
Abstract
Infection with influenza A (H1N1) virus contributes significantly to the global burden of acute respiratory diseases. Glucose uptake and metabolic changes are reported in different cell types after infections with different virus types, including influenza A virus. Alteration of glucose metabolism specifically in immune cells has major health consequences. The aim of this study was to monitor glucose concentration in unstimulated and stimulated U937 human monocytes with infectious or heat inactivated H1N1 or Staphylococcus aureus or in nonpathogenically stimulated monocytes with phorbol-12-myristate-13-acetate. Stimulated or unstimulated U937 human monocytes were subjected to H1N1 infection for different time points and the glucose profile in the growth medium was measured post infection. Results showed that regardless to whether the initial stimuli on U937 cells were of pathogen or nonpathogen origins, challenge infection by H1N1 causes a significant reduction of glucose levels 36 h post infection. In conclusion, H1N1 infection has a direct effect on the glucose uptake of U937 cells in vitro. This effect can be related to either H1N1 infection or cell differentiation status that might occur due to the exerted stimuli.
Introduction
In April 2009, the Center for Disease Control and Prevention identified two cases of 2009 pandemic influenza A (H1N1) virus infection in a boy and a girl from southern California (14). The virus rapidly spread worldwide (6) as >214 countries and overseas territories or communities reported laboratory-confirmed cases that created the 2009 WHO reported pandemic (pdm09), causing 18,449 deaths (29,30). Accordingly, the H1N1 virus became a worldwide major public health problem, including the Eastern Mediterranean Region (2).
Although monocytes are nondividing cells with a short half-life and an immediate immune response that makes viral replication difficult or almost impossible, viruses have found ways to adapt to replication in these cells (18). Macrophages are classified into two main classes known as M1 and M2, which express different functional programs in response to microenvironmental signals (27). This polarization is mainly dependent on the exposure to different stimuli from infectious or noninfectious origins (1,16,20).
Although cell culture systems are reductive and lack complexity of intact in vivo models, they, being also a cost-effective method, facilitate exploration of possible intracellular pathways and signaling processes when cells are challenged with conditions mimicking clinical situations. Furthermore, identification of mechanisms and pathways provides an understanding that may lead to novel therapeutic strategies (24).
U937 is a human hematopoietic cell line with exceptional characteristics derived from a patient with generalized histiocytic lymphoma (26). The origin of the cell line was shown by its capacity for lysozyme production and strong esterase activity indicating that the U937 is a neoplastic histiocytic cell line (26). U937 being (pro)monocytic cell lines can be differentiated in vitro into either various types of macrophages or dendritic cells in response to different stimuli (7). Soon these cells have become a diverse continental model in studying different biological parameters, including glucose metabolism (9,15,21,22,31).
Transformations of different cell types after infection with tumor viruses have been described to initiate both increased glucose metabolism and morphological changes (4,23). Increase in both uptake of extracellular glucose from, and release of lactate release into, cell culture supernatant was reported 12 h post infecting Madin–Darby Canine Kidney (MDCK) cells with H1N1 (A/PR/8/34) compared with mock-infected cells (19). U937 cells have also shown increased glucose uptake from their culture medium upon infection with type 1 human immunodeficiency virus (12). Recently, infection of U937 cell with H1N1 virus or stimulation by toxins of bacterial origin resulted in synergistic effects with recommendation of cautious interpretations (13).
The aim of this study was to investigate the glucose profiles in growth media of unstimulated, pathogen-stimulated, or nonpathogen-stimulated U937 cells before and after infection with H1N1 influenza virus.
Materials and Methods
U937 cells were propagated in RPMI medium (1640, Biowhittaker; Lonza, Belgium) supplemented with 10% fetal calf serum (FCS; Euroclone, Italy), and 10% of penicillin/streptomycin (Biowhittaker; Lonza) (28). For each assay, 1 × 106 cells/mL were used. Influenza A virus (H1N1 A/PR/8/34) was prepared by plasmid rescue system, counted and propagated in MDCK cells (3,11). U937 cells were induced by either infectious or heat inactivated H1N1 or Staphylococcus aureus at multiplicity of infections (MOI) 0.1. After 48 h, supernatants were harvested and stored at −80°C. Alternatively, U937 cells were treated with 100 μL/mL human serum (HS) collected from healthy donor, 100 ng/mL phorbol-12-myristate-13-acetate (PMA; Abcam, Cambridge, UK) or a combination of both to induce cell differentiation. Control untreated cells were included. After 24 h, both stimulated and unstimulated cells with all the previously mentioned stimuli were exposed to live H1N1 at an MOI of 0.1 for 12, 24, and 36 h. Cell supernatants were harvested at each time point from uninfected unstimulated cells (NI), H1N1 infected unstimulated cells (VL), H1N1 infected previously infectious H1N1 stimulated cells (LV VI), H1N1 infected previously heat inactivated H1N1 stimulated cells (KV VI), H1N1 infected previously infectious S. auerus stimulated cells (LB VI), H1N1 infected previously heat inactivated S. auerus stimulated cells (KB VI), H1N1 infected previously HS stimulated cells (HS VI), H1N1 infected previously PMA stimulated cells (PMA VI), and H1N1 infected previously HS and PMA stimulated cells (HS PMA VI). Culture medium controls were collected at 0 h from culturing U937 cells (M Cont 0 h). These were later subjected for quantification of glucose concentrations using the Precicheck blood glucose monitoring system (AC-300-AC-305 Fia Biomed GmbH, Emsdetten, Germany) which relies on glucose dehydrogenase activity. At each time point post each treatment, glucose concentration was assessed six times on six independent strips. Data were analyzed using the Graphpad Prism version 8.0.1 (GraphPad Software, Inc., La Jolla, CA) statistics software. Significance was assessed using the Student's t-test to compare uninfected to infected cells after 36 h or ordinary one-way analysis of variance (ANOVA) followed by Tukey's multiple comparisons test to compare different stimuli of infected cells after 12, 24, and 36 h. p-Values <0.05 were considered statistically significant.
Results
Glucose profile in medium of unstimulated U937 cells post-H1N1 infection
A significant decrease in glucose level was observed 36 h post-H1N1 infection (PII) compared with the M Cont 0 h and U937 control cells after 36 h (n = 6, p < 0.005; Fig. 1).

Glucose profile in medium of unstimulated U937 cells PII: 1 × 106 U937 cells/mL were exposed to live H1N1 at an MOI of 0.1 for 36 h. A significant decrease in glucose level was observed 36 h PII compared with M Cont 0 h and U937 cells control after 36 h (n = 6, ***p < 0.0005). MOI, multiplicity of infections; PII, post-H1N1 infection.
Glucose profile in medium of U937 cells challenged with live or inactivated H1N1 or S. aureus then actively infected by H1N1
Initially stimulated U937 cells with LV (A), KV (B), LB (C), or KB (D) all at MOI = 0.1 showed a significant decrease in glucose levels at 36 h PII compared with the M Cont 0 h and stimulated U937 control cells after 36 h (n = 6, p < 0.05; Fig. 2).
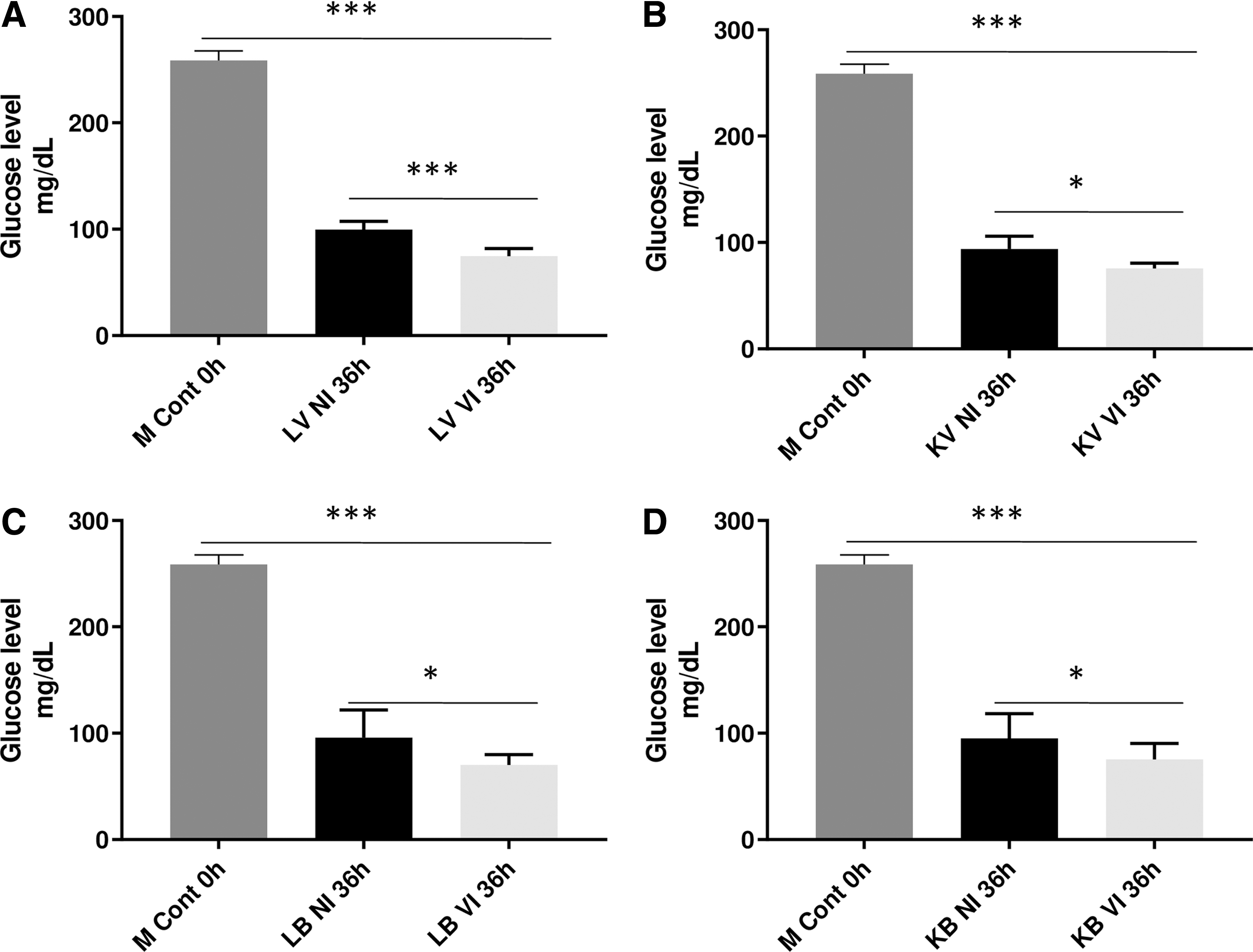
Glucose profile in medium of U937 cells challenged with live or inactivated H1N1 or Staphylococcus aureus then actively infected with H1N1: 1 × 106 U937 cells/mL were induced by either infectious or heat inactivated H1N1 or S. aureus at MOI 0.1, then exposed to live H1N1 at an MOI of 0.1 for 36 h. Initially stimulated U937 cells with LV
Glucose profile in medium of U937 cells stimulated by PMA or HS followed by H1N1 infection
Initially stimulated U937 cells with 100 μL/mL HS (A), 100 ng/mL PMA (B) or a combination of both (C), showed a significant decrease in glucose levels at 36 h PII compared with the M Cont 0 h and stimulated U937 control cells after 36 h (nonsignificant in case of HS. PMA stimulation; n = 6, p < 0.05; Fig. 3).

Glucose profile in medium of U937 cells stimulated by PMA or HS followed by H1N1 infection: 1 × 106 U937 cells/mL were treated with 100 μL/mL HS, 100 ng/mL PMA or a combination of both then were exposed to live H1N1 at an MOI of 0.1 for 36 h. Initially stimulated U937 cells with 100 μL/mL HS
Multiple statistical comparisons of glucose levels in media of U937 initiated by different stimuli followed by H1N1 infection
Glucose measurement at each stimulus was plotted and compared at 12 h (Fig. 4A), 24 h (Fig. 4B), and 36 h (Fig. 4C) PII. The initial assessment of statistical significance by one-way ANOVA (n = 6, p < 0.05; Fig. 4) was followed by multiple comparisons of effect of each stimulus to the others at each time point PII of U937 and results are summarized in Table 1.
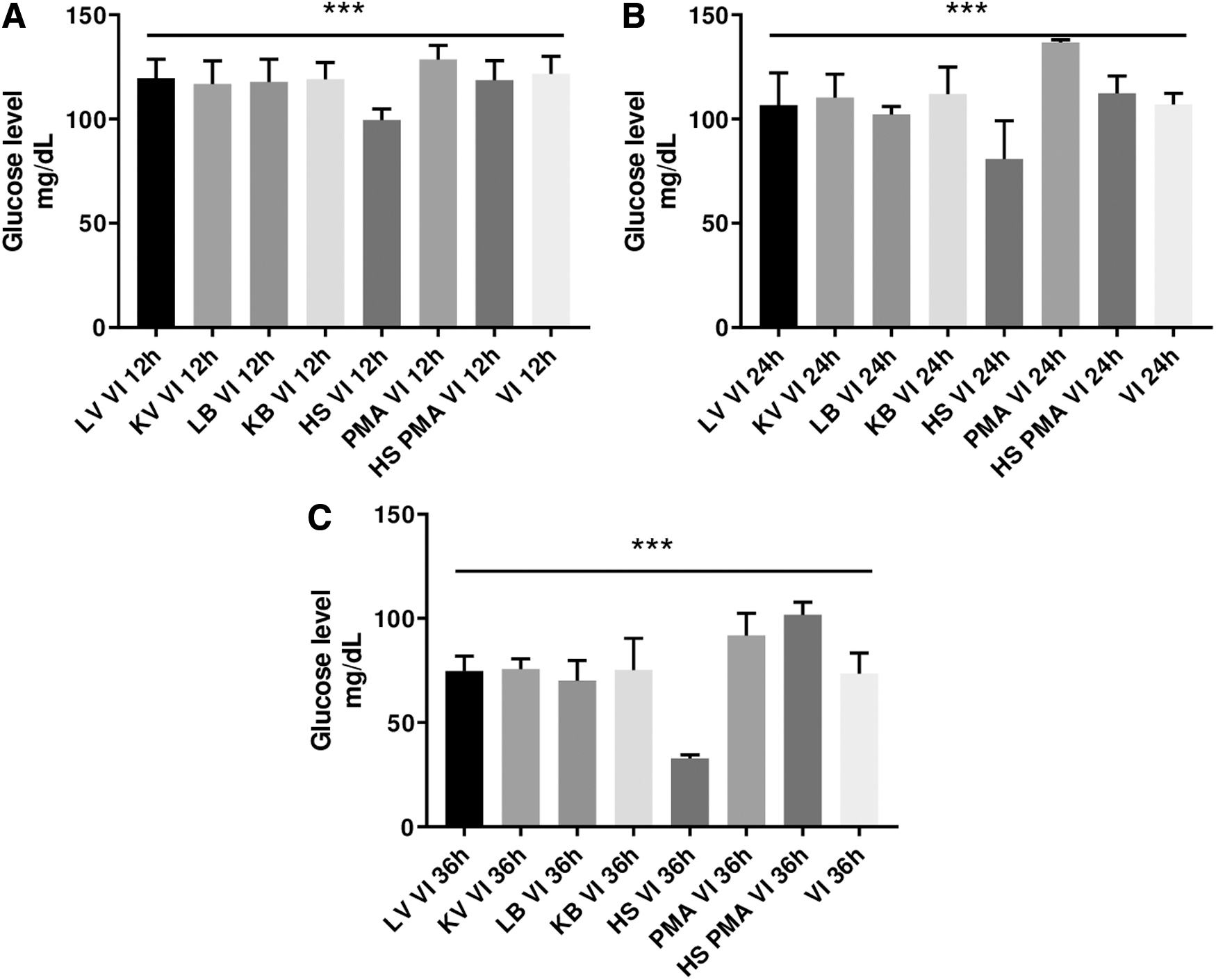
Statistical comparisons of glucose levels in media of U937 initiated by different stimuli followed by H1N1 infection: glucose measurement at each stimulus was plotted and compared at 12 h
Multiple Statistical Comparisons of Glucose Levels in Media of U937 Initiated by Different Stimuli Followed by H1N1 Infection
HS, human serum; PMA, phorbol-12-myristate-13-acetate.
Discussion
The key finding in this study is the significant decrease of glucose concentration in growth media of stimulated U937 monocytes which were later subjected to infection with H1N1. Regardless to the initial stimuli, glucose uptake started at 12 h and reached its maximum at 36 h PII. This may support the conclusion of Ritter et al. (19) that influenza virus infection had its main impact on glucose metabolism during late stages of the virus replication and that the early virus replication phase was not a critical burden for cellular metabolism. However, taking into account that in Ritter et al.'s study, the drop in glucose concentration was recorded after 12 h of infecting MDCK cells, whereas in our study the drop in glucose concentrations in the growth media significantly peaked at 36 h post infecting U937 monocytes. This difference may be due to the differential adaptation of the virus and the differential susceptibilities of the two cell lines to influenza infection. Ritter et al. (19) related metabolic changes to apoptosis of the cells and not to the virus replication itself, while in our hands, the cell viability was constant during all time points post H1N1 infection (viability data are not included) which may either suggest the ability of the H1N1 to adapt to replication in monocytes or alternatively the attempt of monocytes to hinder virus replication.
When U937 monocytes were prestimulated with viral or bacterial antigens before being exposed to active H1N1 infection, the glucose concentration showed the same profile such as unstimulated cells with regard to the significant uptake of glucose from the medium after 36 h of infection. It has been shown that incubation of peripheral blood monocytes with live influenza induced a significant increase in the expression of co-stimulatory molecules (e.g., cytokines with direct antiviral effect and molecules that could augment other immune cells activation) which are distinguished from those expressed after inactivated virus stimulation (5).
Live bacteria induced secretion of inflammatory mediators when stimulating human monocytes (8,10). Monocytes stimulated with heat inactivated S. auerus have demonstrated expression of tumor necrosis factor alpha, interleukin (IL)-1β, and IL-6 production as early as 4 h after exposure suggesting synergistically induced inflammation and activation upon H1N1 infection (13). Clear differential production of nitric oxide upon monocyte stimulation with live or killed bacteria was also observed (10). In our study, cells pretreated with either live or killed virus or bacteria showed statistically significant pattern of glucose uptake from the medium and the glucose uptake profile was almost the same in cases of all stimuli which might further question whether the glucose uptake profile is related to H1N1 adaptation to replicate in monocytes or to the monocytes trials to interfere with virus replication.
Both HS factors and PMA, being the common chemical inducer of monocyte differentiation, stimulate U937 monocyte propagation to macrophages (25,28). Our results reveal that PMA-stimulated U937 cells are highly resistant to the uptake of glucose from the medium compared with HS-stimulated U937 cells. It might appear that PMA hinders the uptake of glucose from the growth medium when used in combination with HS to stimulate U937 monocytes. This might be related to multiple mechanisms suggested to be exerted by HS factors to induce monocyte differentiation which supports the notion that mechanisms to activate macrophages are very heterogeneous and oppose the conventional concept of M1/M2 binary activation (17,25).
Conclusion
This study contributes to the increasing awareness of monocyte and macrophage diversity when it comes to stimuli from their microenvironment. In addition, it provides new evidence on the usefulness of glucose uptake and metabolism as markers for challenging human monocytes with H1N1 infection regardless to the initial stimuli the cells were exposed to. This can be related to H1N1 adaptation to replicate in macrophages or the macrophages exerted mechanisms to stop virus replication.
Footnotes
Authors' Contributions
M.M.B. designed the experiments, critically discussed the results with all authors, and suggested the structure of the article. K.A. performed the experiments, discussed and statistically analyzed the results, and drafted the initial article. D.N.A. supervised and trained K.A. on cell culture performance, virus titration, and kinetic monitoring of infected cells. T.K.M., A.S.M., N.N.S., M.M.B., D.N.A., and M.K. proposed additional experiments, discussed and critically analyzed the results, edited and revised the article. All authors read and approved the submitted article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by a governmental fund from the National Research Center (NRC) of Egypt to support the PhD thesis of K.A. and additional NRC grants awarded to A.S.M., D.N.A., and M.M.B. M.M.B. particularly acknowledges Prof. Richard Webby from St. Jude Children Research Hospital, Memphis, who provided the plasmids used to generate the PR8-H1N1 through a signed material transfer agreement with the National Research Centre of Egypt.