
Abstract
The importance of anti-HIV antibodies mediating antibody-dependent cell-mediated cytotoxicity (ADCC) in protective immunity against HIV is recognized recently. The purpose of this study was to measure the functional ADCC response at different stages of HIV infection in a well-defined HIV+ cohort, including 20 recently infected individuals, 30 with long-term slow-progressive, 24 with short-term slow-progressive and 32 with progressive HIV infection using a rapid fluorometric ADCC assay. The antibodies mediating ADCC were found in all disease stages. These antibodies were detectable at as early as 25 days after the estimated date of infection, however, did not influence the viral load set point probably indicating no major influence on the early course of the disease. However, the frequency and magnitude of functional ADCC responses were associated with higher CD4+T cell count and lower viral load and were significantly lower in progressors compared with other groups. The usefulness of the ADCC responses in longer viral control was assessed in a subset of participants with slowly progressing HIV infection. In these individuals, the ADCC responses observed at the visit 1 were found to be increased over time and were associated with lower plasma viral load estimated 4 to 15 years later in the disease course. Overall, the study findings confirm the role of ADCC antibodies in reducing the viral burden and also indicate the probable role of sustained functional ADCC responses in reducing the viral burden during the later period of HIV infection.
Introduction
Despite the test and treat strategy for HIV control in infected individuals, a vaccine for prevention of HIV transmission remains the ultimate goal. Although antiretroviral therapy (ART) has changed HIV infection into chronic manageable disease, the lifelong use of ART results in various side effects, adds costing, and thus adherence becomes an important issue. The moderately successful HIV-1vaccine trial RV144 has shown that anti-V1/V2 IgG antibodies mediating antibody-dependent cell-mediated cytotoxicity (ADCC) were associated with protection against HIV (15). Additionally a number of studies has shown the correlation of ADCC responses with slower disease progression (2 –4,6,12,14,21,23,27,29,33) ADCC-mediating antibodies have also shown to have a role in protection to infants against the transmission through breastfeeding (21,24).
ADCC responses are measured using different readouts. Some assays detect the activity of the effector cells such as natural killer (NK) cells in the presence of anti-HIV antibodies and the HIV antigens, whereas some assays detect lysis of the target cells coated with HIV antigens in the presence of HIV antibodies and effector cells (7,12,13,30). Various reports have shown that different readouts of ADCC assays do not necessarily correlate with each other (22). We have previously shown that the HIV-infected individuals with nonprogressive HIV infection, that is, long-term slow progressors (LTSPs) showed higher ADCC responses than progressors in NK cell activation heterologous assay (18).
The NK cell activation was measured as increased interferon-gamma (IFN-γ) secretion and CD107a expression in the presence of anti-HIV-positive serum and HIV-1peptides. This was a surrogate marker and did not necessarily measure the functionality of ADCC-mediating antibodies in terms of killing of the target cells. Hence, we assessed the functionality of ADCC responses by determining the target cell membrane integrity as a surrogate for HIV antibody-dependent killing in the infected individuals using the rapid fluorometric antibody-dependent cell-mediated cytotoxicity (RFADCC) assay. The assay measures the uptake of dye-labeled target cell (HIV-1 Env-coated CEM.NKr cells) membranes by the effector cells in the presence of plasma from HIV-infected individuals using flow cytometry readout (13).
The functional ADCC response was measured in HIV-infected individuals at various stages of infection; individuals with recent HIV infection (RHI), with short-term slow-progressive (slow progressors, SPs), long-term slow-progressive disease: LTSPs and progressive disease (Progressors). We observed that ADCC-mediating antibodies were present in almost all HIV-infected individuals with considerably lower magnitude in progressive disease. In a subset of individuals with short- and long-term slow-progressive disease, we also found that the ADCC responses detected earlier in the disease were increased over time and associated with lower plasma viral load 4 to 15 years later in the disease course.
Materials and Methods
Study subjects
Study participants were identified from individuals followed up regularly at outpatient clinics of the National AIDS Research Institute. Asymptomatic ART-naive HIV-1-infected individuals with stable CD4+T cell count (>500 cells/mm3) for more than 3 years of follow-up were identified as short-term SPs, whereas those with follow-up for more than 7 years were identified as LTSPs (26). Progressors were identified as ART-naive HIV-1-infected individuals with CD4+T cell count <500 cells/mm3. The HIV-positive individuals with less than 10 months duration between the last negative and first positive HIV test and with history of recent HIV exposure were identified as individuals with RHI (19). Twenty-four SPs, 30 LTSPs, 32 progressors, and 20 individuals with RHI were enrolled in the study along with 20 HIV-uninfected healthy individuals as controls. Twenty milliliters of whole blood sample was collected from these participants in EDTA anticoagulant after obtaining written informed consent. The plasma and peripheral blood mononuclear cells (PBMCs) were separated and stored at −80°C and −196°C, respectively, as described previously until tested (20). The study (Protocol No.: NARI/EC Protocol No.: 2012-06) was approved by the Institutional Ethics Committee. CD4+T cell counts and plasma viral load were measured as described previously (20). For the individuals with RHI, the estimated date of infection was calculated as the mid time point of the period between the last HIV-negative and first HIV-positive test (28) and the plasma viral load set point was calculated as the geometric mean titer of plasma viral load values obtained between 6 and 15 months after the estimated date of infection as described previously (19).
ADCC-mediating antibodies in the plasma of study participants
RFADCC assay
The functional ADCC activity in the plasma samples of study participants was assessed by RFADCC as described previously (10). In brief, the CEM-NKr cells (target cells) were labeled with the intracellular dye carboxyfluorescein succinimidyl ester (CFSE) and the membrane dye PKH26, pulsed with 3 μg of HIV-1C 96ZM651 gp120 recombinant protein envelope protein (Cat. No.: 10080, NIH AIDS reference reagent program) and incubated with the PBMCs from healthy HIV-negative donor (as a source of effector cells) and 1:100 diluted plasma from the study subjects for 4 h. Only target cells and target+effector cells without plasma of the study subjects were kept as controls. After incubation, the cells were stained with anti-CD14 APC and anti-CD3 PECF594 antibodies (both from BD Biosciences) for 30 min, fixed and acquired on FACSAria™ Fusion (BD Biosciences), and analyzed using FlowJo 10.0.6 software. The ADCC response was measured as the frequency of CD14+ monocytes taking up the PKH26 dye from the lysed target cells (Fig. 1a) The background response (effector+target cells only) was subtracted from the values obtained with the respective plasma samples. The response was considered to be positive when the value of % PKH26+CD3−CD14+ monocytes in the test well were above the mean response in 20 HIV-uninfected healthy controls +2SD. The mean RFADCC response for 20 healthy individuals is 0.56% and the cutoff value obtained is 4.10%.
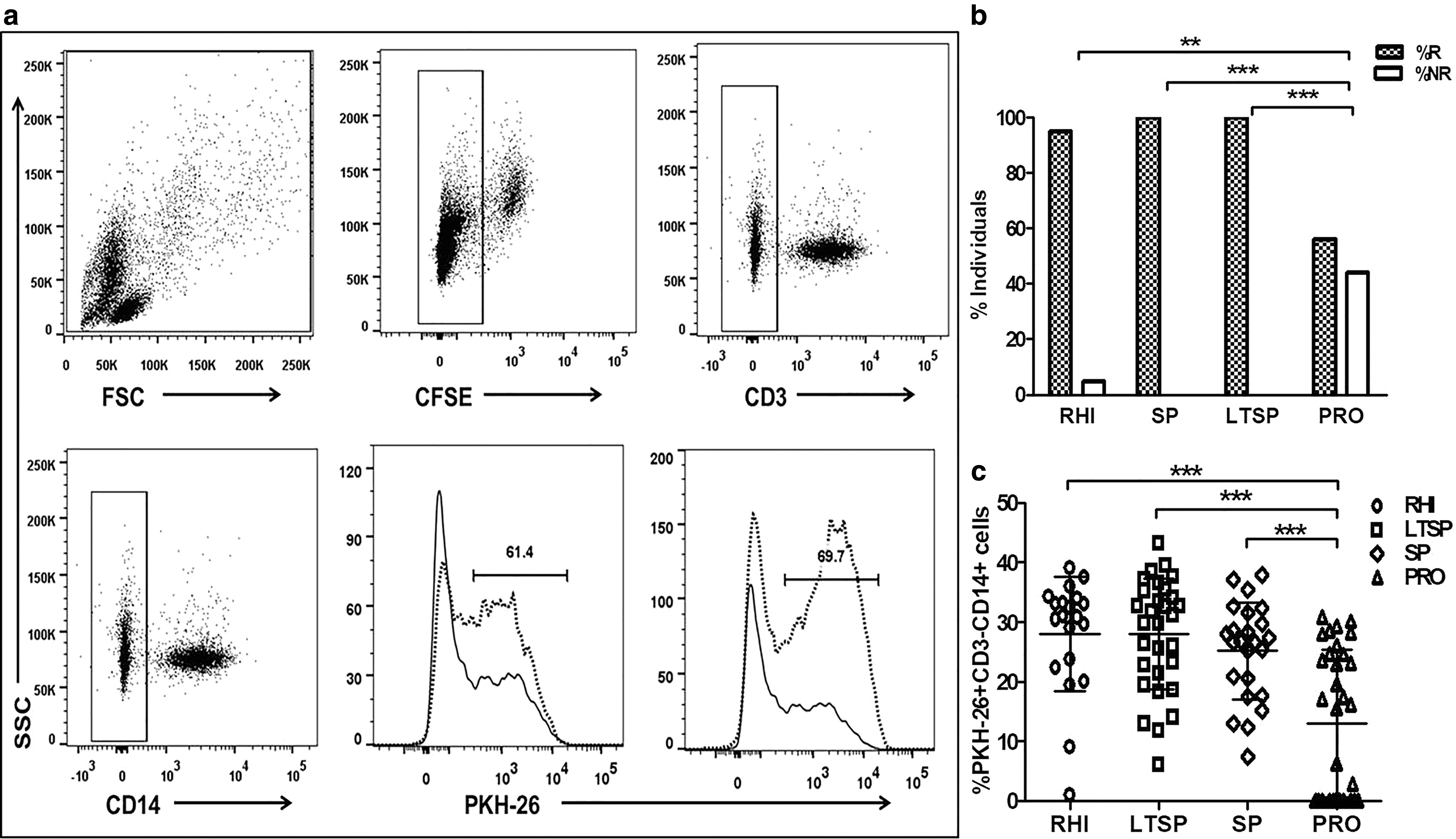
RFADCC assay. The gating strategy
ADCC-ICS assay
The anti-HIV antibody-mediated NK cell activation was measured in terms of IFN-γ secretion and expression of degranulation marker CD107a as described previously (7,30). Briefly, 150 μL whole blood from a healthy donor (as a source of NK cells) and 50 μL of heat-inactivated heparin anticoagulated plasma [final concentration as 1:4] from the study participants were incubated with or without HIV envelope protein (Env C prot) (NIH AIDS reference reagent program) in the presence of Brefeldin-A (10 μg/mL; Sigma), Monensin (0.68 μL/mL; BD Biosciences), and anti-CD107a APCH7 antibody (clone-H4A3) for 5 h at 37°C in 5% CO2. After incubation, cells were stained with anti-CD3 PerCP (clone-SK7) and anti-CD56 PECy7 (clone HCD56) (both from BioLegend) antibodies. The cells were then permeabilized using 1 × FACS perm II solution (BD Biosciences) before addition of anti-IFN-γ antibody (clone-B27) (BD Biosciences). The plasma+effector cells without Env C protein was considered as a negative control for the respective sample. The cells were acquired on FACSAria Fusion (BD Biosciences) and data were analyzed using FlowJo 10.0.6 software. The gating strategy is shown in Figure 4a. The lymphocytes were gated based on forward and side scatter and the NK cells were identified as CD3-CD56dim cells. The percentage of activated NK cells was represented as the sum of the percentages of NK cells expressing only CD107a/IFN-γ and both CD107a and IFN-γ. The response was considered to be positive if the total percentage of activated NK cells was greater than the mean of the percent activated NK cells observed in the samples of 20 HIV-negative donors plus 2SD.
Statistical analyses
Statistical analyses were performed using GraphPad Prism, version 5.0 (GraphPad Software, San Diego, CA). Two-tailed nonparametric t-test (Mann–Whitney) was used to carry out group comparison and Fisher's exact test was used for the binary data. Spearman rank correlation was used to test the correlation of ADCC responses with CD4+T cell count and viral loads. Wilcoxon matched pairs t-test was used to analyze the paired data. To compare genderwise differences in the ADCC responses (percent male and female responders), one-sided two-sample test of proportions was applied. p values <0.05 were considered significant.
Results
Demographic and immunological details of study participants at the study visit are given in Table 1. The age of all participants from LTSP, SP, and progressors were similar (p > 0.05), whereas RHI showed significantly lower age than that of LTSP, SP, and progressors (p < 0.0001 for three groups). At the study visit, the median CD4+T cell counts in SPs and LTSPs were 749 cells/μL (interquartile range [IQR]: 644–967) and 819.5 cell/μL (IQR: −551 to 1012), respectively, which were significantly higher than that of progressors (median: 398.5 cells/μL, IQR: 341–446.8) (p < 0.0001 for both). While the median CD4+T cell counts in RHI group (median: 613.5 cells/μL; IQR: 467.5–737.8) was significantly lower than seen in SPs (p = 0.0152) and LTSPs (p = 0.033), and was significantly higher than seen in progressors (p < 0.0001). The median plasma viral load values in SPs and LTSPs were 1,620 HIV RNA copies/mL; (IQR: 171.3–9017) and 3,233 HIV RNA copies/mL (IQR: 117.3–26355). These values were significantly lower as compared with the median viral load in progressors (48,116 HIV RNA copies/mL; IQR: 8868–142172) (p < 0.0001 for both). The median plasma viral loads of RHI at the enrollment visit (median: 4,352 HIV RNA copies/mL; IQR: 434.3–31250) was statistically indifferent from LTSPs (p > 0.05 for both) and was significantly lower than progressors (p = 0.0003). RHI group had a median plasma viral load set point of 6,973 RNA copies/mL (IQR: 2160–19962 RNA copies/mL). The CD4+T cell count and viral loads were similar between SPs and LTSPs (p > 0.05). The mean CD4+T cell count for all visits in SPs (median: 785.5 cells/μL; IQR: 657–831.8) and LTSPs (median: 756 cells/μL; IQR: 632.3–1061) as well as mean plasma viral load throughout all follow-up visits in the cohort in SPs (median: 2,051 HIV RNA copies/mL; IQR: 190–9017) and LTSPs (median: 2,250 HIV RNA copies/mL; IQR: 210.3–16151) were similar to the plasma viral load values obtained at the study visit (p > 0.05).
Demographic and Immunological Details of Study Participants at the Study Visit
HC, healthy controls; IQR, interquartile range; LTSP, long-term slow progressor; RHI, recent HIV infection; SP, slow progressors.
ADCC responses at various stages of HIV infection
The ADCC-mediating antibodies were found to have appeared in RHI as early as 25 days after the estimated date of infection and 19 out of 20 (95%) RHI showed ADCC response within the first year of HIV acquisition with an average duration of 173 days after estimated date of HIV infection. The mean magnitude of the ADCC response (measured as % PKH26+CD3−CD14+ monocytes) shown by 20 RHI was found to be 27.99% (IQR: 22.75–33.80) (Fig. 1c).
Whereas in the established HIV infection, the mean magnitude of ADCC responses shown by 24/24 SPs, 30/30 LTSPs, and 18/32 progressors were 25.17% (IQR: 18.35–31.13), 28% (IQR: 20.93–35.35), and 12.98% (IQR: 0–24.68), respectively. Both the frequency (no. of responders) and the magnitude of ADCC responses were significantly higher in RHI (p = 0.037; p < 0.0001 respectively), SPs (p < 0.0001; p = 0.0004 respectively), and LTSPs (p < 0.0001; p < 0.0001 respectively) as compared with the response observed in progressors (Fig. 1b, c), while there was no significant difference in the magnitude of ADCC responses in RHI, SP, and LTSP groups (Fig. 1c) (p > 0.05).
ADCC responses from HIV-infected groups (RHI, SPs, LTSPs, and progressors) together were found to be associated with higher CD4+T cell count (p < 0.0001; r = 0.3752) (Fig. 2a) and lower plasma viral load measured at the study visit (p = 0.0121; r = −0.2440) (Fig. 2b). However, the ADCC responses observed within the first year of HIV infection in RHI did not show any significant association with plasma viral load set point but showed a trend of positive association (p = 0.07) (Fig. 2c).
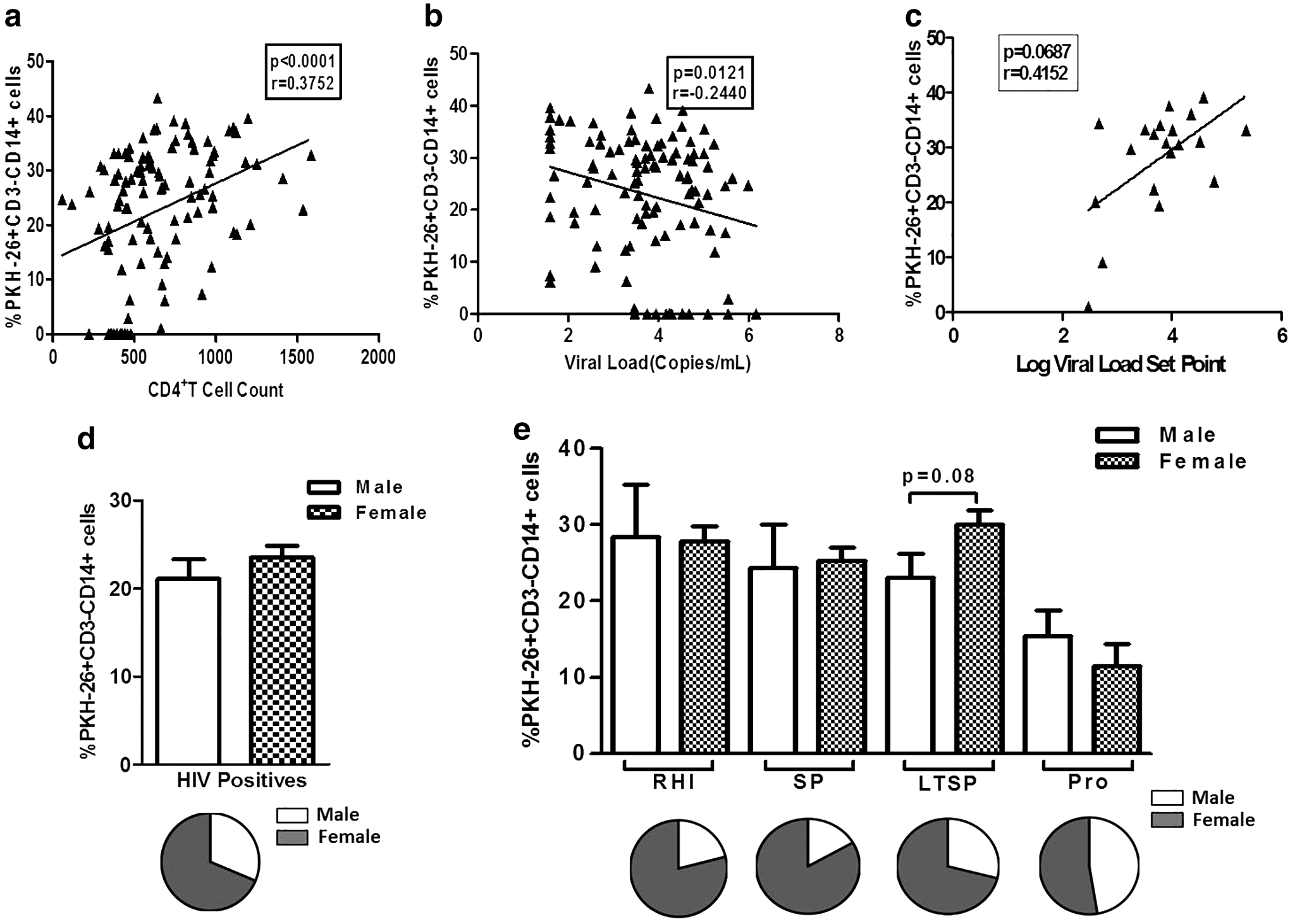
The scatter plots show the association of % PKH26+CD3−CD14+ cells (on Y-axis) with CD4+T cell counts (on X-axis)
We have previously reported higher reference CD4+T cell counts in Indian women as compared with the men (31), so considering the fact that the nadir CD4+T cell count would be higher in women as compared with men, we analyzed the magnitude of ADCC responses in male and female participants in study groups. Overall, the magnitude of ADCC responses was slightly higher in female participants (Fig. 2d). Within the study groups, especially in the LTSP group, the magnitude was higher in female participants, however, in the progressors' group, the responses were lower in female participants (Fig. 2e). The percent of female responders were higher overall and also in RHI, SPs, and LTSPs than the percent male responders but not significantly, whereas the percent of male and female responders was similar in progressors (Fig. 2d, e).
LTSPs and SPs showed increased ADCC activity over the time
Our cross-sectional analysis has shown that the nonprogressive HIV infection was associated with higher ADCC responses. We wanted to assess the usefulness of ADCC responses during the disease course. To analyze this we compared the RFADCC responses in the paired plasma samples from 18 study participants (13 LTSPs and 5 SPs) collected at visit 1 (when the participant was enrolled in the institutional cohort) and at the study visit. The duration between visit 1 and the study visit ranged between 4 and 15 years. The ADCC responses were found to be significantly increased at the study visit (mean: 26.47; IQR: 19.03–32.80) as compared with the visit 1 (mean: 19.67; IQR: 12.60–25.65) (p = 0.0378) (Fig. 3a). Interestingly the ADCC responses observed at visit 1 were correlated significantly although, marginally with lower plasma viral load measured at the study visit which was 4 to 15 years after the enrollment in the cohort (p = 0.055) (Fig. 3b). Although we could not carry out such analysis in progressors due to the unavailability of samples, data indicate that early ADCC responses might have a role in controlling plasma viral load in the later course of disease progression.
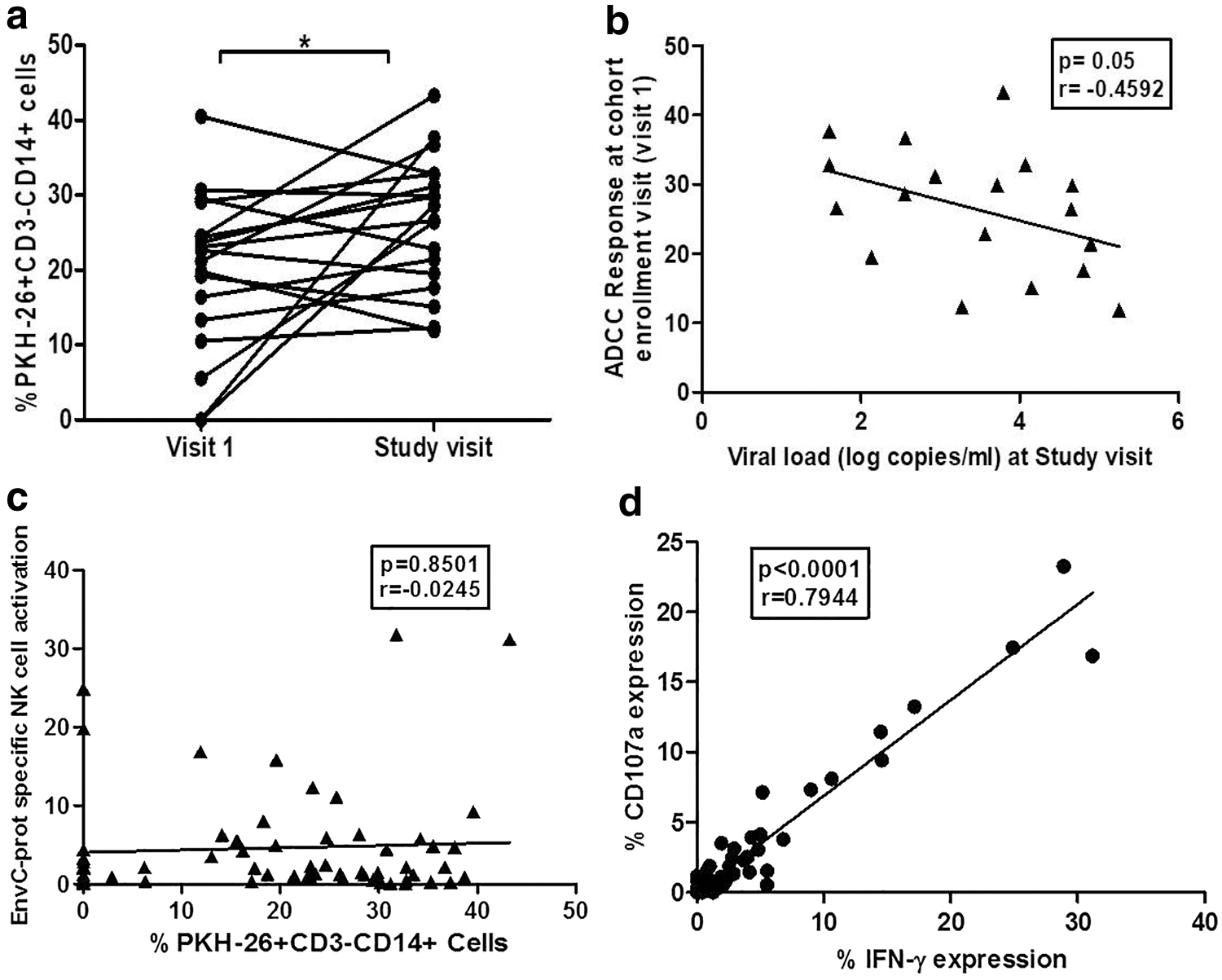
Functional ADCC responses observed over the time in the course of HIV disease. The graph
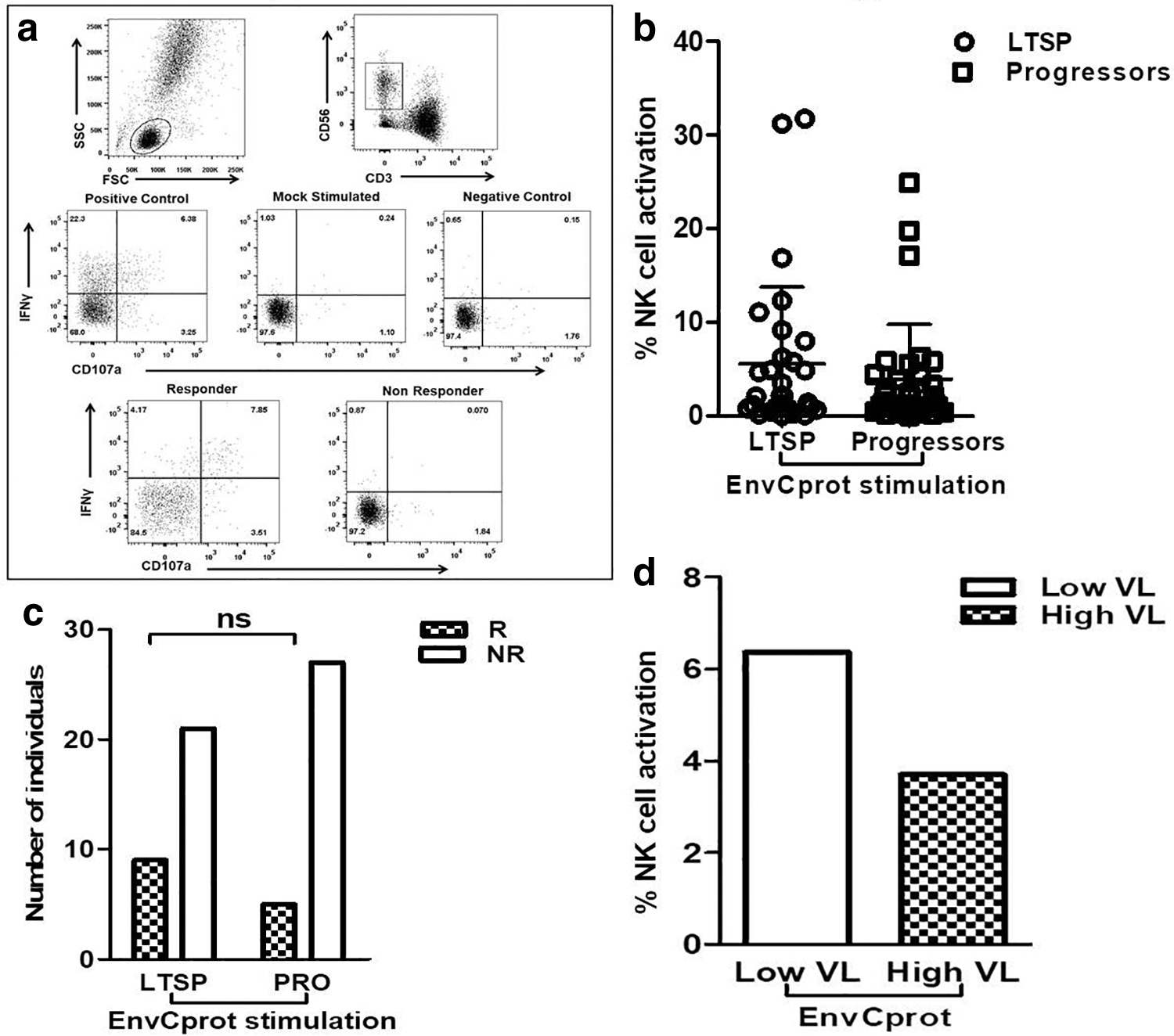
Antibody-dependent NK cell activation (ADCC) assay.
No correlation between the killing in RFADCC and the activated (IFN-γ secreting and degranulating) NK cells
The percent NK activation shown by 30 LTSPs and 32 progressors after HIV EnvCprot stimulation was 5.57% (IQR: 0.88–6.69) and 3.93% (IQR: 0.74–4.38), respectively; showing marginally higher NK cell activation in LTSPs as compared with the progressors but it was not statistically significant (Fig. 4b). The number of responders, 9 out of 30 (30%) LTSPs and 5 out of 32 (15.62%) progressors, was also similar (p > 0.05) (Fig. 4c). The NK cell activation was not associated with higher CD4+T cell count and or lower viral load (p > 0.05); however, in the LTSP group, the LTSPs with low viral load (<10,000 HIV copies/mL) showed the trend of higher % NK cell activation than those with high viral loads (>10,000 HIV copies/mL) (Fig. 4d). Although both the RFADCC and NK cell activation assay determine ADCC activity, no association was observed between the responses observed in both the assays (Fig. 3c). The envCprot-specific CD107a and IFN-γ expression by individual's NK cells showed strong association (p < 0.0001, r = 0.7944, Fig. 3d).
Discussion
The understanding of immune mechanisms contributing to the efficient HIV control is important in the development of efficacious HIV vaccine. The Fc-mediated functional characteristics of anti-HIV-1antibodies, such as ADCC, have been implicated in HIV control (3,4,7,9,10,16,24,25,27) and protection in the RV144 phase-3 HIV vaccine trial (15). The HIV infection leads to a number of antibody effector responses, but the functional coordination of antibodies and greater polyfunctionality of antibody effector functions is associated with control of the virus in the absence of ART (1). In this study, we determined the functional ADCC responses measuring the lysis of the target cells in the presence of effector cells using RAFDCC assay and HIV antibodies in individuals at various stages of the HIV disease progression, including RHI. The ADCC responses in RHI have been rarely studied as the identification of these individuals is difficult (8). We observed that the RFADCC response was detectable as early as 25 days after the estimated date of infection supporting the previous observation by Chen et al. (5). The viral load set point is known to predict the further HIV disease progression (4,5,12,17,27). In contrast to the findings of Chen et al., there was no association between the ADCC responses and plasma viral load set point, but we observed a trend of positive association (5). We could not assess the NK cell activation in these patients as performed by Chen et al. (5). Different readouts in both the studies could be one of the reasons for the different study findings. Also the sample size of RHIs in the present study could be the limitation.
We observed that RFADCC responses were detectable at all stages of HIV infection with higher responses in recent and nonprogressive HIV infection and these functional responses were associated with higher CD4+T cell count and lower plasma viral load confirming previous observations (7,16). We have previously showed similar findings using NK cell activation assay (18). However, the comparison between both the readouts of ADCC responses showed no association between them as reported by others (7). To avoid the variability in both assays we have used the same source of effector cells and the Env C protein in both the assays. Hence, the absence of the correlation might indicate that after stimulation, the different NK cell functions might not be occurring simultaneously as suggested by Chung et al. (7). Additionally, as reported by Wren et al., other serum factors like cytokines also affect ADCC antibody-induced NK cell activation profile (32). In agreement with this, we could observe some individuals showing high IFN-γ expression and low degranulation or vice versa, in spite of an overall association between EnvCprot-specific CD107a and IFN-γ expression (p < 0.0001, r = 0.7944, Fig. 3d). Other reasons for this absence might include different readouts of these assays, and difference in the antigen presentation to the effector cells. The RFADCC uses antigen-coated target cells while ICS assay uses primary cells as antigen-presenting cells. Considering our observations in the present and previous study (18), and also reported by others, we can argue that although the RFADCC and NK cell activation use different readouts and may not show significant association, both the assays underscore the potential role of ADCC-mediating antibodies in virus control (3,5,7,10,12,16,24,27,30).
However, this observation also confirms the requirement of more standardized assay easier to be adapted in clinical trial setting, in which the HIV vaccine generating ADCC-mediating antibody response is under investigation.
The Indian women are known to have high reference CD4+T cell count than men (31). Also at higher CD4+T cell levels (>350 cells/mm3), mean HIV-RNA has been noted to be lower in women (11). We did observe higher RFADCC responses in female participants than male participants, especially in LTSP group and higher number of female responders in all groups, except progressors. It might be possible that stronger ADCC responses in female contribute to their nonprogressive disease status. The additional data on longitudinal samples in both progressors and SPs would be helpful in obtaining more information regarding this.
In addition to the cross-sectional analysis of the association of RFADCC responses and the plasma viral load values, we were interested in understanding whether the ADCC responses could influence the viral load in the future disease course. Interestingly, we observed negative (although marginal) association between the plasma viral load values at the study visit with the earlier RFADCC responses detected for plasma of visit 1 (i.e., at the visit 4–15 years before the plasma viral load values were obtained). We also observed significant increase in the magnitude of RFADCC responses in these individuals. All these individuals had slow or no HIV disease progression. We could not assess the earlier responses in progressors; however, this observation indicates that sustained ADCC responses are important in controlling plasma viral load for a long period contributing to the nonprogression of HIV disease.
In summary, the study showed that ADCC responses are detectable in early HIV-1C-infected Indian individuals and the ADCC responses in established chronic HIV infection might contribute to the long-term virus control. The study also confirms the earlier observations that more standardized assays need to be developed that can be adapted in clinical trial setting for testing of HIV vaccine-mediating ADCC responses.
Footnotes
Acknowledgments
The authors thank the study subjects for their participation in this study. They also thank the staff of Departments of Immunology and Virology for providing the CD4 count and plasma viral load data. The investigators thank the University Grants Commission (UGC), Government of India, for supporting Ms. Sneha Talathi with CSIR-UGC-NET fellowship.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Sources of support: Institutional funding by National AIDS Research Institute, Pune.