
Abstract
Adaptive antibody responses provide a crucial means of host defense against viral infections by mediating the neutralization and killing infectious pathogens. At the forefront of humoral defense against viruses lie a subset of innate-like serum antibodies known as natural antibodies (NAbs). NAbs serve multifaceted functions in host defense and play an essential role in early immune responses against viruses. However, there remain many unanswered questions with regard to both the breadth of viral antigens recognized by NAbs, and how B cell ontology and individual antigenic histories intersect to control the development and function of antiviral human NAbs. In the following article we briefly review the current understanding of the functions and source of NAbs in the immune repertoire, their role during antiviral immune responses, the factors influencing the maturation of the NAb repertoire, and finally, the gaps and future research needed to advance our understanding of innate-like B cell biology for the purpose of harnessing NAbs for host defense against viral infections.
What Are Natural Antibodies?
The term “natural antibody” broadly encompasses antibody reactivity present in sera, in the absence of infection or deliberate immunization. The evolutionary conservation of natural antibodies (NAbs) in virtually all vertebrate species strongly suggests they play a significant role in the host immune system. Immunoglobulin M (IgM) antibodies arise first during B cell ontogeny and constitute a significant portion of the natural immunoglobulin repertoire. In adults, NAbs are maintained at relatively stable concentrations, and broaden to encompass antibodies of multiple immunoglobulin isotypes, including IgG and IgA antibodies (67). Natural human serum Ig contains a diverse repertoire of antibodies that bind to distinct carbohydrate and phospholipid epitopes, which occur as constituents of diverse classes of macromolecules including antigens of viral, bacterial, and fungal pathogen origin and self-antigens (42,68).
NAbs serve multiple primary functions within the host immune system; first, they provide an important first-line defense against virulent infectious microorganisms, including viral, bacterial, and fungal pathogens (11,21,26,61,103,110,124). The role of NAbs in host antiviral immunity will be discussed in greater detail later in this article. Second, NAbs serve “housekeeping” functions through the recognition of neo-self-epitopes generated during various cellular processes. NAbs promote the clearance of self-antigens generated during natural cellular senescence, which otherwise potentiate autoimmune disease, and modulate the activation of phagocytes after engulfment of IgM-opsonized apoptotic cells (59,130). NAbs have similarly been shown to modulate immune responsiveness to products of host molecule oxidation (19,36) and allergenic particles (77,107,108), and additionally exhibit tumor surveillance functions through the recognition of altered self-oncoantigens (25,85,133). Finally, NAbs play roles in the development and selection of the B cell repertoire, and the regulation of antigen-specific B cell responses (20,46,99,100).
Ontogeny of NAb-Producing B Cells in Mice
B cell subsets in mice are anatomically compartmentalized and exhibit distinct functions. The production of NAbs is associated with innate-like subsets of B cells, including the B-1 and, to a lesser extent, marginal zone (MZ) B cells (15,35,74,91). B-1 B and MZ B cells express Ig receptors that react with highly conserved antigens and mediate innate-like B cell effector functions, in contrast to B-2 B cells (also known as follicular B cells), which participate in germinal center-dependent adaptive immune responses (15). Mouse B-1 B cells express a CD19+B220LowCD5+/−CD43+CD11b+/− phenotype and can be further differentiated into CD5+ B-1a B cells and CD5− B-1b B cells (72). B-1 B cells are enriched in the peritoneal and pleural cavities, and the omentum, a site of fetal B cell lymphopoiesis and a CXCL13-dependent point of entry for B-1 B cells to the peritoneal cavity (PerC) (5,119,121). The low frequencies of B-1 B cells in spleen and lymph nodes, suggest that they also circulate through secondary lymphoid organs (72). In addition to producing NAbs, B-1 B cells have been shown to contribute to the generation of adaptive antibody responses to T-independent antigens during infection with a variety of pathogens, including influenza virus (11,34) and multiple bacterial species (3,4,26,53,61,141,142). They also contribute to secretory IgA in response to the commensal microbiota (28,79,84). MZ B cells, that express a CD19+B220+CD21HiCD23LowCD9+CD36+ phenotype, populate the MZ at the border between lymphoid follicles and red pulp of the spleen, and provide surveillance and rapid defense against blood-borne particulate antigens (90,92,136,137).
In mice innate-like B cell subsets are the product of specialized developmental programs that enrich for autoreactive B cell receptor clonotypes (72). B-1 B cell precursors arise predominantly during fetal and neonatal life from distinct progenitors (52,73,96). Their development is dependent on the induction of the transcription factor Arid3a (146), which is reciprocally regulated by the Lin28b/Let-7 axis that governs fetal versus adult lymphopoietic programs (145). MZ B cells are derived from adult bone marrow B cell progenitors and depend on signaling through Notch2 after engagement of the ligand delta-like 1, expressed by stromal cells of the splenic MZ (115,128). The selection of innate-like B cell precursors into both the mature B-1 and MZ B cell compartments requires B cell receptor signaling-dependent selection (15,64). Innate-like mouse B cells formed during fetal and perinatal development exhibit preferential utilization of IGHD-proximal IGHV variable-region regions (87,109,139,140), and minimal junctional diversity because of reduced terminal deoxynucleotidyl transferase (TdT) activity during these periods (16). These developmental constraints are paramount to the establishment of stereotyped B cell clonotypes that comprise a large proportion of the NAb-producing B cell repertoire, as ectopic expression of TdT transgenes was shown to disrupt the programmed development of highly conserved TEPC15+ B cell clonotypes, which normally protect mice from infection with Streptococcus pneumoniae through their recognition of phosphorylcholine (PC)-containing cell wall antigens (17,89). The unique developmental pathway of innate-like B cells is therefore central to their role in the immune system.
Innate-Like B Cells in Host Defense
NAbs preexisting in serum represent an important first line of defense that can neutralize and, in some cases, directly kill invading pathogens. Although the nature of NAb-producing cells is subject to debate, many studies have demonstrated that B-1 B cells are a prominent source of NAb in mice (11,80). Within the B-1 B cell compartment, both B-1a and B-1b B cells have been shown to contribute to the secretion of NAb (34,35,53,61,66,105). MZ B cells (69,91) and non-B-1 fetal precursors (111) have additionally been shown to contribute to natural IgM production. PerC-localized B-1 B cells, which do not spontaneously secrete antibody in situ, are precursors to NAb-producing cells that populate the spleen and bone marrow of mice, suggesting that some degree of differentiation is necessary to initiate NAb secretion by innate-like B cells (74,106). In fact, two pathways through which innate-like B cells produce NAb have been described. The first parallels the classical differentiation of B cells into plasma cells, and relies on Blimp1 to promote the acquisition of plasma cell phenotypes, characterized by reduced expression of CD19 and expression of CD138 (111,116) (Fig. 1A). Of interest, NAb-producing plasma cells in bone marrow have been found to exhibit a dependency on interleukin-5 (IL-5), like B-1 B cell-derived IgA+ plasma cells that populate the intestinal mucosa (9,97), distinguishing them from B-2 B cell-derived IgG-producing plasma cells that are dependent on IL-6 and other niche-derived factors (111). B-1 B cells have additionally been shown to contribute to NAb production as nonterminally differentiated antibody-secreting cells (ASCs), through a Blimp1-independent pathway that does not result in expression of CD138 or the downregulation of surface immunoglobulin (35,116) (Fig. 1A). Although the molecular signature of B-1 B cells remains to be fully characterized, the enrichment of natural IgG3 ASCs in this compartment suggests it will be interesting to learn how these pathways contribute differentially to host immunity (116).
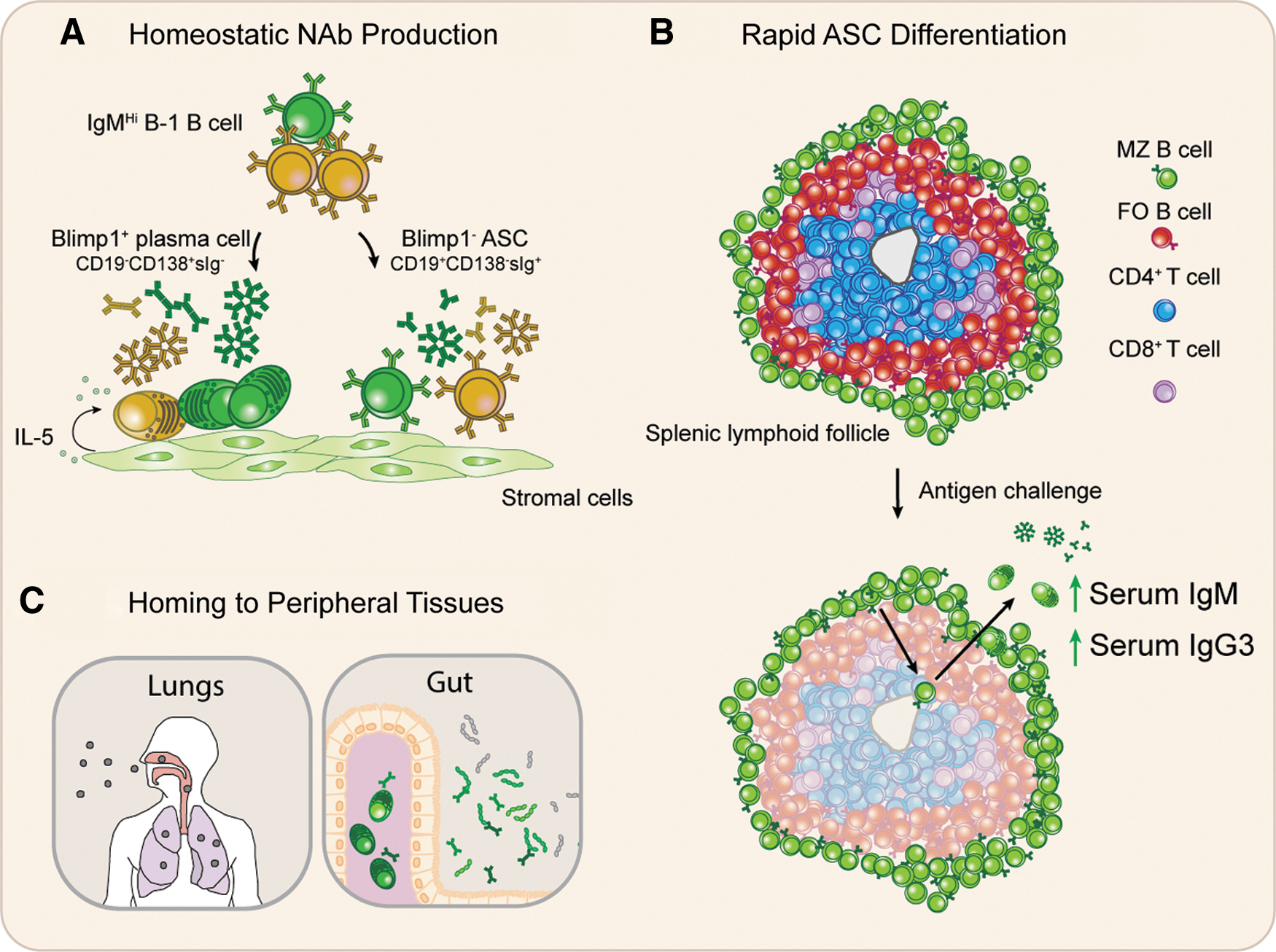
Immune functions of innate-like B cells. Innate-like B cells play a pivotal, multifaceted role in host defense against infection by bacterial and viral pathogens.
Both B-1 B cells and MZ B cells are additionally poised for rapid mobilization and secretion of antigen-specific antibodies, which provides an additional fast-acting defense against pathogens (90,92). B-1 B cells rapidly differentiate into ASCs after stimulation by molecules such as lipopolysaccharide and peptidoglycan, which is accompanied by limited clonal proliferation (143). The Toll-like receptor 7 agonist Resiquimod (R848), which bears structural similarities to viral nucleic acids, has also been shown induce proliferation and Ig secretion by mouse B-1 B cells, suggesting B-1 cells are additionally poised to mount rapid humoral responses after the encounter of viral antigens (51,125). Both B-1a and B-1b B cells have been shown to contribute to antigen-specific immunity in mice (4,61,141). Similarly, challenge with bacterial glycolipid antigens drives rapid migration of antigen-specific MZ B cells to splenic follicles, which subsequently translocate to the red pulp as plasma cells producing antigen-specific IgM and IgG3 antibodies (92) (Fig. 1B). Although NAbs and extrafollicular immune response-derived antibodies exhibit lower affinity for their antigens than GC-derived antibody and in some cases are polyreactive, they provide an essential rapid protection during the time required to form later appearing GC-dependent adaptive immune responses (98).
B-1 B cells in mice further contribute to host defense by localizing to sites of infection and producing antibody locally (Fig. 1C). In mouse models of viral pneumonia associated with influenza, infection-induced type 1 interferon mobilizes B-1 B cell migration to lung-draining mediastinal lymph nodes through the activation of CD11b integrin, which is expressed by a subset of mouse B-1 B cells (134). B-1 B cells secreting NAb have also been reported in the lung parenchyma at steady state (34). Innate-like B cell subsets in mice therefore serve to limit the dissemination of infectious pathogens through their expression of templated B cell receptors reactive with conserved antigens, the steady-state secretion of NAb, and their capacity to mobilize during antigen challenge to produce rapid increases in antigen-specific serum Ab.
NAbs in the Human Immune System?
The existence of a human equivalent of mouse B-1 B cells is somewhat controversial. In contrast to CD5 and CD43 expression by mouse B cells, which effectively differentiates B-1 B cells from other B cell subsets, these markers are modulated by cellular activation in human B cells (131). Furthermore, whereas the unique anatomical distribution of mouse B-1 B cells has aided in their characterization, the analysis of resident cells of the peritoneal and pleural cavities is not easily accomplished in humans. However, like mouse B-1 B cells, human fetal B cells are enriched for clonotypes exhibiting polyreactivity for autoantigens (33,83,95), and fetal human B cells additionally exhibit lower levels of TdT-dependent junctional diversity and a preferential utilization of specific VH genes (95,113). The human omentum is also a site of fetal B cell lymphopoiesis (119,120); thus, there are several conserved features of fetal B cell lymphopoiesis between humans and mice. CD20+CD27+CD43+CD70− B cells in umbilical cord blood and adult peripheral blood bear similarities to mouse B-1 B cells, including the spontaneous secretion of antibody and the expression of largely unmutated immunoglobulin genes enriched for reactivity to common NAb targets, such as PC (56,58,114). However, the classification of these cells as human B-1 B cells has been contested (37,39,57). Although humans possess a MZ-like B cell population, characterized by CD27-expressing IgM+ memory B cell phenotype, human MZ B cells are more widely distributed than those of mice, and it is not clear to what extent they contribute to homeostatic NAb production (32,135). Thus, the precise nature of NAb-producing B cells in humans remains unclear, and to date it is unknown whether NAbs in humans share a similar ontogenetic program as those in mice.
It is clear, however, that humans possess serum antibodies functionally analogous to mouse NAbs. Although rare in humans, selective IgM deficiency commonly presents in the clinic with repeated chronic infections by bacterial, viral, fungal, and protozoan pathogens (60,65,71), mirroring the essential role of NAbs and induced IgM in host defense of mice. IgM-deficient patients additionally exhibit high frequencies of allergic disease (81), and there are reported cases of IgM deficiency co-associating with autoimmune disease including rheumatoid arthritis (8), systemic lupus erythematosus (126), glomerulonephritis (6), and myriad of other human autoimmune diseases (81). These findings strongly suggest that certain human antibodies perform “housekeeping” functions, similar to those described for mouse NAbs. Moreover, serum concentrations of specific antibodies have been shown to be predictive of disease progression in several settings, including: (i) oxidized low-density lipoprotein and advanced glycation end product modification-reactive Abs in cardiovascular disease (43,47), (ii) laminaribiose (Glcβ1–3Glcβ)- and chitobiose (GlcNAcβ1–4GlcNAcβ)-reactive Abs in Crohn's disease (40) and (iii) Glcα1–4Glcα-specific antibodies in relapsing–remitting multiple sclerosis (117).
Antibodies reactive with a multitude of phospholipid and carbohydrate epitopes are abundant in the human serum Ig repertoire (14,42,68). Encompassed in the human serum NAb repertoire are antibodies reactive with common antigen targets of mouse NAbs, such as PC and N-acetyl-
NAbs in Antiviral Immunity
The impact of experimental conventions used during the study of host immune responses on our ability to observe the contributions of NAbs in antiviral immunity was succinctly described by Ochsenbein and Zinkernagel: “In many virological and immunological assays it is a convenient practice to predilute sera to variable extents, from 1:10 up to 1:80, to avoid so called ‘nonspecific background’ signals; the consequence of this practice is that naturally occurring antibodies are not normally detected or are not registered.” (103)
Thus, serum is often diluted in immunological assays beyond the functional range of NAbs and their reactivity for viral antigens, or neutralizing activity against infectious viruses are therefore imperceptible and have remained largely unstudied.
However, studies specifically examining NAbs have found significant evidence for their early and critical role in preventing the tissue dissemination of viruses. In fact, mice deficient in Ig production, as a result of genetic ablation of recombination activation genes (RAG) or the IgM heavy chain (μMT) exhibit viral titers 10–100 times greater in peripheral tissues than wild-type mice and show increased rates of morbidity after viral challenge (103) (Fig. 2). Similarly, mice deficient exclusively in secretory IgM exhibit impaired clearance of influenza virus, indicating that B-1 B cell-derived natural IgM serves an important role in host defense against viruses (12). Low titers of IgM NAbs that recognize vesicular stomatitis virus (VSV) protect against VSV-induced encephalitis and have long been appreciated as components of the NAb repertoire in mice (54,103), and NAbs have also been shown to be an important defense against infectious influenza virus (11,34,70). The transfer of natural serum antibodies to antibody-deficient mice is sufficient to delay mortality after challenge with infectious VSV or influenza virus, indicating the significance of their early role during antiviral immunity (10,103). Like mice, humans with selective IgM deficiency are at an increased risk of viral infection, and often present with infections caused by varicella zoster virus, cytomegalovirus, and molluscum contagiosum virus (81). Similarly, humans have been shown to possess low titers of natural IgM antibodies that recognize influenza virus (13). Thus, as is the case in mice, IgM represents an important front-line of defense against infectious viruses in humans.
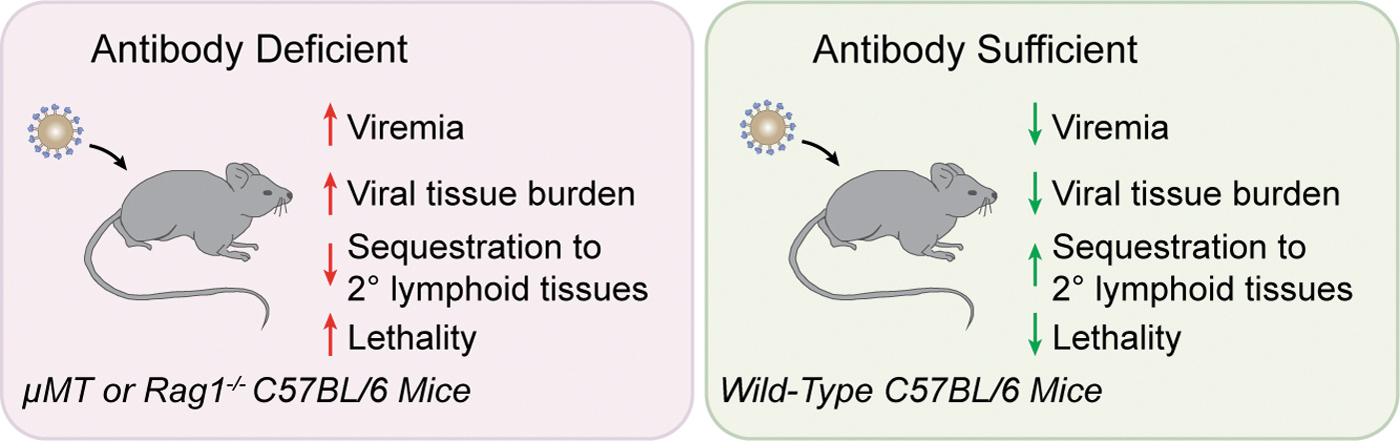
Phenotype of NAb-deficient mice after viral challenge. NAbs play an essential role in the early defense against viral infection in mammalian hosts. When challenged with a low virus dose, mice lacking B cells and deficient in normal serum antibodies exhibit increased mortality (103). Viral burden in blood and tissue is high in these animals, relative to their antibody-sufficient counterparts, and the sequestration of viral particles to secondary lymphoid tissues is reduced.
In vitro neutralization of viral particles mediated by mouse natural IgM has been reported for lymphocytic choriomeningitis virus (LCMV) (103), vaccinia virus (54), and influenza (70), and the mechanisms underlying NAb-mediated virus neutralization appear heterogenous. Passive NAb transfer to antibody-deficient mice does not require intact complement activity to neutralize VSV, as depletion of complement components by treatment with cobra venom factor did not significantly impact the efficacy of transferred protection (103). It is possible that NAbs neutralize VSV directly by interfering with its invasion of host cells (Fig. 3A). Conversely, in the case of influenza infection, NAb-dependent activation of complement pathways was required for effective neutralization of virus (70). Consistent with the notion that viruses are considered to be resistant to complement-mediated lysis, the neutralization of influenza virus was instead dependent on robust deposition of C3b and occlusion of hemagglutinin epitopes, necessary for recognition and fusion with host cells (Fig. 3A) (70). Human influenza-reactive NAbs were similarly shown to neutralize virus in vitro through complement-dependent means (13). Similarly, antibody-mediated deposition of C4 was recently shown to neutralize human adenovirus 5 (Ad5), a model organism for nonenveloped viruses, by blocking entry to the cell cytosol through inhibition of the capsid disassembly process in a C3 independent pathway (23). Thus, NAbs reactive with viral antigens serve multifaceted roles with respect to co-opting the classical complement pathway to neutralize infectious viruses.
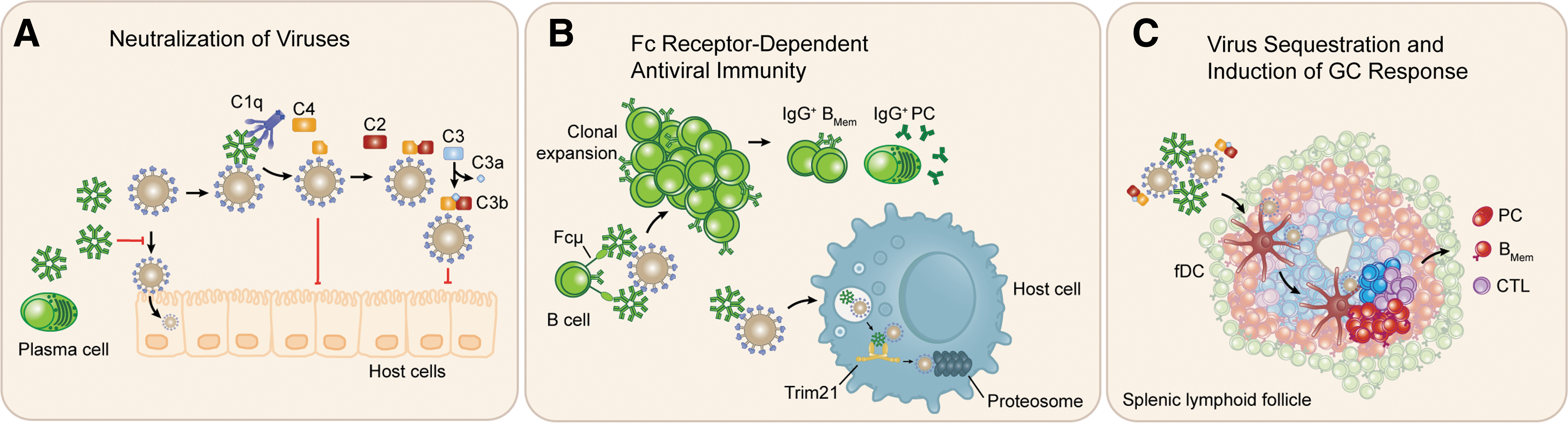
Modes of NAb function against viral infection. NAbs function in several modes contributing to host defense against viruses.
NAbs may further promote protection from viral infections through interactions with receptors for the fragment crystallizable (Fc) region of immunoglobulin molecules. With respect to natural IgM, two such Fc receptors have been directly implicated in antiviral immune responses. Trim21, an intracellular receptor for Ig Fc regions with high affinity for IgG molecules has also been shown to bind to Fc region of IgM molecules and neutralize infection by the adenovirus (86) (Fig. 3B). Trim21 was also shown to work synergistically with antibody-dependent C4 deposition to mediate robust neutralization of adenovirus (23). The IgM-specific Fcμ receptor has also been shown to play vital B cell intrinsic roles in the early response to viral infection. Nguyen et al. demonstrated that conditional deletion of Fcμ receptor in mouse B cells results in impaired clonal expansion of hemagglutinin-reactive B cells during the early phases of infection, and reduced numbers hemagglutinin-reactive IgG plasma cells and memory B cells (101) (Fig. 3B). Thus, in addition to mediating complement-dependent neutralization of infectious viruses, natural IgM seems to play important roles in Fc receptor interactions that promote virus neutralization and the generation of high-affinity antibody memory B cells.
Perhaps the most significant outcome for NAb-mediated opsonization of viruses is to sequester them to secondary lymphoid tissues (93,103,104). This occurs, in part, through the localization of antigen–antibody complexes to follicular dendritic cell networks in lymphoid follicles through interactions with CD35 (complement receptor 1) and CD21 (complement receptor 2), which promotes induction of germinal center-dependent adaptive immune responses to viral antigens (112) (Fig. 3C). This effect has been observed in mouse models of infection by influenza and VSV, wherein NAbs lower infectious influenza virus titers, and impede tissue dissemination (10,103). These features of NAb activity seem to be essential for the development of class-switched antiviral antibodies, effective T lymphocyte activation, and the generation of long-term immunological memory to influenza virus (12,44,78)
NAb Responses to Viral Envelope Glycoproteins
Viruses often utilize host glycosylation machinery during viral replication and assembly in host cells. NAb reactivity and neutralization potential against viral pathogens may therefore be closely associated with the host species from which a virus originates. Because of this feature of viral replication, certain self-reactive NAbs can provide protection against chronic viral infections. This effect is exemplified by the family of human broadly neutralizing HIV antibodies that react with high-mannose modifications of gp120 viral envelope glycoprotein, such as the monoclonal antibody 2G12. 2G12 potently binds and neutralizes primary isolates of HIV and SIV through recognition of a conserved Manα1–2Man motif on the oligomannose glycan (29,82). High-mannose-binding antibodies that recognize the HIV glycoproteins exhibit features of autoreactive antibodies, perhaps because of the derivation of viral antigenic epitopes from host carbohydrates. Of interest, 2G12 also binds mannose-containing cell walls of some Candida species, suggesting the HIV-neutralizing potential of the human NAb repertoire may be influenced by exposure to pathogens bearing conserved mannose structures (41). Self-reactivity has been reported in other broadly neutralizing HIV human monoclonal antibodies, such as MAbs 2F5 and 4E10, which show reactivity to the widely expressed lipid cardiolipin (1), and broadly neutralizing antibodies against stem epitopes of influenza hemagglutinin have also been reported to exhibit autoreactivity (7). The autoreactivity of these antibodies have given rise to speculation that the NAb-producing B cell repertoire could be leveraged through vaccine design to elicit broadly neutralizing antibodies.
NAb-mediated viral neutralization through recognition of viral envelope glycoproteins may be especially important in protection against zoonotic viral infections. For example, humans and old-world primates, which do not generate endogenous αGal1–3αGal glycan modifications because of the evolutionary loss of the α-galactosyl transferase (αGalT) enzyme, generate high titers of αGal-reactive serum antibodies. Natural anti-αGal antibodies effectively opsonize virus containing αGal-modified capsid antigens through antibody-mediated classical complement cascade activation (127). αGal-modified capsid antigens are common to animal viruses, including rhabdoviruses, rabies virus, some nonprimate lentiviruses, and foamy viruses, indicating the breadth of protection potentially provided by this NAb defense (127). Similarly, when considering infection by arthropod-borne arboviruses such as dengue flavivirus, which originate from insect cells and express insect host cell-derived glycans, insect glycan-reactive NAbs may represent an important front-line defense, before the virus's acquisition of host glycosylation during replication within mammalian host. Natural anti-αGal antibodies provide immediate protection in mouse models of insect-borne malaria transmission, and can be boosted by vaccination with αGal conjugates to provide sterilizing immunity in αGalT-deficient mice (144). Of interest, similar effects have been reported during transmission of hepatitis C virus (HCV), wherein viral capsid glycoproteins are modulated by host ABO blood group oligosaccharide alloantigens and some degree of immunity to HCV can be demonstrated in allotype mismatched infections (132). Although the presence of blood group alloantigen-reactive NAbs in the human immune repertoire remains enigmatic, perhaps they represent a barrier to infection against viruses bearing alloantigen-derived carbohydrate epitopes.
NAb recognition of viral envelope glycoprotein modifications hold implications beyond host defense and can significantly impact the efficacy of therapeutic viral vectors. Specifically, the galactosylation of VSV G glycoprotein, after production of VSV in αGal-positive cell lines, resulted in failed gene delivery to human cells because of anti-αGal antibody-dependent, complement-mediated inactivation and blockade of viral particle replication (50). Conversely, during treatment with PROSTVAC-VF, a poxvirus-based therapeutic cancer vaccine now in phase III clinical trials for the treatment of advanced prostate cancer, preexisting serum NAbs to blood group A trisaccharide (BG-ATri), and antibodies reactive with the Forssman disaccharide (GalNAcα1–3GalNAcβ) induced during treatment correlated with enhanced antitumor immune responses in recipients (30,31). αGal modification of tumor vaccine antigens has also been used to improve antitumor responses in mouse models of pancreatic cancer (38), and has been shown to enhance phagocytosis and cross-presentation of tumor antigens by human antigen-presenting cells in the presence of purified anti-αGal human antibodies (88). Thus, NAb responses to viral glycoproteins can significantly impact the successful delivery of viral vector-based gene therapies and tumor antigen vaccines.
Origins of the NAb Repertoire
As discussed previously, the recruitment of B cell progenitors into NAb-producing B cell subsets occurs by positive selection through B cell receptor signaling. During fetal B cell lymphopoiesis, this is thought to occur predominantly in the context of self-antigens. Unlike class-switched antibody isotypes, serum concentration of IgM antibody is not affected in germ-free mice, and some classes of NAbs are found at comparable levels in germ-free and conventionally raised mice (36,63,103). However, microarray analysis of global serum antibody reactivity in germ-free mice revealed significant deficiencies in carbohydrate-reactive antibodies commonly observed in conventionally raised mice (22). Germ-free (GF) mice also exhibit blunted antibody T-independent antibody responses to α1–3 glucan carbohydrate antigens after immunization with Enterobacter cloacae, indicating that microbial colonization can alter the clonal response to polysaccharide vaccine antigens (75). Moreover, it is clear that neonatal exposure to bacterial antigens can result in striking and long-lasting changes to the clonal composition of antigen-specific NAbs. Neonatal immunization of mice with S. pneumoniae and E. cloacae results in the expansion of B cells expressing immunoglobulin idiotype determinants distinct from those that dominate the resting repertoire of unimmunized adult mice (76). Thus, the clonal repertoire of NAb-producing innate-like B cells may be shaped significantly by the early-life antigenic history of an individual that, by extension, may impact their susceptibility to infectious, autoimmune, or allergic disease.
Human antibodies with specificities similar to mouse NAbs also bear signatures of environmental antigen modulation. Although found at low levels in umbilical cord blood, antibodies reactive with carbohydrate and phospholipid epitopes increase significantly with age (62,76,138), and serum concentrations of NAbs directed toward PC and GlcNAc antigens are clearly modulated by infection with S. pneumoniae and Streptococcus pyogenes (Group A Streptococcus), respectively (55,118). Unlike the NAb-producing B cell clonotypes of mice, which express predominantly germline Ig gene rearrangements, PC-reactive human B cells exhibit significant somatic mutation of their Ig gene variable regions (45). Although the relationship of these human B cells to NAb production is unclear, these findings may support a role for progressive antigen encounter at pulmonary and gut mucosal tissues in the selection of B cells that contribute to NAb production in humans. The effects of IG gene mutations on antigen specificity, affinity, pathogen-neutralizing potential, and immune modulating activity of human NAbs are poorly characterized, but their presence suggests that early life encounters with pathogen-, commensal microorganism-, and vaccine-derived antigens may modulate the antiviral activity of NAbs discussed previously.
Heterogeneity of Fine Specificity Within the Human NAb Repertoire
The global glycan and phospholipid reactivity observed in human sera results from myriad individual antibodies with distinct, and in some cases, overlapping fine specificities. However, the NAb repertoire is often described in aggregate as a single entity, and few studies have extensively analyzed individual NAb specificities within the global repertoire. Although human sera clearly exhibit substantial reactivity toward host glycan and glycolipid structures, the fine antibody specificity of NAbs may differ broadly among individuals, and across Ig subclasses within individuals (14,42,68). Despite the well-conserved, oligoclonal nature of human antibodies directed toward PC, GlcNAc, αGal and A/B blood group antigens, individual clones comprising these antibody pools exhibit variable fine specificity: PC-reactive monoclonal antibodies derived from EBV immortalized human B cells were shown to exhibit considerable heterogeneity in terms of affinity to different synthetic derivatives of PC (123), and several studies have described differences in the fine specificity of αGal-reactive antibodies in human sera (18,102), which is clearly modulated in cases of blood group B alloantigen-positive individuals (49). Moreover, although the human NAb repertoire contains abundant mannose-reactive antibody that can bind to HIV envelope glycoproteins, the majority of these polyclonal antibodies exhibit no neutralizing activity, indicating that distinct mannose-reactive B cell clonotypes are required for effective neutralizing NAb responses (129). Thus, subtle differences in the sequence and the degree of mutations of immunoglobulin genes may produce significant differences in the capacity of antibodies to recognize defined T-independent epitopes as arrayed in different contexts, which may subsequently affect their function during the opsonization, neutralization, and lysis of infectious pathogens.
Remaining Questions
Despite the fact that NAbs have been recognized for decades as an important component of the host immune system, very little is known regarding the origin and differentiation of NAb-producing plasma cells or the regulation of NAb production in serum and mucosal secretions in humans. These shortcomings have limited the characterization of the Ig gene repertoire of human NAb-producing B cells and the exploration of the mechanisms underlying the conserved NAb reactivity to glycan glycolipids, including viral antigens. We must address these gaps in knowledge, as glycans exist at the forefront of most cellular interactions; in the glycocalyxes that decorate each cell in the body, on mucin proteins that constitute important barriers in mucosal tissues, and as surface antigens expressed by myriad pathogens where they play essential roles during infection.
The advent of high-throughput sequencing approaches to characterize immunoglobulin heavy chain gene rearrangements has provided unprecedented glimpses into the clonal diversity of the human B cell repertoire, but has also revealed convergent and conserved elements (27,122). Although past sequencing-based analyses of the human B cell repertoire have been heavily skewed toward the analysis of blood or tonsil-derived B cells, recent efforts to establish a VH gene-rearrangement-based atlas of B cell clonal distribution over multiple tissues of individual human organ donors revealed distinct networks of B cell clonal lineages that distribute across and inhabit tissues in the human body (94). The recent demonstration that lung tissue-resident B cells play an important role during secondary immune responses to influenza infection in mice (2) strongly suggest that coupling high-throughput antigen receptor sequencing efforts, such as those pioneered in the formation of the Human B cell Atlas, with the targeted analysis of antigen-specific B cells will provide unprecedented clarity of the relatedness of systemic and mucosal B cell responses to viral antigens. Extending such strategies to the analysis of B cells bearing reactivity toward known NAb antigens may provide insight into the origins of NAb-producing B cells in humans, and the distribution of these cells throughout the body. Given the role NAbs play in the context of antiviral immunity, these approaches may illuminate many of the outstanding questions outlined above and provide avenues to harness or manipulate humoral NAb immunity to improve the efficacy of virus-based vaccines and immunotherapies.
Footnotes
Acknowledgments
The authors thank Dr. Denise Kaminski for her support in editing and revising this review.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by NIH grants R01 AI14782, U01 AI100005 and U19 AI142637 (Project 1) awarded to J.F.K.