
Abstract
The porcine circovirus type 2 (PCV2) capsid (Cap) protein and porcine parvovirus (PPV) VP2 protein have been studied in vaccines to control postweaning multisystemic wasting syndrome (PMWS). Virus-like particle (VLP) vaccines are nonreplicative vectors that deliver epitopes and induce immune responses. However, most VLP vaccines are recombinant proteins expressed in eukaryotic systems and are expensive and complex. In this study, the full-length PCV2-Cap and PPV-VP2 proteins were expressed in Escherichia coli, which self-assembled into VLPs. The highly soluble proteins were purified using Ni-chelating affinity chromatography. The proteins self-assembled into VLPs of ∼20 nm (Cap VLP) and 25 nm (VP2 VLP) in diameter. The immunogenicities of Cap VLP and VP2 VLP were determined in piglets coinfected with PPV and PCV2 postimmunization. The results suggested that Cap VLP and VP2 VLP did not antagonize each other. The combined vaccine induced stronger humoral and cellular immune responses and provided the best protection against PPV and PCV2 coinfection. On a farm containing PMWS-infected pigs, the combined Cap VLP and VP2 VLP vaccine significantly improved piglet growth indices; the average daily weight gains were significantly higher than those of the Cap VLP vaccine and nonimmunized groups. Thus, Cap and VP2 protein expression in E. coli is feasible for large-scale VLP vaccine production. The combined vaccine may be a promising candidate vaccine for better preventing PMWS-associated diseases coinfected with PCV2 and PPV.
Introduction
Postweaning multisystemic wasting syndrome (PMWS) is an emerging disease that commonly affects piglets at 3–18 weeks of age, with mortality ranging from 4% to 20% (2). The typical clinical signs of PMWS are enlarged lymph nodes, progressive weight loss, diarrhea, icterus, and respiratory disease, which are widespread in swine herds (24). Porcine circovirus type 2 (PCV2) is the etiologic agent of PMWS (1). However, single PCV2 infections do not always develop into PMWS, and the full clinical disease is believed to arise in combination with other cofactors (29), such as other viruses, environmental conditions, or stimulation due to stress reactions (20,28). Experiments have proved that PCV2 coinfected with porcine parvovirus (PPV) causes a more severe disease than that of a single infection. Many researchers believe that the initial viral coinfection occurs in animal macrophages where PPV stimulates macrophage proliferation, regulates the production of related cytokines, and affects the host immune system and apoptosis, thus providing suitable conditions for PCV2 infection and replication. These factors cause PCV2 accumulation and aggravate the clinical symptoms of PMWS (3,18,19, and 27).
VP2 protein is the major PPV capsid (Cap) protein and is essential for viral infection and protection in animals (4,14). The PCV2 Cap protein, encoded by the viral ORF2 gene, is the major structural protein and has a type-specific epitope (12,36). VP2 and Cap proteins assembled into virus-like particles (VLPs) induce a high-level neutralization antibody response and provide effective immunoprotection in animals. Conversely, the proteins unassembled into VLPs do not induce the neutralization response, which is based on recognition of different protein structural forms. Antibodies are generally directed against the whole virion, which induces neutralizing antibody (NA) production. However, recognition of monomer proteins results in production of nonneutralizing antibodies against the immunodominant epitope (33,35). The VP2 and Cap proteins are ideal target antigens for new vaccine research.
Vaccine immunization is the primary means of preventing PPV and PCV2 diseases. To effectively prevent PPV and PCV2 diseases, commercial vaccines mainly include inactivated vaccine, attenuated vaccine, chimeric vaccine, DNA vaccine, and subunit vaccine (5). However, the effect is poor for preventing PMWS coinfected with PPV and PCV2 in piglets. In China, only first-born sows are immunized with the PPV vaccine, which may be another reason PMWS is common (21).
In this study, soluble Cap and VP2 proteins were expressed in an Escherichia coli expression system, and the purified Cap and VP2 proteins self-assembled into VLPs in Tris-buffer. The two VLPs were mixed and emulsified with ISA201 adjuvant, and then the piglets were immunized with Cap and VP2 VLPs together or singly. Evaluating the humoral and cellular immune responses confirmed that a combined Cap-VP2 VLP vaccine was produced. This vaccine is safer, more effective, and cheaper to produce. The combined vaccine prevents multiple immunizations, which greatly optimizes the immunization procedure and reduces the clinical stress caused by vaccine immunizations. In addition, the combined vaccine prevented both viruses simultaneously, among which the Cap VLP can effectively reduce the PCV2 infection. Moreover, the VP2 VLPs of the combined vaccine both reduced the PPV infection and significantly reduced the severe PCV2 disease caused by PPV coinfection, thus effectively preventing PMWS. The combined vaccine achieved the idea of one needle protecting against two diseases and promoted control of PMWS. This study provides insight into a combined VLP vaccine against PMWS coinfected with PPV and PCV2.
Materials and Methods
Animals, viruses, and cells
Three-week-old piglets were selected based on serological diagnosis, PPV and PCV2 infection detected via real-time polymerase chain reaction (PCR), and the antibodies detected via hemagglutination inhibition (HI) and enzyme-linked immunosorbent assay (ELISA), respectively. PRRSV and PRV antibody presence was confirmed to be negative via ELISA.
PPV (NJ strain) and PCV2 (CD strain) were maintained in our laboratory. The titers of the pooled ST and PK-15 cells were 107.5 TCID50/mL and 106.5 TCID50/mL, respectively. The experimental challenge model of coinfection with PPV and PCV2 was confirmed to replicate the typical symptoms of PMWS and was used to evaluate the vaccine's effectiveness against PCVAD. The PQZ vector was constructed in our laboratory from the pET24a (+) vector.
The experiments were carried out in the livestock infection disease facility at the National Research Center of Engineering and Technology for Veterinary Biologicals (Nanjing, China).
Expression and purification of the recombinant proteins in E. coli
The entire PCV2 ORF2 gene and PPV VP2 gene optimized through E. coli codons usage were designed based on PCV2 (GenBank accession no. HM009332) and PPV (GenBank accession no. JQ710893), with BamHI/HindIII site at both ends. The optimized nucleotide sequences were synthesized (Genscript, Nanjing, China) and cloned into the PUC57 vector and constructed with the recombinant plasmids PUC57-Cap and PUC57-VP2. The recombinant plasmid was treated with BamHI/HindIII, and then cloned into the expression vector PQZ to generate the recombinant expression plasmids PQZ-Cap and PQZ-VP2. The accuracy of the target gene was confirmed by restriction enzymes and DNA sequence analysis.
The recombinant expression plasmids PQZ-Cap and PQZ-VP2 were transformed, respectively, into E. coli BL21(DE3), and the single clone of the transformant was grown in Luria-Bertani (LB) medium with 50 μg/mL ampicillin at 37°C overnight with shaking at 220 rpm. The culture was diluted at 1:80 in fresh LB medium and incubated at 37°C with shaking at 220 rpm until the OD600 reached 0.6–0.7. Isopropyl β-
Purification of protein VLPs and vaccine preparation
Soluble Cap and VP2 proteins were purified via Ni-chelating affinity chromatography (Thermo, Shanghai, China) followed by desalting through ultracentrifugation. The pellet was resuspended with Tris buffer (50 mM Tris, 150 mM NaCl, 0.2% Triton-X 100, 1 mM protease inhibitor, pH 7.2) overnight for VLP self-assembly. Transmission electron microscopy (TEM) was used to confirm the self-assembled into VLPs of two proteins. Concentrations of the two VLPs were determined using BCA Kit (Thermo) using bovine serum albumin as a standard. Purified VLPs were diluted to 500 μg/mL, and then emulsified with ISA201 adjuvant (SEPPIC, Paris, France) at a 1:1 volume ratio water-in-oil-in-water as per the manufacturer's instructions.
Experimental design
The National Research Center of Engineering and Technology for Veterinary Biologicals approved the experimental protocol for both studies. Table 1 summarizes the experimental design.
Experimental Design
VLP, virus-like particle; PBS, phosphate-buffered saline; Cap, capsid.
Trial 1 (immune efficiency under experimental infection)
Forty 3-week-old piglets, which were confirmed to be free of PPV and PCV2, were randomly divided into 5 groups, and groups 1–3 were immunized intramuscularly with 2 mL of the corresponding vaccine. A booster dose with 2 mL was given 2 weeks later, and all groups were housed in one room until 1 day before challenge. After 14 days, animals in groups 1–4 were challenged intranasally and intramuscularly with combined PCV2 and PPV; the dual inoculum was administered intranasally at 0.5 mL per nostril and 2 mL intramuscularly in the neck. The challenge doses of PPV and PCV2 was 1.25 × 107.5 TCID50 and 1.25 × 106.5 TCID50, respectively. After inoculation, the piglets were housed separately.
Trial 2 (immune efficiency under natural conditions)
This experiment was conducted on a farm in northern Jiangsu Province. The farm had shown a clinical disease pattern of PMWS ∼1 year before the experiment. Ninety 3-week-old piglets of commercial crossbreeds were confirmed to be positive for presence of the PCV2 and PPV antibody and antigen, which were randomly divided into three groups. Table 1 summarizes the vaccine administration; group A and B immunized intramuscularly with 2 mL corresponding vaccine. A booster dose with 2 mL was given 2 weeks later. Daily weight gain was chosen as the primary study parameter. All groups were housed in one room, and the experiment was terminated at 18 weeks of age.
Clinical evaluation
Following the challenge, the piglets' rectal temperatures were recorded daily until 28 days postchallenge (dpc). All animals were monitored daily for clinical signs of PMWS such as coughing, dyspnea, pallor, lethargy, and diarrhea. In trial 1, all piglets were weighed before challenge and at 35 dpc. In Trial 2, all piglets were weighed before immunization and at 18 weeks. The mean average daily weight gain (ADWG: g/pig/daily) was compared for each group.
Determination of antibody titers
In Trial 1, all serum samples collected at −14, 0, 7, 14, 21, 28, and 35 dpc were detected for the presence of immunoglobulin G (IgG) and neutralization antibodies.
PCV2 IgG was detected using a commercial ELISA Kit (Keqian, Wuhan, China). The absorbance reading of each well at 630 nm was recorded using a spectrophotometer. The results were expressed as the ratio of OD630 produced by the immunized groups compared with the negative group. Samples with a ratio >2.1 were considered positive. PPV antibody titers were calculated via HI assay, all serum samples were two-fold diluted until the samples were diluted to negative; the reciprocal of the highest serum dilution was the antibody titer.
A PCV2 NA assay was conducted using indirect fluorescence antibody titration. The serum was diluted twofold in minimum Eagle's medium starting at 1:2, which was heat-inactivated at 56°C for 30 min. An equal volume of PCV2 (1000 TCID50/mL) was added to the serum samples and incubated at 37°C for 1 h. The mixture was then inoculated into 96-well plates containing 50–60% confluence of PK-15 cells and incubated at 37°C for 72 h. Next, the plates were fixed with absolute ethyl alcohol at 4°C for 30 min, then washed with phosphate-buffered saline (PBS) (pH 7.2) three times, and dried and incubated with a 200-fold dilution of antibody to PCV2 at 37°C. After 1 h, the plates were washed five times with PBS, then stained with fluorescein isothiocyanate-labeled SPA (Boshied, Wuhan, China). Cells were visualized using an inverted fluorescence phase-contrast microscope (Zeiss, Guangzhou, China). The neutralization titers were determined as the reciprocal of the highest dilution of serum sample that provided 50% protection for the cells.
Twofold dilutions were replicated starting at 1:10. Heat-inactivated (56°C, 30 min) serum samples were mixed with 1000 TCID50 of the PPV virus equally and incubated at 37°C for 1 h. The mixture was then inoculated into 96-well plates containing 30–40% confluence of ST cells and incubated at 37°C for 7 days. The PPV neutralization titers were determined as the reciprocal of highest serum dilution that induced 50% neutralization under a light microscope.
Cytokine analysis
Peripheral blood mononuclear cells (PBMCs) were isolated from serum samples (Trial 1) collected at 0, 7, 14, 21, 28, and 35 dpc using the density gradient method (Dingguo, Beijing, China) as per the manufacturer's instructions. After isolation, total RNA was extracted using TRIzol reagent (Takara, Dalian, China). The total RNA (DNase-treated) was reverse-transcribed into cDNA using a reserve transcription kit (Takara). The cDNA were used as templates to determine the cytokine mRNA expression (interleukin [IL]-8, tumor necrosis factor [TNF]-α, IL-1β, IL-10, and IFN-γ) via quantitative real-time PCR. Table 2 lists the details of each primer set used for detection. The amplification was performed in 20 μL reaction mixture containing 10 μL 2 × SYBR Master Mix (YIFEIXUE, Nanjing, China), 0.4 μL of each primer, 7.2 μL nuclease-free water, and 2 μL of the extracted DNA. The reaction was run in a real-time thermocycler (LightCycler; Roche, Swiss) with the following cycling times: 1 cycle at 95°C for 10 min and 40 cycles at 95°C for 15 sec and 60°C for 60 sec.
Primer Sequences Used for Quantitative SYBR Green-I Real-Time Polymerase Chain Reaction Amplification
PCV2, porcine circovirus type 2; PPV, porcine parvovirus; IL, interleukin; IFN, interferon; TNF, tumor necrosis factor.
Each experiment included the no-template control (NTC) and no-reverse-transcription controls; the NTCs were assumed to be negative and reliable if the quantification cycle was >35.
Quantification of viral load in the samples
DNA was extracted from the samples using a DNA mini kit (Takara) as per the manufacturer's instructions. To quantify the PPV and PCV2 viral loads, extracted DNA was analyzed in triplicate via real-time PCR. The reaction mixture and reaction process were as previously described in section “Cytokine analysis”.
DNA was quantified by comparing the standard curve generated using 10-fold diluted plasmid standards of 102–1010 copies/μL, respectively. The cutoff level for a positive sample was set as 102 template copies based on validation experiments. Table 2 lists the details of each primer set.
Gross pathology and histopathology
In Trial 1, all piglets were euthanized and necropsied at 35 dpc. Gross lesions in the heart, liver, spleen, lungs, kidneys, tonsils, brain, thymus, and lymph nodes were recorded for each animal. All grossly evident lesions were collected and fixed in 10% neutral buffered formalin. All samples were fixed, processed, embedded in paraffin wax, sectioned, and stained with hematoxylin and eosin for light microscopic evaluation.
Statistical analysis
The experimental data were statistically analyzed using SPSS (v20.0; IBM, Armonk, NY) via two-way analysis of variance. The results are expressed as the mean ± standard deviation.*p < 0.05 was considered statistically significant, and **p < 0.01 was considered to indicate a high degree of significance compared with the control group.
Results
Characterization and purification of recombinant proteins
The recombinant expression plasmids PQZ-Cap and PQZ-VP2 were constructed to efficiently express the Cap and VP2 proteins in E. coli. High-level production soluble proteins of 27 kDa for Cap and 67 kDa for VP2 were found in the supernatant fraction via SDS-PAGE, respectively. Specificity was verified via WB using anti-PCV2 and anti-PPV swine immune sera, respectively (Fig. 1).
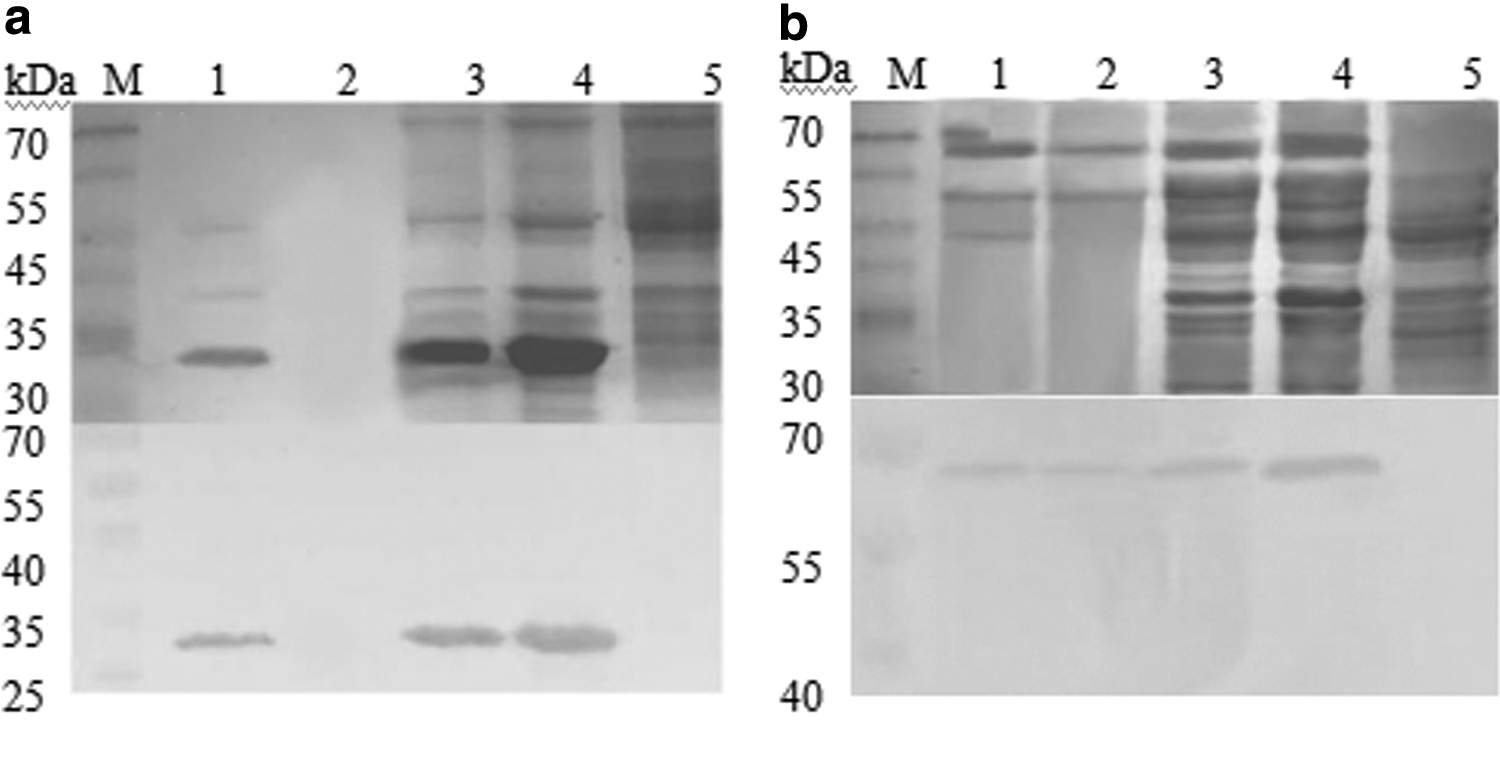
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and western blotting for detection of Cap and VP2 fusion proteins produced in Escherichia coli BL21.
The purified proteins were observed through TEM to confirm that the VLPs had self-assembled in the Tris buffer. The Cap proteins appeared as VLPs with diameters of ∼20 nm (Fig. 2a), and the VP2 proteins self-assembled into VLPs with diameters ranging from 20 to 25 nm (Fig. 2b).

Electron microscopy analysis of purified Cap VLPs
Clinical evaluation
In trial 1, no clinical signs of disease were observed in groups 3 and 5, and all animals remained healthy until necropsy at 35 dpc. Piglets in groups 1 and 2 showed lethargy, anorexia, mild dyspnea, and intermittent fevers. The control animals in group 4 had significantly higher mean temperatures than those of the other groups from 9 to 16 dpc (Fig. 3). Group 4 also began to show PMWS symptoms at 14 dpc, including coughing, weight loss, diarrhea, jaundice, and other typical symptoms, and 2/5 piglets had red skin lesions on their limbs and abdomen. Clinical disease began to resolve after 28 dpc, and most animals returned to normal by 35 dpc.
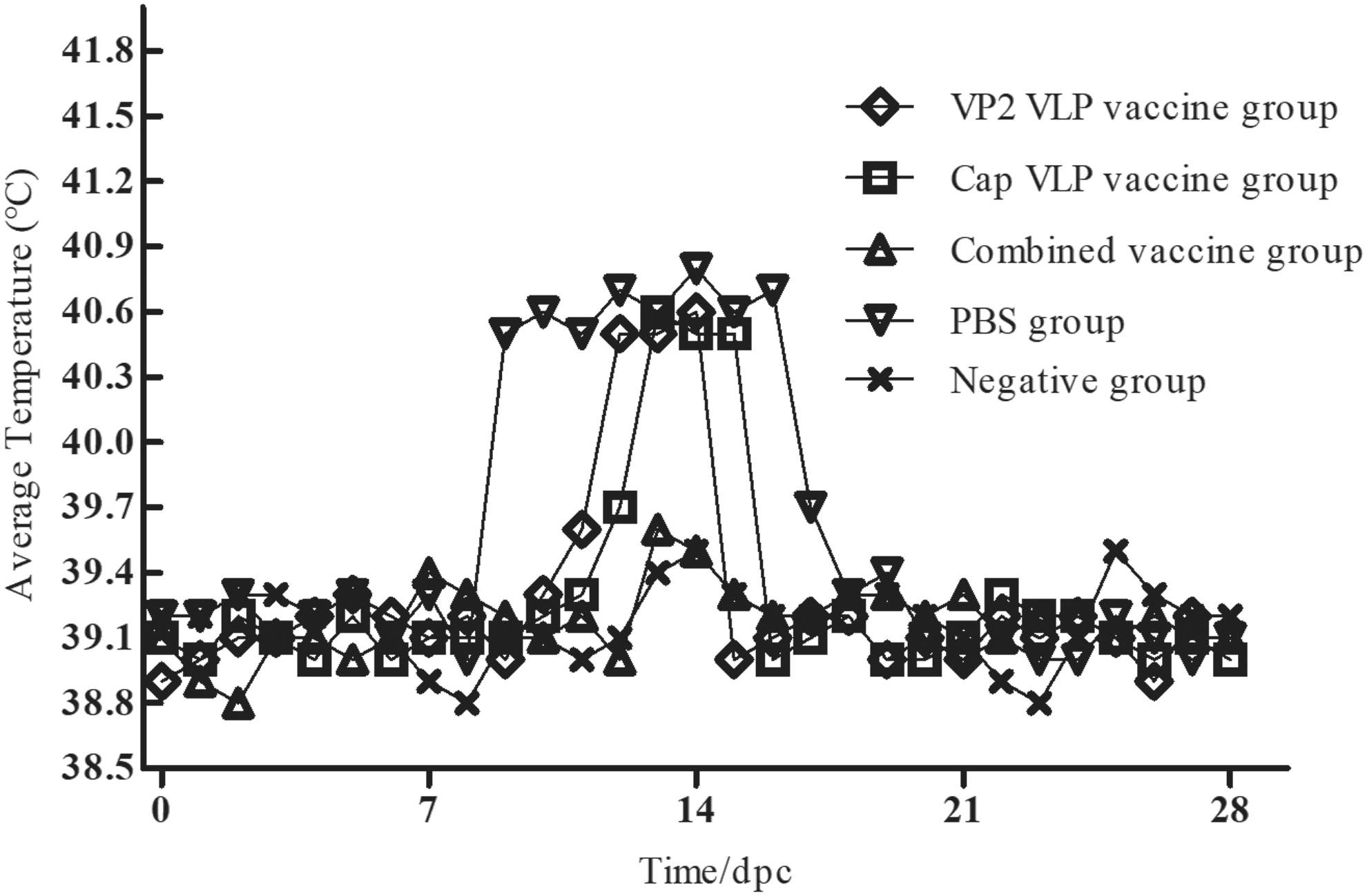
Rectal temperatures in each group from 0 to 28 dpc. dpc, days postchallenge.
In trial 1, the ADWG did not differ between groups 3 and 5, but was significantly higher than that of the single vaccine groups (p < 0.05). The control piglets in group 4 showed severe wasting; two lost ∼2.5 kg between 14 and 21 dpc, and the ADWG was significantly lower than in the combined vaccine groups (p < 0.01; Fig. 4).
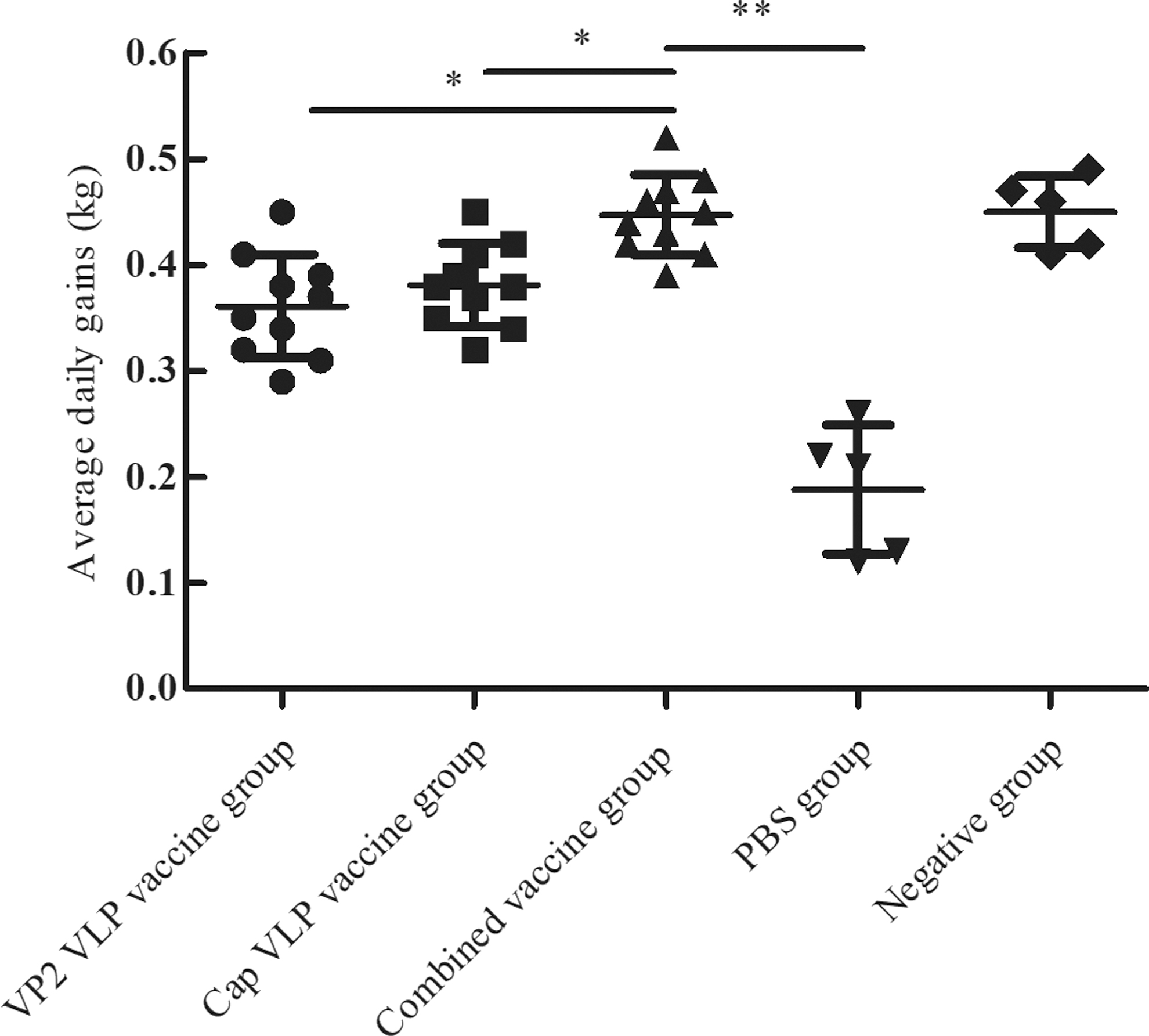
Daily weight gain for each experimental group after challenge. The mean daily weight gains were compared for each group. Data were analyzed via analysis of variance at 35 dpc. *p < 0.05 was considered statistically significant; **p < 0.01 was considered highly significant.
In trial 2, all piglets were weighed at 18 weeks. The overall growth performance of the combined vaccine group was significantly higher than that in the other two groups (p < 0.01). Most piglets in the negative group showed severe wasting, and three only gained 10 kg during the experiment (Fig. 5).
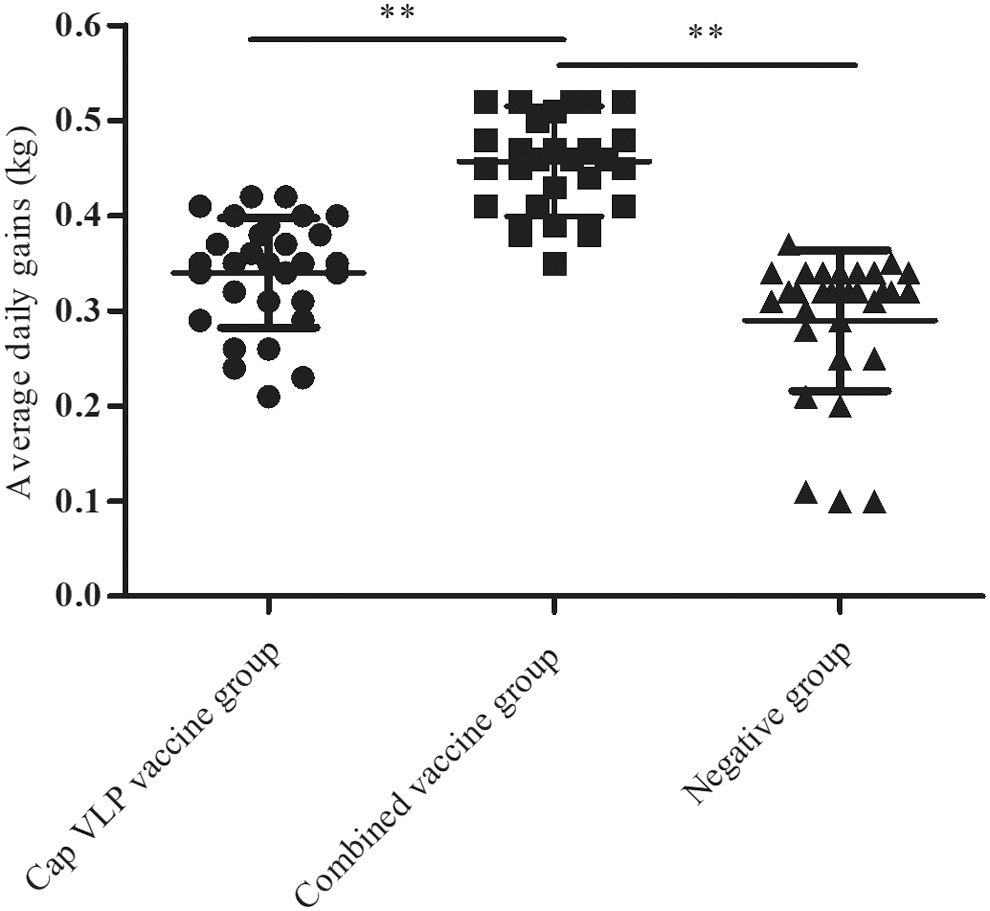
The daily weight gain for each experimental group. **p < 0.01 was considered highly significant.
Determination of serum antibody titers
Figure 6a summarizes the specific PCV2-antibody titers at −14, 0, 7, 14, 21, 28, and 35 dpc. Before challenge, the anti-PCV2 antibody in the group 3 piglets was significantly higher than that in group 2, ranging from 1:400 to 1:3200. The piglets in groups 1 and 4 remained free of PCV2-specific antibodies. After the challenge, antibody levels in groups 2 and 3 continued to rise and were the highest at 21 dpc. Groups 1 and 4 seroconverted at 14 dpc and showed significantly lower PCV2-specific antibodies than groups 2 and 3 until 35 dpc (p < 0.01). Figure 6b summarizes the PPV HI antibody titer trends. Before challenge, piglets in groups 1 and 3 had HI antibodies to PPV ranging from 1:128 to 1:256, but groups 2 and 4 seroconverted at 7 dpc, and their antibody levels were significantly lower than those in groups 1 and 3. This trend continued until 21 dpc. The antibody levels in groups 2 and 4 had risen to similar levels to those of group 3 by 28 dpc.
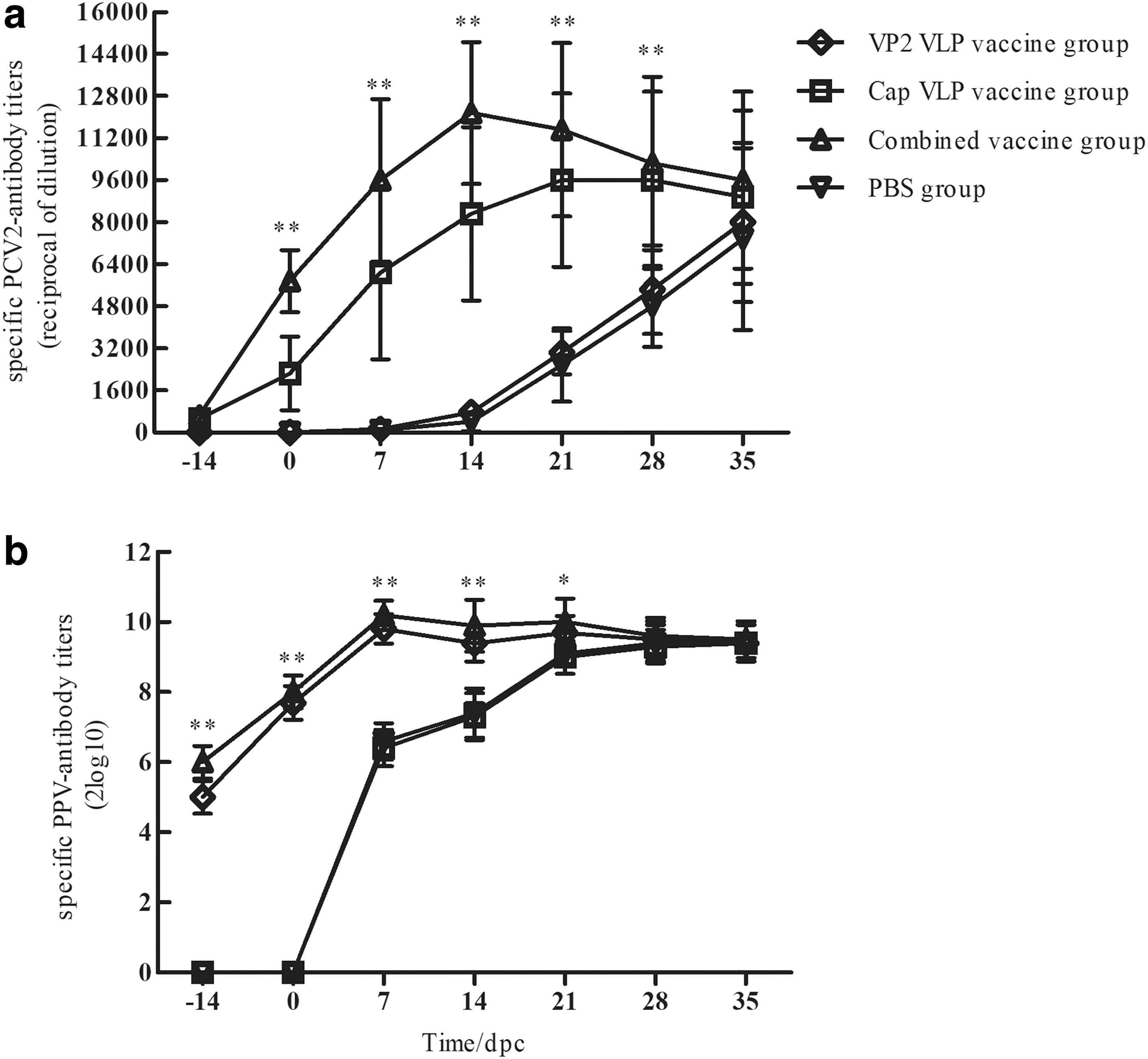
Serum antibody titers.
Figure 7 summarizes NA in serum samples. Before challenge, group 3 showed 1:12–1:16 NA levels against PCV2, which was significantly higher than that of the other groups (p < 0.01). After challenge, the NA levels dropped between 7 and 14 dpc, but returned to higher levels thereafter. Samples from groups 1 and 4 seroconverted at 21 dpc and continued to rise until reaching the highest level at 35 dpc. At 35 dpc, the NA levels did not significantly differ among the groups (p > 0.05; Fig. 7a).

Serum neutralizing antibody responses to PCV2
The NA titers against PPV were confirmed in ST cells. Before challenge, piglets in groups 1 and 4 had higher NA levels ranging from 1:160 to 1:320, which was significantly higher than that of the other groups (p < 0.01), but showed a small drop postchallenge. NA were detected at 14 dpc in groups 2 and 4, and the levels had risen to near those of groups 1 and 3 by 35 dpc (Fig. 7b).
Piglets in group 5 remained free of antibodies against PPV or PCV2 until the end of experiment at 35 dpc (data not shown).
Cytokine response
Cytokine mRNA expression (IL-8, TNF-α, IL-1β, IL-10, and IFN-γ) was evaluated in the piglets' PBMCs. Before challenge, the cytokine mRNA expression increased in the vaccinated groups (groups 1, 2, and 3), IFN-γ mRNA expression was higher than that in the control group (group 4; p < 0.05), and was highest in group 3. After challenge, the IL-8 and IL-1β levels were significantly increased in the vaccinated groups and higher than that in the control group at 14 and 21 dpc, but gradually decreased thereafter. In the control group, the mRNA expressions of IL-8 and IL-1β had risen to near those of the vaccinated groups by 28 dpc. TNF-α mRNA expression in the vaccinated groups increased and was higher than that in the control group at 21 and 28 dpc, especially in group 3, which was the highest of all groups. The control group showed no changes in TNF-α, which remained lower at all time points. The IFN-γ mRNA expression decreased progressively in all vaccinated groups, but remained higher than that in the control group until 28 dpc. IL-10 and IFN-γ were negatively correlated, and IL-10 mRNA expression increased faster in the control group than in the vaccinated groups, which was significantly higher than in the vaccinated groups at 14 and 21 dpc (p < 0.01). Groups 1 and 3 did not differ until the end of experiment (Fig. 8).
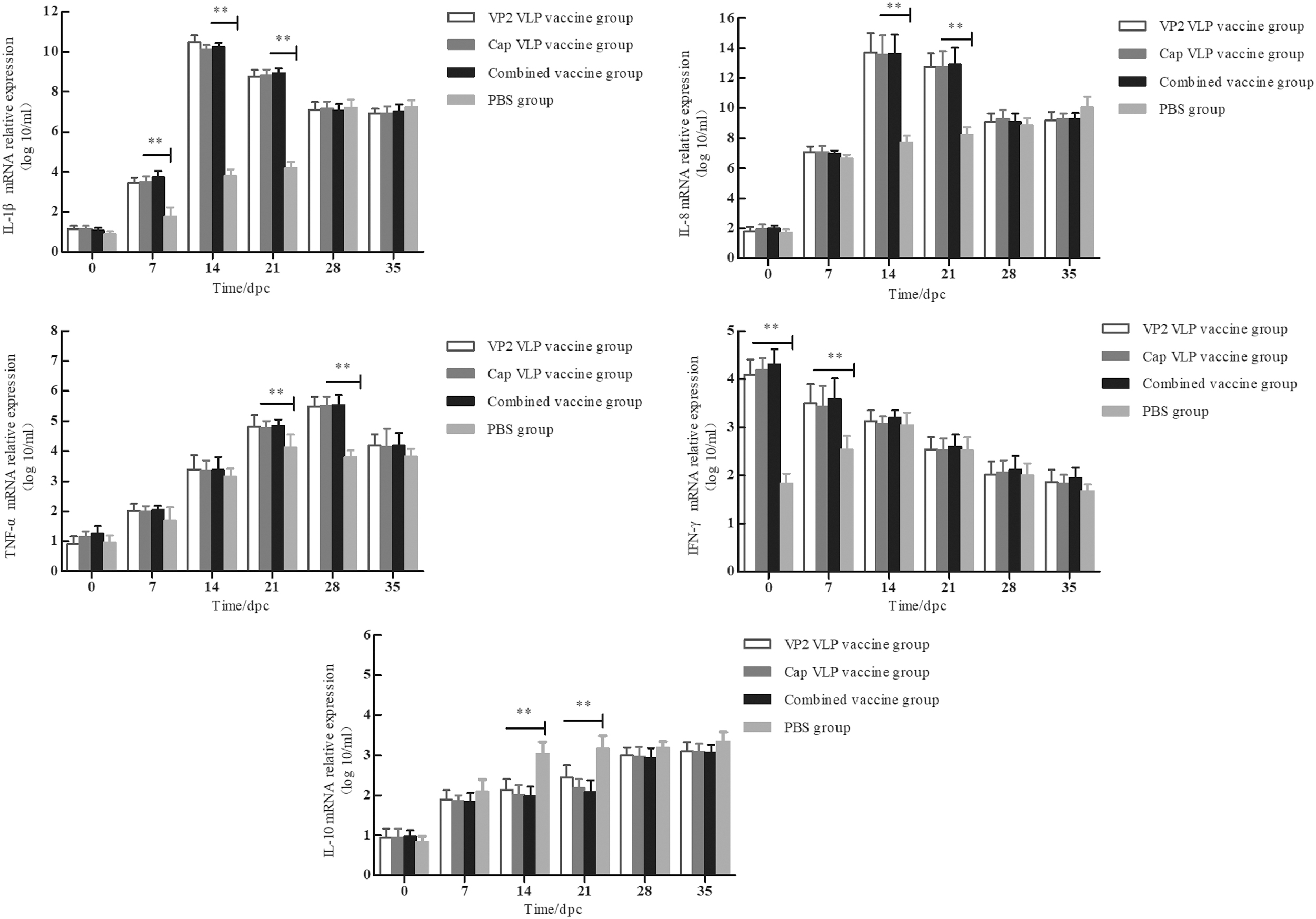
Mean group cytokine responses, including IL-1β, IL-8, TNF-α, IFN-γ, and IL-10, in PBMCs as determined by quantitative real-time PCR at 0, 7, 14, 21, 28, and 35 dpc. **p < 0.01 was considered highly significant. IL, interleukin; TNF, tumor necrosis factor; IFN, interferon; PBMC, peripheral blood mononuclear cell; PCR, polymerase chain reaction.
Cytokine mRNA expression (IL-8, TNF-α, IL-1β, IL-10, and IFN-γ) in the negative group remained low until the end of the experiment at 35 dpc (data not shown).
Determination of viral load in samples
Quantitative real-time PCR analysis of the serum showed that all piglets were PCV2-negative before challenge. Viremia was detected in all groups at 7 dpc, which continued to rise until 14 dpc in groups 2 and 3, but decreased thereafter and remained very low by 35 dpc, and the viremia in group 3 was significantly lower than that in group 2 at 14 and 21 dpc (p < 0.05). Moreover, the viremia in group 4 was always significantly higher than that in groups 2 and 3 (p < 0.01), but did not differ from that in group 1 (p > 0.05; Fig. 9a).
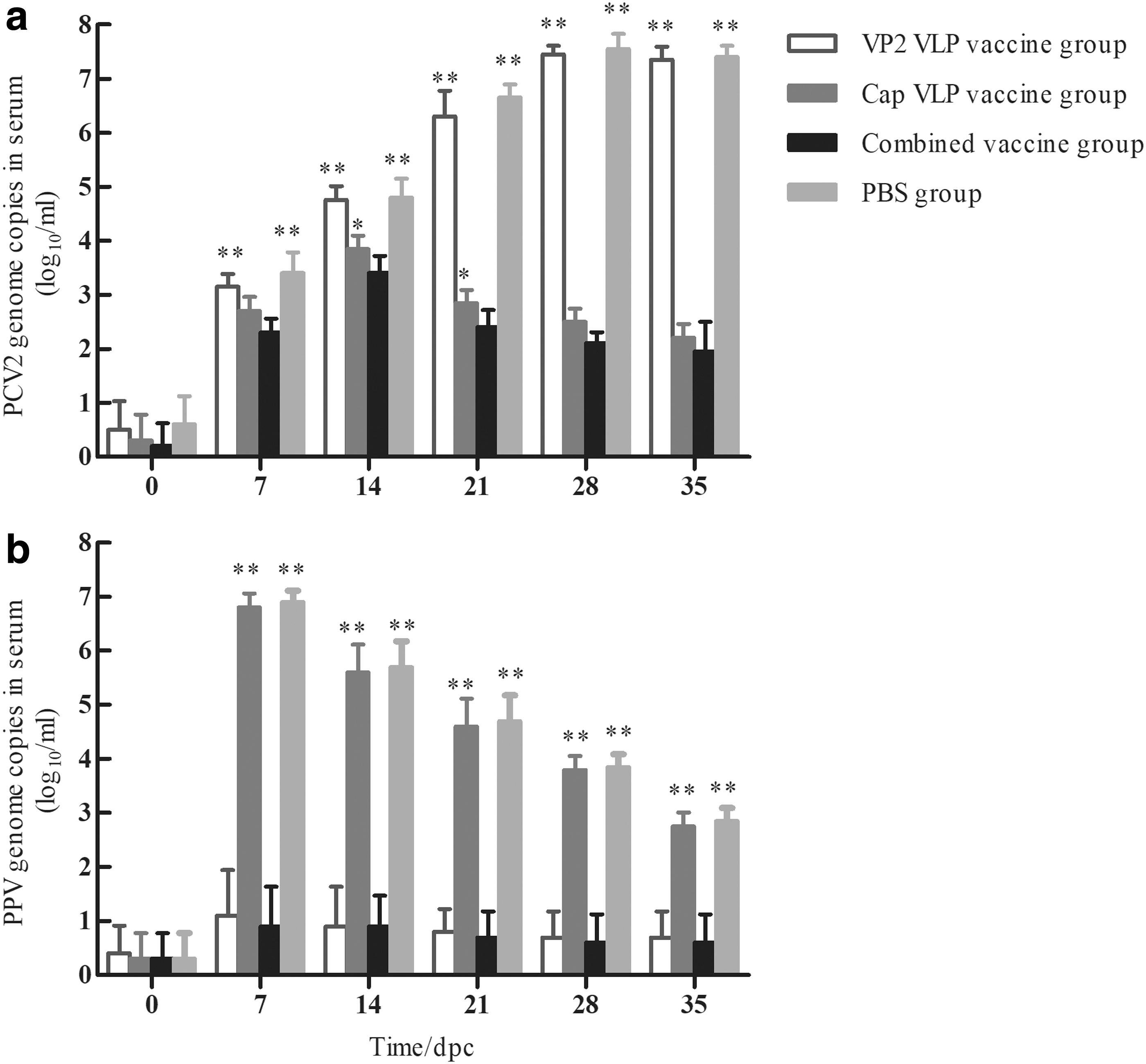
PCV2 viremia
PPV viremia was detected sporadically at low levels in groups 2 and 3 at each time points. In contrast, groups 1 and 4 piglets had substantial viremia from 7 dpc, which decreased thereafter and was detected in low levels at necropsy at 35 dpc (Fig. 9b).
In addition, inguinal lymph node (ILN) samples were collected in each group at 35 dpc, and the viral loads were detected. After challenge, the PCV2 viral loads in groups 1 and 4 were significantly higher than those in groups 2 and 3 at 35 dpc (p < 0.01), and the levels in group 3 were significantly lower than those in group 2 (p < 0.05; Fig. 10a).

PCV2 viral load
PPV viral loads in the ILNs were detected in groups 2 and 4 at 35 dpc, and the levels in group 4 were significantly higher than those in group 2 (p < 0.05), which was negative in groups 1 and 3 (Fig. 10b).
The piglets in group 5 remained free of PPV or PCV2 in the serum and ILNs until the end of the experiment at 35 dpc (data not shown).
Macroscopic and microscopic lesions
At necropsy, groups 3 and 5 had no remarkable gross lesions, and 4/10 animals in group 1 and 3/10 animals in group 2 had mildly enlarged lymph nodes. In contrast, most piglets in group 4 had severely enlarged lymph nodes, and severe edema of the lungs and kidneys, and 3/5 piglets had a few hemorrhagic spots in the spleen with a distinct serrated infarct at the edge (pictures not shown).
The relevant organs were collected at necropsy to prepare histopathological sections for evaluating the microscopic lesions (Fig. 11). Samples in group 3 and 5 were normal. However, mild-to-moderate microscopic lesions were observed in groups 1 and 2. Moreover, the pathological tissue changes in group 4 were more severe than those in groups 1 and 2. Severe diffuse interstitial pneumonia and alveolar edema occurred in the lungs (Fig. 11a). The liver showed severe water-like degeneration and vacuolated cytoplasm (Fig. 11b). The tonsils showed hyperchroma and lymphocyte infiltration (Fig. 11c). The heart sections showed lympho-histiocytic myocarditis and vacuolated cytoplasm (Fig. 11d) with a proliferation of red medulla with numerous necrotic cells and neutrophils and vague boundaries between each cell (Fig. 11e). Most piglets in group 4 had lymphoid depletion and histiocytic replacement of lymphoid follicles (Fig. 11f). Overall, the lesions in group 4 were consistent with PMWS and were significantly more severe than those in the vaccinated groups.
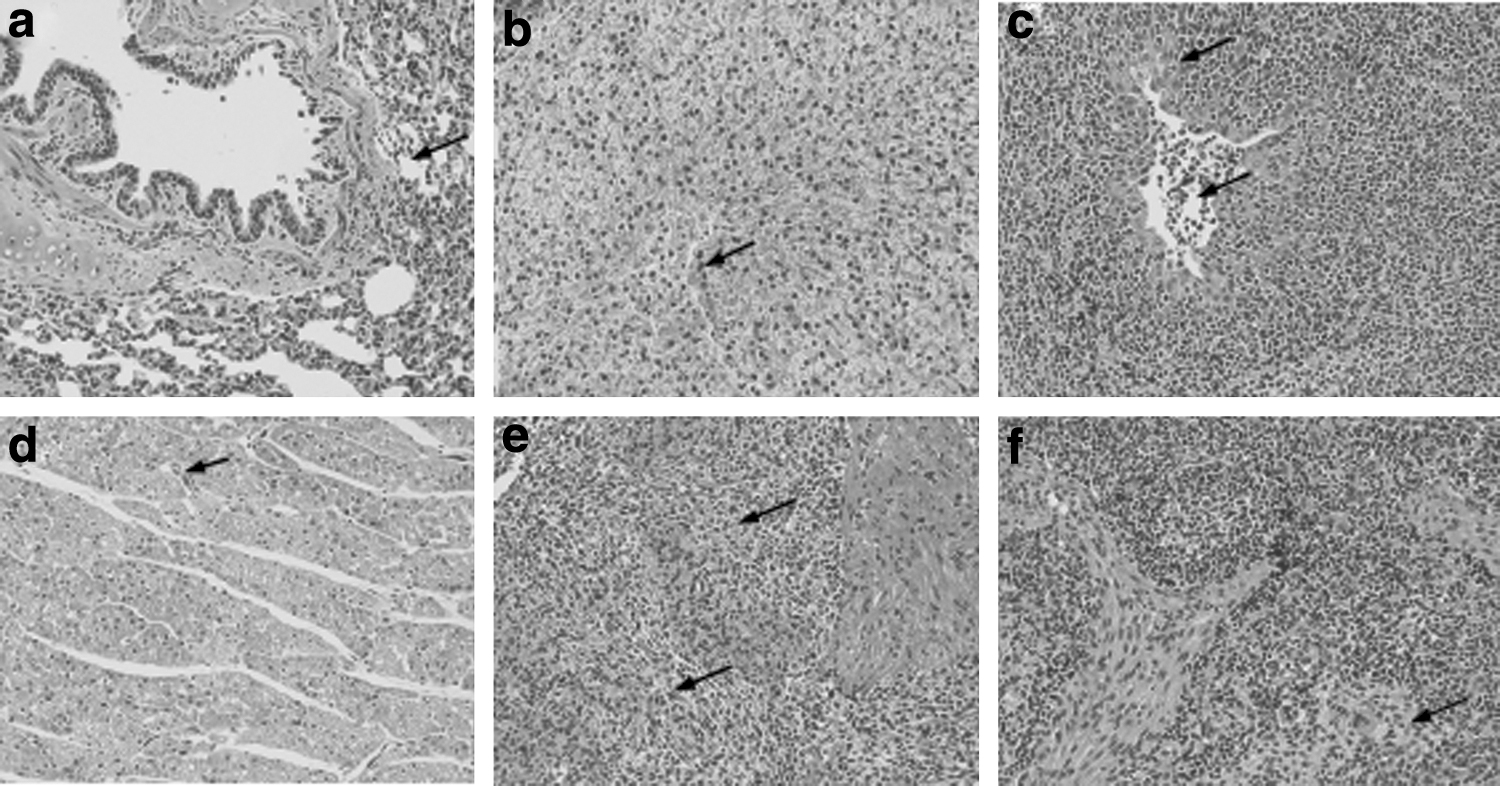
Microscopic lesions in the control piglets (arrows) at 35 dpc (hematoxylin and eosin stain, 200 × ).
Discussion
PMWS is a wasting disease caused by PCV2 infection. Since it was first reported in Canada in 1991, the disease has been found to be prevalent worldwide and has caused huge economic losses to the swine industry (30,31). Many experiments have confirmed that PPV enhances PCV2 infiltration and aggravates the clinical symptoms of PMWS. Currently, preventive measures for PMWS are vaccine immunizations (15). However, the PPV vaccine is only used in first-born sows in China and rarely used in piglets with the PCV2 vaccine. This poses risks for preventing PPV and PCV2 disease and is extremely unfavorable for controlling PMWS. Many studies have found that multiple immunizations cause a clinical stress response, which may increase the risk of PMWS (16,32). Therefore, a combined vaccine that can simultaneously prevent PPV and PCV2 should become the main research direction of preventing PMWS.
The VLP vaccine has been confirmed to highly biologically safe and is one of the most promising new vaccines. In this study, the PPV VP2 gene and PCV2 ORF2 gene nucleotide sequences were optimized for E. coli codon use. Recombinant proteins were overexpressed in BL21(DE3) cells with most of the VP2 proteins (67 kDa) and Cap proteins (27 kDa) present in the supernatant fraction on SDS-PAGE and western-blot analysis. In Tris buffer, purified proteins can self-assemble into VLPs, with diameters of ∼25–30 nm (VP2) and 20–25 nm (Cap) under TEM. Compared with other expression systems, the E. coli expression systems have short production cycles, are simple to scale-up, and are inexpensive (37). This study was designed to use VLPs emulsified with ISA201 adjuvant to prepare the vaccine. The VLP vaccine induced higher antibody levels against PPV and PCV2 in 3-week-old piglets; the VP2 and Cap VLPs did not antagonize each other when mixed, and titers of the combined vaccine were higher than those of the single vaccines. Piglets immunized with the VLP vaccine were coinfected with PPV and PCV2, and the combined vaccine immunization significantly reduced the viral load levels of PCV2 and PPV in the serum and tissue compared with those of the single vaccine, and more importantly eliminated clinical disease. All piglets in the combined vaccine group showed no typical pathological changes of PMWS either macroscopically or microscopically. However, most piglets in the control group showed severe PMWS symptoms at different time points postchallenge. In addition, the single vaccine with VP2 and Cap VLPs designed in the study partially reduced the incidence in piglets, but did not completely prevent PCV2 and PPV infection, correspondingly. Particularly, the single VP2 VLP vaccine only minimally protected against PMWS. In addition, trial 2 was designed to compare the immune efficacy of the VLP vaccine in a PMWS-infected farm. Daily weight gain was chosen as the evaluation index of piglet growth performance. The results showed that the daily weight gains of the combined vaccine group were significantly higher than those of the Cap VLP vaccine group and the negative group, which reflected the application value of the combined vaccine.
Evaluating immune mediators, such as cytokines and related lymphocyte subpopulations involved in the immune response, can contribute to understanding the efficiency of host immune responses to vaccines and the subsequent immunoprotection. Innate proinflammatory (TNF-α, IL-1β, IL-8) and adaptive immune (IFN-γ, IL-10) cytokines are thought to be related to the viral load and the presence of clinical signs of PMWS (13,25). IFN-γ is usually produced by host cells during adaptive response to a vaccine or virus and can effectively resist viral replication and activate NK cells to clear the virus (11,23). IFN-γ expression increased rapidly, and the highest level occurred at 7 dpc in the PBS group, but gradually decreased thereafter, which was consistent with the gradual increase in viral load at 14 dpc. The combined vaccine groups presented a stronger cellular immune response and higher IFN-γ expression levels, which allowed the virus to be eliminated rapidly in the early stages of infection. The results of the IFN-γ expression show the positive effect of vaccinations on cellular immunity and account for the reduced viral replication. In addition, macrophages were stimulated to secrete IL-8 and IL-1β, which are related to inflammatory response and viral clearance. Their expressions are upregulated when the viral amount is low, and lesions are slight at the early phases of infection. The IL-8 and IL-1β expression levels could reveal the piglets' ability to react against viral infection through effective inflammation (7). In this study, IL-8 and IL-1β expression levels were significantly increased in the vaccinated group compared with those of the PBS group in the early phase (7–21 dpc) of infection, which remained higher in the combined vaccine group at 28 dpc. Conversely, IL-8 and IL-1β expression levels were lower in the PBS group during infection, which may explain the severe clinical symptoms and pathological injuries and was consistent with other authors' results (22,26).
TNF-α is closely related to viral replication during infection. In this study, TNF-α expression levels gradually increased after challenge in the vaccinated groups, especially in the combined vaccine group, and remained relatively high until 28 dpc. In contrast, TNF-α gradually increased in the early phases of infection and played an antiviral role, but decreased in the later phases, which also verified the persistence of PCV2 viremia in the single vaccine group and PBS group.
IL-10 is a regulatory factor and is considered to be an immunosuppressive and anti-inflammatory mediator. It inhibits macrophage function and synthesis of some cytokines (6,8). PPV and PCV2 can stimulate IL-10 release from PBMCs and tissues (34), which is associated with developing severe PMWS (9,10). In this study, IL-10 and IFN-γ showed opposing levels in the early phase of infection, and piglets in the PBS group showed significantly higher IL-10 levels than did the vaccinated groups, which may play a role in PMWS onset. Increased IL-10 expression levels in animals with severe disease may be associated with low levels of proinflammatory cytokines and IFN-γ, which also explains the significant immunosuppression that occurs in animals infected with PMWS (17).
In this study, the piglets immunized with the combined VLP vaccine showed strong humoral and cellular responses. The results confirmed that the two VLPs designed in this study did not antagonize each other. The combined vaccine protected piglets without PMWS symptoms and improved their growth indices. The new combined vaccine has good clinical applications and is ideal for developing the combined vaccine.
Footnotes
Disclaimer
All animal experiments were performed in accordance with protocols approved by the Institutional Animal Use Committee.
Author Disclosure Statement
No competing financial interests exist.