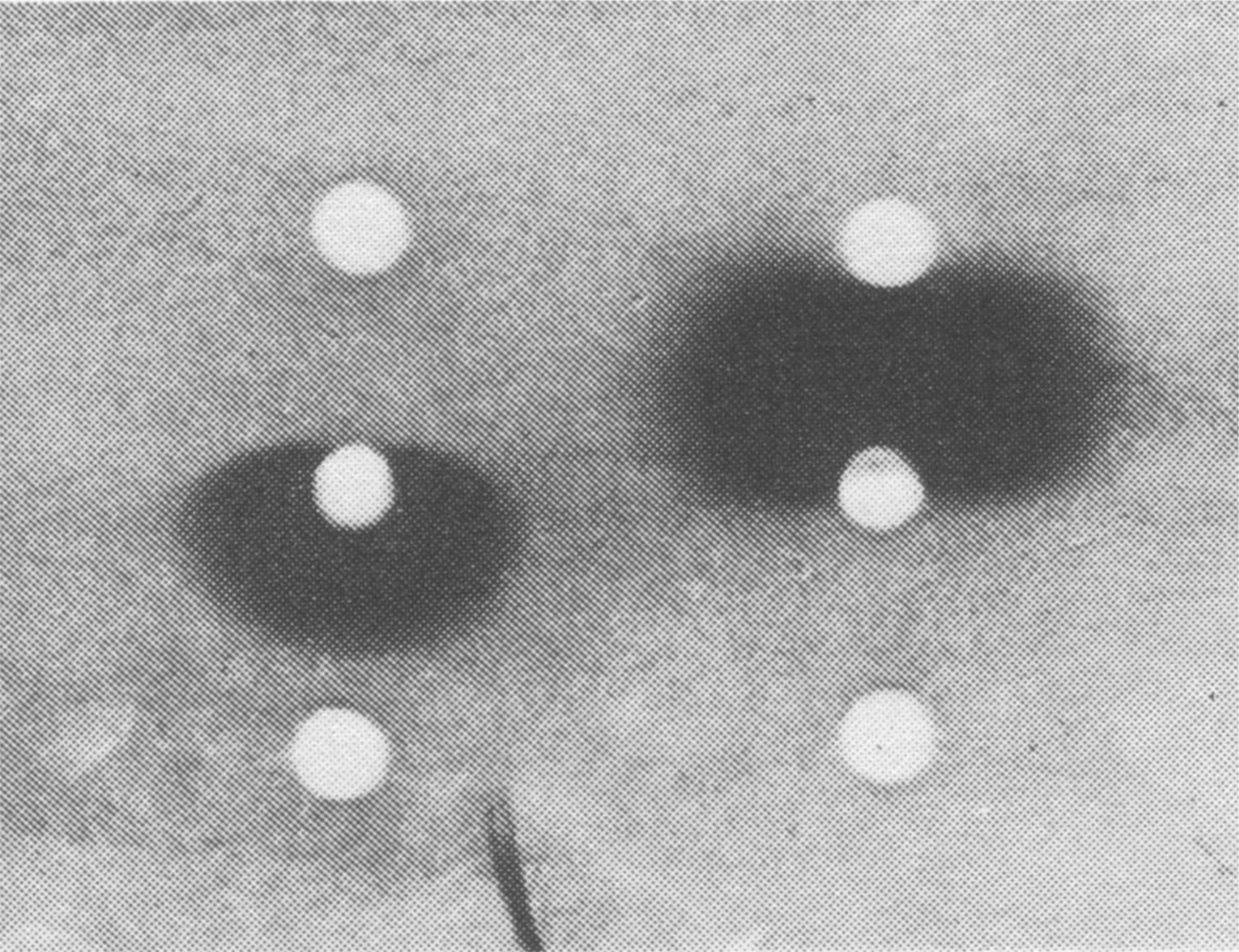
Test of the allotype of the clonal products; inhibition pattern of two clonal products. Top wells contained anti-b4 allotype serum; central wells contained test culture fluid; bottom wells contained anti-b6 allotype serum. This figure is reproduced with permission [from Luzzati et al. (59) Copyright Wiley-VCH Verlag GmbH & Co.].
Besides the fact that the results unequivocally demonstrated the clonal character of the products of the microculture wells in these experiments, they reassured scientists in more general terms that Poissonian sampling reliably indicated the clonal nature of a response, regardless of the experimental setup.
Heterogeneity of B cells reacting with T cell factors
Under the conditions of limiting dilution, it seemed that one T cell cooperated effectively with a very low number of B cells; in fact, a single B cell would be the best guess. These (somewhat esoteric) issues were debated in the late 1970s, but it was just a conjectural argument, often using the catchphrases: “kiss and go,” or “kiss and die.” In the late 1970s of the last century, there was among scientists a fallacy argument related to the so-called antigen-specific factors. The argument was that since the B cell response was a specific event, the soluble components that helped B cells were also antigen-specific. This turned out to be false. Nevertheless the population of responding B cells displayed a special profile of heterogeneity, such that cells responding to soluble helper factors behaved as if there were matching subsets of B cells and T cell products. Supernatants derived from mitogen-induced cells when added to samples of B cells (that contained antigen-specific B cells and antigen) were not able to provide help to all replicate wells (52). Only decades later, an explanation for this interesting finding was found to be based on B cell activation potential, not antigen specificity, as will be presented in more detail in a later section of this article.
T suppressor cell titration: frequency estimates
If cell populations were available that contained suppressor elements and no helper activity, LDA titration would be just a “mirror image” of the T helper cell titration. All responding cultures would be Poissonially negative (not containing suppressor cells). Admittedly, such a constellation (of pure suppressor cells) was not available—it was always a mixed population. In the experiments initiated by Corley and Kindred, it was contemplated that among the mixed-lymphocyte response-primed cells, there must be some kind of suppressive activity (10). In fact, suppression was so dominant that in bulk cultures no anti-SRBC response could be elicited (since a few suppressor cells dominated the entire population). Partitioning into microcultures apportioned the suppressor cells into a few microculture wells. The titration curve shown below allowed researchers to distinguish help from suppression and allowed an estimation of T cell frequencies. In the iconic experiment, results of which are depicted below (Fig. 7), the frequency was fTh = 1/2,000 and fTs = 1/20,000

Estimation of the frequencies of helper and suppressor cells in a population containing both cell types. This figure is reproduced with permission [from Luzzati et al. (59) Copyright Wiley-VCH Verlag GmbH & Co.].
This “mix” of help and suppression had previously raised a suspicion that suppressor (Ts) activity might be a phantom, and that “too much help is suppression.” In the experiments described above, the two T cell entities were clearly disentangled.
Activation of B and T cells by macrophage populations
In most LDA experiments, it was feasible to ensure that one cell type was limiting, and all other required entities were in excess. Only such titrations seemed to give meaningful results, especially when it came to frequency estimates. The intention of experiments which we called “partition analysis” was to identify cells that apart from the titrated population were needed for the response, with special attention to the macrophage population (35). Adherent peritoneal cells were established in the Terasaki plates, and either T cell or B cell stimulation was induced. It turned out that two types of accessory activities could be established. When spleen cells from BDF1 mice were supplemented with Concanavalin A and T cell growth factor, an adequate accessory cell activity for T cell stimulation prevailed, while if spleen cells from BDF1 mice were supplemented with LPS and dextran sulfate, the accessory activity favored B cell stimulation. Interestingly, the failure of B cell activation in some cultures was later traced to the presence of suppressor cells.
Epitopes: dinitrophenol, trinitrophenol, phosphorylcholine, carbohydrate, arsonate
The terms “epitopes” and “paratopes” were coined by Jerne (27). In everyday parlance, epitopes were meant to be portions of antigens that were recognized by paratopes—structures on the variable portion of antibody molecules. Later, the usage became less precise (especially when extended to T cell epitopes), but in B cell immunology, “epitopes” meant antigenic determinants usually in the context of carrier molecules. It was a common knowledge that antibodies were capable of recognizing both small and large molecules in a specific manner, but when small molecules (haptens) were used to elicit B cell responses, they had to be tagged to a large molecule. Thus haptens had to be coupled to a carrier. Conveniently, red cells were good carriers, and haptens were easily attached to these carriers either covalently or by adsorption.
It turned out that some of these hapten-carrier combinations were useful for induction of antibody responses both in vivo and in vitro. Usable haptens were dinitro- or trinitrophenol, phosphorylcholine (PC), and also various types of carbohydrates. Frequency estimates were obtained for various haptens (4,11). Of special interest were anti-PC antibodies, since all responding clones expressed a restricted idiotypic property (binding TEPC-15 myeloma). Interestingly, pneumococcal antigen was T-independent in vivo, while it required T cell help in in vitro stimulation.
Selective recruitment of recirculating lymphocytes
Thoracic duct lymphocytes (TDL) were not only a convenient source of cells retrieved from the circulation of the organism, but they also were a useful indicator of events occurring during the ongoing immune response in the intact animal. Thus, when TDLs were placed in microcultures with antigen for 5–6 days, they responded and produced antibody. Preinjection of SRBC into the animal 24 h before collecting TDL altered the in vitro response to SRBC and horse red blood cell (HRBC). It prevented a specific anti-SRBC to occur, while allowing a normal third party response to HRBC to occur. Various studies of timing, addition(s) of helper factors, depletion of certain cell types, and reoccurrence of the responsiveness at later stages lead to the conclusion that the specific unresponsiveness of the TDL was the result of selective recruitment of specific B precursors from the recirculating lymphocyte pool (78).
Affinity, avidity, and repertoire
The degree of the molecular attraction of an epitope to the structure of paratope (antibody combining site) is referred to as affinity. Affinity describes how strong two molecules bind to each other. Affinity is taken to be the equilibrium association constant, characterizing the paratope-epitope pair. Formally, it is the reciprocal of the concentration of epitopes at which half of the paratopes will be occupied.
Since the interaction refers often to binding molecules consisting of several epitopes, the overall capacity to antibody-antigen binding is referred to as avidity. The most famous dissertation publication on avidity is by Jerne (25) with a title “A Study of Avidity Based on Rabbit Skin Responses to Diphtheria Toxin-Antitoxin Mixtures.”
The repertoire of antibody is the number of different paratopes available for interacting with the spectrum of antigens. It was assumed that the repertoire of paratopes (in the mouse system) was in order of 107 molecular specificities, meaning that the sum of all antibody molecules of an individual is capable of binding with “perfect fit” to 107 complementary epitopes. Since the binding is degenerated (one antibody can bind to several spatially similar structures) and since the interaction is due not only to affinity but also avidity, it was assumed that the repertoire of the size of 107 covered an antigenic spectrum of at least 108 different structures. The repertoire of an individual was considered “so-to-say” complete meaning that it could recognize most antigens in nature (before tolerance or other forms of repertoire modeling), but it was less complete than the repertoire of the entire species.
In most instances antigenic stimulation gave rise to antibody molecules with a wide range of affinities and avidities, and only after a prolonged maturation (repeated antigen exposure) the response narrowed down to more avid antibodies. Interestingly, some antigens (e.g., PC) induced antibody of a narrow range of avidities—thus yielding antibody of restricted heterogeneity.
Alacrity
The immune system does respond not only to a broad spectrum of antigenic assault, but does so both under favorable and under adverse conditions. We have introduced the concept of alacrity in the context of a special kind of cell heterogeneity that dictates whether a cell will engage in the immune response or not (50).
While affinity and avidity are pointing toward the strength of the binding to epitopes, alacrity is directed toward the physiological milieu of the response. Alacrity of a cell (or a cell population) relates to eagerness, or readiness to respond under given boundary conditions, sensing the physiological environment (molecules interacting with the membrane, molecules trafficking through membrane channels, osmotic pressure, pH, relative proportion of other cells, cell density, and so on). The alacrity ensures that whatever physiological conditions prevail, some ligand-specific cells will be ready to engage in the response.
A nice demonstration of alacrity was obtained upon titrations based on a broad range of additions of essential amino acids, spanning from amino-acid-starvation to a poisonous overdose. The titration of phenyl alanine revealed that with the highest added dose (10 × ), the clonal response dropped to a level of only 3 responding cultures out of 60 (instead of 58/60 at the standard concentration). It was not at all surprising to observe that a dose exists which is completely inhibitory (90). But what about those cultures yielding a good response under these adverse conditions? Today we interpret the results such that among the population of precursor cells, there were a few cells with sufficient alacrity to engage in the response under adverse boundary conditions; others remained idle.
Numerical taxonomy of clones
Advances in the field of monoclonal antibodies distorted our perception by suggesting that learning on one B cell was learning on all B cells. We wished to strengthen the notion that B cell heterogeneity is more than a mix and superimposition of monospecificities. We used and utilized the high-throughput proteomic analysis of clones of lymphocytes, quantified a large number of molecular components, and established the clonal relatedness applying Principal Component Analysis (51).
Clonal taxonomy showed that some clones were related to each other not only in terms of their molecular inventory but also in terms of their ability to respond. The numerical taxonomy approach might become an adequate basis for establishing a mathematical model of alacrity. One such comparison of clones is given in Figure 8.
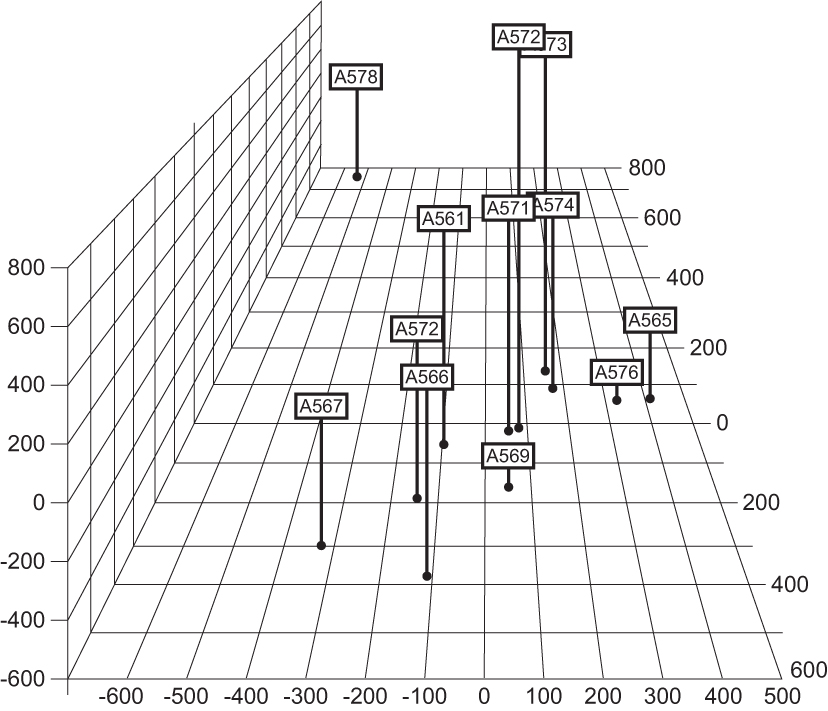
Establishing clonal relatedness applying Principal Component Analysis in high-throughput proteomic assay. This figure is reproduced with permission [from Lefkovits et al. (51); PNAS].
Discussion
Although the initial aim of the microculture methodology was estimation of frequencies of immune cells, it offered gratuitously data on other quantitative aspects of cellular interactions, among others, the clone sizes. In a somewhat doctrinaire approach, we concentrated on processes that yielded single-hit kinetics, other processes we have mostly neglected. We interpreted “V shaped curves” as a productive separation of helper effects from suppression activities. Plots of Fo and of F+ yielded frequency estimates for helper and suppressor cells, respectively. In most cell populations containing a mix of help and suppression, the frequency of helper cells was substantially higher than the frequency of suppressor cells—often by an order of magnitude (fTh = 1/10,000, fTs = 1/1,000). Suppressor activity (despite lower frequency) was always found to be dominant over helper activity. We were aware of the fact that we would fail to recognize instances in which the frequency of Ts would be much higher than the frequencies of Ths. It is well possible that failed attempts to induce immune responses (in human and rabbit PBL cultures) were often due to the presence of dominating suppressor cells. “Leveling off” indicated that the titrated cell population could not come to full expression at high cell input, because some components required for the response were missing. “Multihit” curves were indicative of a disproportionally poor response at low cell input. In most instances, we were able to identify the missing component and thereupon obtain single-hit curves. Decades later, we reconsidered all those “strange” results and realized that they were well in agreement with the alacrity hypothesis.
When B cells were brought in contact with antigen in the absence of added T cells, an abortive response occurred yielding a small clone size, while in the presence of T cells, an antigen-specific clonal proliferation of B cells occurred and a hidden vegetative pool was created (48,75). At the progression of the response, cells exited the vegetative pool and became antibody-forming cells. An early exit of cells from the vegetative pool caused an early appearance of PFCs and a small clone size. Delayed exit allowed expansion of a large vegetative pool, yielding a vigorous response of large clone sizes. The vegetative pool was a hidden portion of the clone (mostly not detectable as antibody-forming cells), thus (in our view), PFC counts were not an indication of clonal growth, but rather of a clonal decay (each antibody-forming cell depleted the vegetative pool). With Geoffrey Hoffman, a former member of the Basel Institute, we constructed a computer model, in which the clonal response was a function of a single variable p—a probability that the daughter cell left the vegetative pool (and became an end cell, memory cell, or it apoptotically died). With low p values, the clone size kept increasing, and when p > 0.5, the clone declined (48). Figure 9 indicates two constellations of clonal proliferation. As shown in this figure, as long as the cell remains in the proliferating pool, it expands (full dots), but upon changing to a mature cell (memory cell, or obtaining another physiological signal), it is lost from the pool. At the right portion of the figure a situation is depicted, in which a cell upon undergoing the first and second round of cell division terminates the clone. The lower pictures show that an increasing p-value depicts the clonal progression up to p > 0.5, thereupon the clone decays. Incidentally, this model is in principle the same as for a nuclear chain reaction (the reaction has to be moderated (through a neutron moderator) to maintain the reaction and prevent explosion)
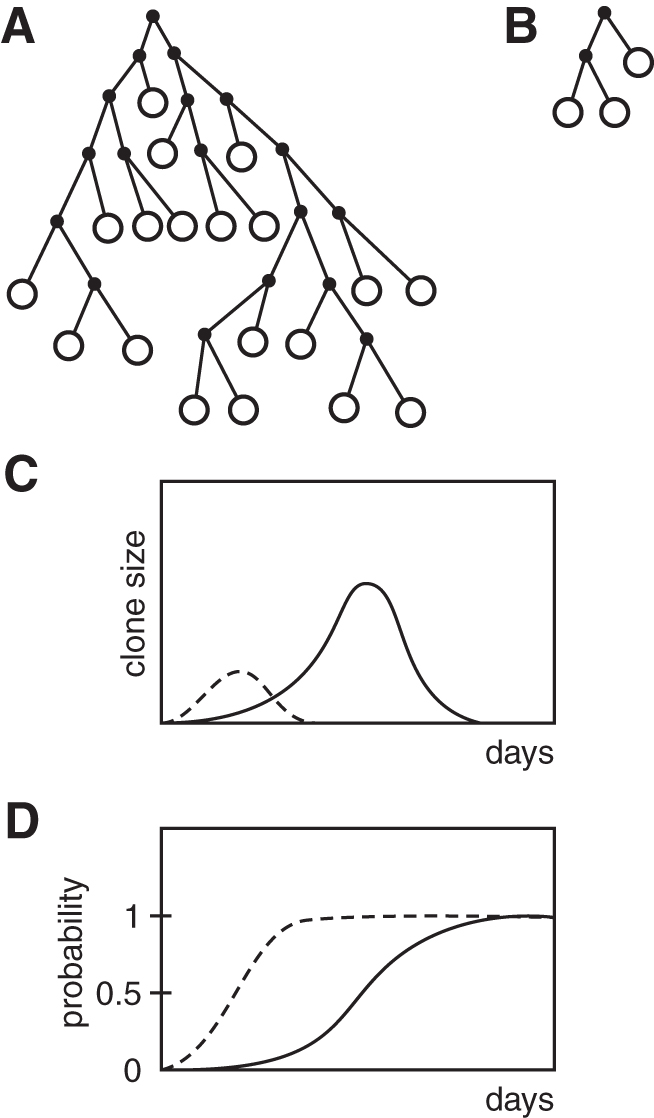
Probability model of the vegetative pool. The p-value stands for any functional property that drives the proliferation; as long as the p-value is lower than 0.5, the clone grows. (
Additional historical experiments indicated the following:
The expressed repertoire for B cells of a healthy mouse was predicted to be 107, and it was considered to be complete, which meant that it could recognize any shape of an epitope with a reasonably good affinity, and all required specificities were available in the organism before tolerance and other forms of repertoire molding. As soon as antigen appeared in the blood stream, it was thought to be carried through the lymph nodes containing some 107 to 108 lymphocytes, from which 10–100 cells might be antigen-specific. This “narrow bottle neck” made the contact with antigen-specific cells almost unavoidable. Antigen was secured on the B cell surface by the combined affinity and avidity of membrane antibodies.
A specific antibody response was shown to be elicited upon cooperation of two types of cells—B cells and T helper cells. One function of B cells was to produce antibody, while one function of T helper cells was to bind carrier and support B cell activation. For both B cells and T cells, initial activation was by a classical Bretscher and Cohn's two-signal event (5). A first signal resulted from the binding of antigen to the Ig receptor of a B cell, while a second signal involved acceptance of short-range cytokine signals from helper T cells, or the signals induced by interactions of membrane molecules (e.g., CD40 on B cells with CD40L on T cells).
Three sets of molecules (Ig, TcR, and MHC) were considered responsible for the specificity of immune responses by virtue of their capacity to bind foreign antigen. All three sets of molecules belonged to the immunoglobulin superfamily, with the most notable feature of homologous domains of ∼100 amino acids that were usually encoded by a single exon and characteristically had a single intradomain disulfide bond.
Antigen presentation was shown to be a rather complex issue. There were at least three principal kinds of antigen presenting cells, dendritic cells, macrophages, and B cells; the former two were involved in antigen presentation leading to T helper cell activation, while the latter ones (B cells) had a dual function of antigen presentation and acceptance of T helper signals. Antigen-specific B cells were highly efficient at presenting the antigen, because the Ig receptor acted as a “concentrating device.” When antigen was present at very low concentrations, dendritic cells or macrophages might fail to acquire amounts of antigen needed for further action, while epitope-specific B cells performed the task.
Repeated entry of antigen into an organism was met by a more effective immune system than at the first encounter. Shortly after the primary immune response took place clonally amplified memory B cells and memory T cells could be detected in the organism. B cells and Th cells were thought to be expanded 10- to 100-fold. For maintaining memory for a prolonged time period, it was assumed that (a) presence of antigen was required, and (b) repetitive restimulation of cells with the antigen trapped in the follicular dendritic cells was required (this could be achieved also by the presence of cross-reactive antigens). Important in the maintenance of memory cells was the trapping of antigen-antibody complexes at the site of homing of memory cells. In the absence of an activation signal, it was thought that the memory might be abolished. These ideas have been debated and modified for decades (16,17,23,36,84, and 87).
For establishment of memory, it was suggested that IgD might play a role (70). The actual memory B cells could produce IgM or antibodies of a switched isotype (IgG, IgA, or IgE). Since a prolonged response led to affinity maturation, the resulting higher affinity of the memory B cells enabled cells to respond to a lower concentration of antigen and to activate helper CD4 T cells more easily. Considering both memory B cells and T cells, the alacrity rule might direct the memory cells to settle at future sites of infection, which would extend protective immunity from a local phenomenon to a systemic one.
Specific tolerance was observed when reactive clones were anergized, deleted, or otherwise incapacitated, while specific immunity developed when clones were induced to proliferate and differentiate into effector cells. Clearance of superfluous cells at the end of infection was considered as important as the clonal expansion of the responding cells, requiring either active cell suppression and/or removal of an activation signal.
Cell walls of various microbes were known to contain polysaccharides, and such antigens actively stimulated B cells “directly,” inducing strong IgM responses (89). It appeared that the switch to other isotypes was rare, although exceptions were noted, such as the switch from IgM to IgG2 in response to pneumococcal polysaccharides (82).
B cells could respond directly to an antigen if that antigen was able to cross-link the receptors on the B cell surface. As examples, DNA, polysaccharides, and glycolipids were found to induce a T cell independent response. Such responses did not result in memory typical of T-dependent responses [the induction of T cell-independent B cell memory has since been better characterized (13)]. Whether thymus-independent antigens—upon binding to the Ig receptors of the specific B cells—were endocytosed or not was considered of lesser relevance to this type of response, since antigens were not specifically associated with Major Histocompatibility Complex (MHC) molecules, and thus were not specifically recognized by helper T cells.
Superantigens were identified as polypeptide components of many pathogens (32,41,42,81, and 86). Instead of binding in the groove of the MHC molecule, they bound to the outside structure of the MHC molecule and stimulated a much larger portion of T cells than did conventional peptide antigens, up to 2–10% of all T cells of the organism. This stimulation often did not prime the immune response. On the contrary, it induced a massive production of various cytokines with the consequence of causing systemic toxicity and immune response suppression toward bacterial antigens (9). One well-known superantigen was staphylococcal endotoxin. Besides bacterial superantigens, viral superantigens were also discovered (80).
Since the stimulation of T cells by superantigens involved a unique cascade of events, a classical memory population was not established and a new pathogenic encounter could resemble a primary infection, or in some cases could exhibit immune tolerance (86).
Upon infection, the number of lymphocytes increased as a rule dramatically. The mitogenic components of the pathogen induced a nonantigen-dependent proliferation of both T and B cells, and the clonal response increased the pool of specific cells. The clonal component appeared to contribute only insignificantly to the overall increase of cell numbers when responses initiated. Nevertheless, a repeated infection might expand the pool of cells specific to that response considerably. Increased antibody concentration upon the expansion of the responding clones meant also an increased concentration of the idiotopes displayed on the newly formed antibody molecules. These idiotopic elements provoked an anti-idiotypic response. Several waves of anti-idiotypic responses as predicted by the “idiotypic network hypothesis” (29) might include a profound perturbation of the cellular balance of the immune system.
Results suggested the following: the immune response occurred in a certain milieu (in the animal organism or under in vitro conditions). Since the cells differed, those which were more compatibly equipped would engage in the response; others “stayed behind.” The environment in which the response occurred determined which specific cells would be activated to respond. The ambiance provided an immense array of nonspecific molecular components that were perceived by some cells as “helpful” and by others as “uninviting.” Accordingly, some cells responded and others remained idle.
Summary
Historical experiments suggested the following: the B cell was a lymphocyte that usually produced antibody of a single specificity, potentially modified by somatic mutations. Upon adequate stimulation, the activated cell underwent a blast transformation and initiated clonal proliferation. Depending on signals, it formed a small or a large clone. It formed what we have called a vegetative pool, proliferating cells that ultimately converted into AFCs. Depending on the conditions governing the response, the vegetative pool increased (at the onset of the response) and then it became exhausted (toward the terminal stage of the response). In certain in vitro culture conditions, cells leaving the vegetative pool became AFCs, producing some 2,000–10,000 antibody molecules/sec/cell, and they did so up to 7 days and then they decayed. Plasma cells that could survive weeks, months, or years were not recognized at the time (20,21,23,36,84, and 87).
It was predicted that the population of B cells retrieved from an intact animal was greatly heterogenous, with a repertoire of some 107 specificities in a mouse, capable of binding 108 or more epitopes. The frequency of cells specific for many epitopes used in experimental immunology was about 1/100,000. The repertoire was proposed to be complete, thus it was capable of recognizing any antigenic determinant from the outside world (at least before tolerization or other types of repertoire molding). The recognition occurred with various degrees of affinity or avidity. The heterogeneity of cells related not only to millions of monospecific cells but was also reflected by the heterogeneity of the molecular inventory of cells.
For the induction of the antibody response, various signals were required; nevertheless, some cells—depending on the milieu of signaling—were more eager to respond than others. Cells that remained idle in a given milieu might vigorously respond under another condition. This behavior was covered by the notion of alacrity.
Small lymphocytes were found to have a minimal amount of cytoplasm with a small number of ribosomes, while blast cells possessed a full-blown functional protein-synthetic machinery, with all components needed for proliferation, maintenance, and repair. A blast cell was a “perfect” member of the clonal vegetative pool, while the plasma cell was considered an end cell, converted to a factory for antibody secretion, with lesser maintenance and functional repair. Antibody was produced at a high rate, and the membrane surface increased as the antibody was exocytosed.
The signaling events as well as the milieu in which the cells dwelled dictated whether the response would take place, whether the clone decayed or memory cells were formed. The responsiveness of both primary cells and of memory cells was closely associated with the alacrity of the cell population. The precise milieu at different ports of antigen entry (to the organism) might influence the alacrity choice of specific cells, dictating which cells would respond and which would not.
It is still controversial to what extent the immune system of the organism comprises B cells capable of producing antibody molecules against self components and why they remain idle under nonautoimmune conditions.
The historical discoveries described here set the stage in the immunology field. The alacrity model might evolve into a comprehensive theory of cellular responses. It takes into consideration the entire expression profile of the cells in the complex environment of the participating cell populations. Factors that modify B cell proliferation, activation, and survival were first revealed by outcome, prompting targeted research of factors, signals, and signal transduction mechanisms. Partner articles in this issue will describe recent developments and further explanations for B cell alacrity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.