
Abstract
Salmonella is among the most serious of foodborne pathogens worldwide and distributed widely in the natural environment; in addition, it has caused severe medical problems and foodborne diseases. Bacterial biofilm was the multicellular community of microorganisms that attached to nonbiological and biological surfaces. Phages and their derivatives are ideal candidates for replacing and compensating antibiotic resistance problems in the future. In this study, a virulent phage of KM15 was isolated from pig slaughterhouse sump samples in Kunming, China. It belonged to the Siphoviridae family, and optimal growth temperature was 42°C, the pH of optimal preservation buffer was 6–7, optimal multiplicity of infection was 0.0001, and the genome size was 41,869 bp. The Salmonella paratyphi A and Salmonella paratyphi B have a broad spectrum of antibiotic resistance and were isolated from clinical patients in the First People's Hospital of Yunnan Province; fortunately, most of them can be lysed by phage KM15. Collaboration of phage KM15 and kanamycin sulfate has a better antibiofilm effect than KM15 and kanamycin sulfate alone, in low-concentration bacterial culture; KM15 has better antibiofilm effect than kanamycin sulfate in high-concentration bacterial culture. The data of this study provided a strong evidence of application of phage to reduce the growth of Salmonella biofilm, which was important for public health.
Introduction
S
Bacterial biofilm was the multicellular community of microorganisms that attached to highly hydrated extracellular matrices and biological and nonbiological surfaces (3,13,18). The extracellular polymeric substance matrix of biofilm acts as a barrier to reduces the penetration of microbiotic and antimicrobial agents into the interior of the biofilm (11,29). Biofilms of microorganisms were highly resistant to acidic condition, antibiotics, heat, and desiccation (9). Bacteria with biofilm were ∼10 1,000 times less sensitive to antimicrobial agents than planktonic bacteria for the reason that extracellular polymeric substances of the biofilm prevent the entrance of antimicrobial agents (27,33). Total elimination of biofilm among husbandry, the food industry, and in clinics is scarcely possible (23).
The extensive use of antibiotics resulted in the problem of multiple resistant bacteria (MRB). According to Ministry of Health, People's Republic of China, data published in 2003, 80,000 people in China die each from overuse of antibiotics (5). People's Republic of China Consumption of Salmonella-contaminated food leads to fever, abdominal pain, and diarrhea after 12–72 h (5). Salmonella is one of the most serious pathogens in fresh produce owing to its strong survival and persistence in the environment (35). The clinical and food industry faces a serious Salmonella contamination problem due to the overuse of antibiotics, resulting in a large number of multiple resistant, clinical, and foodborne Salmonella strains. New ways to control Salmonella contamination are badly needed (20).
The applications of new antibacterial agents and disinfectants to eradicate MRB Salmonella and Salmonella biofilms are urgent needed. Phage and its derivatives can be used as a feasible, novel, and safe biological treatment, elimination, and prevention of Salmonella in clinical and food processing environments and food (7,12,17,24). Above all, phages are harmless for humans, making them safe for applying in food products and clinic (30). A recent study has found that phage has a high efficiency in reducing and control of bacterial biofilms formed on various surfaces of Listeria monocytogenes, Pseudomonas aeruginosa, Escherichia coli, and Salmonella (1,6,14,15,22).
Prevention and control of contamination caused by multidrug-resistant (MDR) bacteria of Salmonella are of great importance for the food and medical industries. The main challenge for phage therapy was the relatively narrow lytic spectrum for host bacteria. Here we report a new isolated lytic phage named KM15, which has better antibiofilm effect than antibiotics in high concentrations of bacterial culture. Therefore, the purpose of this study was to reduce biofilm formation and control mature biofilm of Salmonella by use of phage KM15. Our aim was to determine whether KM15 could be used as an alternative therapeutic agent against MDR Salmonella strains.
Materials and Methods
Bacterial strains and growth conditions
Salmonella paratyphi A and Salmonella paratyphi B strains were isolated from the patients in the First People's Hospital of Yunnan Province, China; Salmonella paratyphi A NA3 was used as host for phage isolation. Salmonella was grown aerobically on Luria-Bertani plate culture or in LB broth culture (Difco, Detroit, MI), incubated at 37°C. Semisolid medium containing of 0.5% agar (m/m) in LB broth culture for phage plaque confirmation and solid medium containing of 1.8% agar (m/m) in LB broth culture were used. All Salmonella strains were stored at −80°C, containing 20% (V/V) glycerol.
Phage isolation and purification
Phage was isolated from pig slaughterhouse sump samples collected from Kunming City, China, in September 2018. The isolation method for phage was according to the following methods with brief modification (32). Briefly, 10 g of slaughterhouse sump sample was mixed with 20 mL sterile normal saline (0.9% NaCl) buffered in a sterile 50 mL tube and then shock cultured for 2 h, 200 rpm at room temperature. Then centrifuged at 5,000 g for 15 min and filtered with a 0.22 μm filter membrane. Ten milliliters of filtered medium was added to 30 mL of LB broth culture containing 0.5 mL Salmonella paratyphi A NA3, culture overnight and incubated for 48 h. After that, cocultures were centrifuged at 8,000 g for 15 min and the supernatant was filtered with a 0.22 μm filter membrane. The filtrate was series diluted 10 times, mixing with 5 mL of 0.5% agar (m/m) molten LB containing Salmonella paratyphi A NA3 (2 × 108 colony-forming unit [cfu]/mL), and immediately added to an LB solid medium plate containing 1.8% agar (m/m). The plaque was observed after overnight culture. Single plaque was selected for phage purification and repeated three times.
The pH, thermotolerance, multiplicity of infection, growth curve, and transmission electron microscopy of isolated phage
The phage stock was diluted to 1 × 108 plaque-forming unit (pfu)/mL with LB broth culture. Put the solution buffer with pH of 3, 4, 5, 6, 7, 8, 9, 10 and 11 into each 1.5 mL sterile centrifuge tube, following added of diluted phage with the titer of 1 × 108 pfu/mL to each tube. Solution buffer of pH 3, pH 4 and pH 5 were prepared by 50 mmol/L citrate buffer, solution buffer of pH 6, pH 7 and pH 8 were prepared by 50 mmol/L phosphate buffer, solution buffer of pH 9 was prepared by 50 mmol/L Tris-HCl buffer, solution buffer of pH 10 and pH 11 were prepared by 50 mmol/L sodium carbonate buffer. Place at room temperature for 1 h to detect the titer of phage in different pH buffers. The experiments were repeated three times. Place 1 mL of diluted phage in a temperature controlled at 4°C, 25°C, 37°C, 42°C, 50°C, 60°C, and 90°C for 1 h, respectively, for thermotolerance detection. Multiplicity of infection (MOI) was the ratio of phage to host bacteria of initial infection. Added phage KM15 stocks in Salmonella paratyphi A NA3 culture medium that according to the MOI of 0.0001, 0.001, 0.01, 0.1 1, 10 and 100, then cultured at 37°C for 8 h. The cultures were centrifuged at 10,000 g for 15 min at 4°C, then the supernatant was filtered with a 0.22 μm filter, and the titer of phage increment solution was determined through the double plate method; the experiment was repeated three times. For growth curve measure, 1 × 108 pfu/mL of phage KM15 was added to LB culture containing 1/250 Salmonella paratyphi A NA3 seed culture according to the optimum MOI and shake culture at 37°C, intermittent sampling was used to determine the titer of phage.
Transmission electron microscopy
The morphology of phage KM15 particles was analyzed through transmission electron microscopy. Briefly, phage stock dilution (titer approximately at 2 × 108–2 × 109 pfu/mL) was deposited on copper grids with carbon-coated formvar films, stained with 2% uranyl-acetate (pH 4.0). Phage samples were imaged using a Philips EM 300 electron microscope, operated at 80 kV in Jiangnan University (Wuxi, China). Phage KM15 was classified and identified according to the International Committee on Taxonomy of Viruses (34).
The extraction, sequencing, and bioinformatic analysis of KM15 genome
First, phage KM15 was purified with a 10 kDa filter (titer of 109–1010 pfu/mL). Briefly, 10 mL of phage culture solution was taken, sterilized by a 0.22 μm filter, added to 15 mL of filter of 10 kDa, then centrifuged at 4,000 g for 1 h until about 200 μL was left in the filter, and then transferred to a new sterile centrifuge tube with pipette for subsequent testing. Purified phage KM15 was treated with RNase and DNase at 37°C for 1 h. Then, the Takara MiniBEST Viral RNA/DNA Extraction Kit (Cat#9766) was used for extraction of KM15 genomic DNA. Restriction endonuclease of NotI, EcorI, XhoI, and HindIII was used for phage genome digestion. Extracted KM15 genome DNA was sequenced through Illumina HiSeq (Sangon Biotech, China). The original sequencing data were evaluated by FastQC and assembled with SPAdes assembler software. The National Center for Biotechnology Information Blast compared the multiple databases of CDD, KOG, COG, NR, NT, PFAM, SwissProt, and TrEMBL, which were used for function annotation information of KM15 genome gene and protein sequence.
Lytic spectrum of phage KM15
The lytic spectrum of phage KM15 was determined through spot test method (8). The reference strains tested for phage KM15 susceptibility were the 2 strains of Salmonella paratyphi B and 10 strains of Salmonella paratyphi A. Generally, 200 μL of host strains (109 cfu/mL) was added to 5 mL LB soft agar (LB broth culture containing 0.5% agar [m/m]), and poured on LB solid plate containing 1.8% agar (m/m). A few minutes later, 10 μL of phage suspension was added and incubated at 37°C for 24 h.
Antimicrobial susceptibility testing
Antibiotic sensitivity of the Salmonella paratyphi B and Salmonella paratyphi A was tested for 17 antibiotics by the minimal inhibitory concentration method. Briefly, taken of 2 mL agent for tested, filtered into the plastic pinkie tube with a microporous membrane, multiple dilution and added to prepared medium for test. Then, 0.05 mL of various bacterial solutions with a concentration of 105–106 cfu/mL was added with a graduated dropper. Incubated at 37°C with 200 rpm for 18 h, then observed the clarity of the tested tube. The tested antimicrobials were gentamicin (GM), cefoxitin (FOX), imipenem (IMI), piperacillin (TZP), amoxicillin (AMC), selectrin (SXT), ciprofloxacin (CIP), ceftriaxone (CTR), tigecycline (TGC), amikacin (AN), ampicillin (AMP), levofloxacin (LEV), cefepime (FEP), aztreonam (AZM), tobramycin (TOB), macrodantin (FTN), and ertapenem (ETP).
The different effects of KM15 and kanamycin sulfate for biofilm scanning electron microscopy
First, 48-well cell slides were placed into a 24-well plate. Seed solution of Salmonella paratyphi A NA3 was inoculated into 100 mL of LB culture with a rate of 4‰. Then, 1 mL Salmonella culture were added in each hole of 24 well plate, at the same time, phage KM15, kanamycin sulfate, mixtures of phage KM15 and kanamycin sulfate were added, added of phosphate buffer (PBS, pH 7.2∼7.4) as mock, the final concentration of kanamycin sulfate was 10 μm/mL and the amount of added phage refer the MOI of 0.1. Following cultured at 37°C for 24 h. For other groups, first, seed solution of Salmonella paratyphi A NA3 was inoculated into 100 mL of LB culture with a rate of 4‰ and cultured for 12 h. Then, 1 mL Salmonella culture were added in each hole of 24 well plate, at the same time, phage KM15, kanamycin sulfate, mixtures of phage KM15 and kanamycin sulfate were added, added of phosphate buffer (PBS, pH 7.2∼7.4) as mock, the final concentration of kanamycin sulfate was 10 μm/mL and the amount of added phage refer the MOI of 0.1. Following cultured at 37°C for 24 h. The plate counting method was used for cfu measure. The recovered culture was washed twice with PBS buffer and fixed with 2.5% (v/v) precooling glutaraldehyde at room temperature for 3 h in a dark place. After that, washed twice with PBS buffer and dehydrated with an increasing ethyl alcohol gradient of 15%, 30%, 50%, 70%, and 100% (v/v) for 10 min with each step. Finally, dry by leaving overnight and gilt in timely. Scanning electron microscopy (SEM) of Salmonella paratyphi A NA3 was observed through a scanning electron microscope with an accelerating voltage of 20 kV.
Biofilm formation ability detected through microplate reader
Seed solution of Salmonella paratyphi A NA3 was inoculated in LB culture with a ratio of 4‰ and cultured overnight. Then diluted 200 times with LB culture and added into a 96-well plate (200 μL/hole), repeated three times. Phage, kanamycin sulfate, and mixtures of phage and kanamycin sulfate were added, respectively, and phosphate buffer added as mock (the final concentration of kanamycin sulfate was 10 μM/mL and added phages with reference of MOI = 0.1). Following cultured at 37°C for 24 h. For other groups, first, the seed solution of Salmonella paratyphi A NA3 was inoculated in 100 mL of LB culture with a rate of 4‰ and cultured for 12 h, and then inoculated in 1 mL Salmonella culture in a 96-well plate; phage, kanamycin sulfate, and mixtures of phage and kanamycin sulfate were added, respectively, and phosphate buffer (PBS) was added as mock (the final concentration of kanamycin sulfate was 10 μM/mL and added phages with reference of MOI = 0.1). Cultured at 37°C for 24 h. The OD600 of culture was measured using an ELISA (Thermo Scientific; EUA). Then discarded of bacteria culture and washed twice with PBS for remove of unattached cells, fixed with 99% methanol for 15 min, following discard methanol and dried at room temperature, stained with 2% crystal violet for 8 min. Washed culture plate with running water until the water was colorless. Absorption light at wavelength of 570 nm was measured with a microplate reader after drying. The experiment was repeated three times.
Results
Characteristics and morphology of isolated phages
A virulent phage, KM15, was isolated from pig slaughterhouse sump samples. The plaque of phage KM15 was 2 mm in diameter after overnight incubation at 37°C (Fig. 1A). Negatively stained purified phage KM15 was observed with an electron microscope. Transmission electron microscopy revealed that KM15 virion has an icosahedral head of 52 ± 2 nm diameter and possesses a noncontractile tail 140 ± 5 nm long (Fig. 1B). The morphology of phage KM15 indicated that it belonged to the Siphoviridae family. The growth curve of phage KM15 was obtained by inoculation on Salmonella paratyphi A NA3 according to MOI of 0.1 at 37°C (Fig. 2A). The latent period of phage KM15 was 60 min. The titers of phage KM15 reached peaks very quickly in 170 min and appeared going down 15 h later. The amplification factor of phage KM15 was ∼200 times.
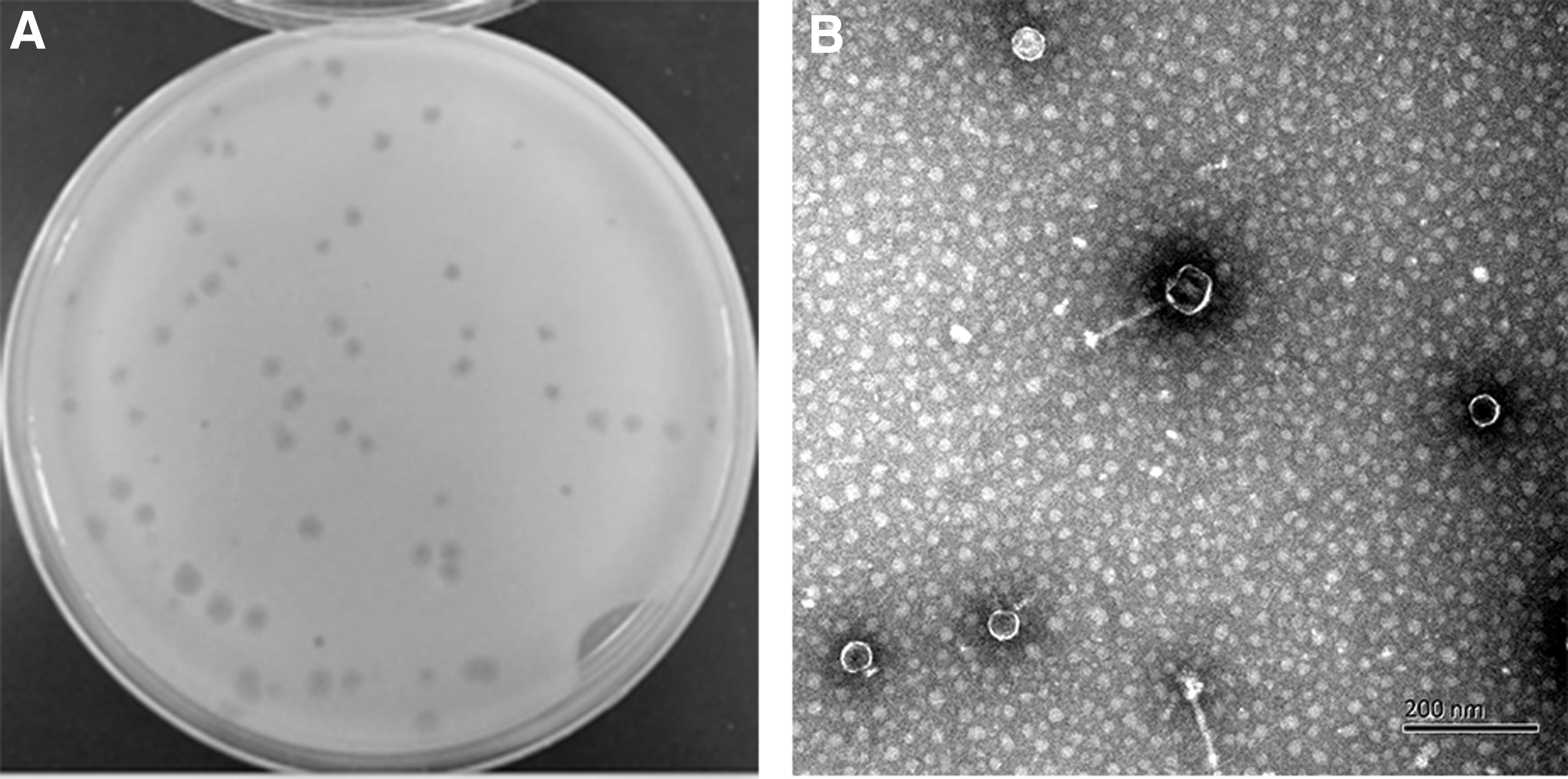

Optimum temperature, pH, and MOI of isolated phage KM15
KM15 has the highest activity after being preserved at 42°C for 1 h, then the activity of KM15 had a noticeable decline at 50°C and complete inactivation until 90°C (Fig. 2B). The result indicated that phage KM15 has low temperature adaptability, which consistent with the optimum survival temperature of Salmonella paratyphi A NA3. Phage KM15 has the most plaque at pH = 6–7, the activity was higher in buffer of pH = 10–11 than pH = 3–4 (Fig. 2C). These results indicated that phage KM15 has good tolerance for alkaline culture. MOI refers to the ratio of the number of phage to host bacteria. The optimum MOI of phage KM15 was 0.0001, the infection ability of KM15 decreased significantly after MOI of 0.001 and reached minimum at MOI of 100 (Fig. 2D).
Characteristic and analysis of genome
The genome size of phage KM15 was 41,869 bp. We have identified 64 protein-coding genes (open reading frames for KM15). Phage KM15 endolysin was located at 5,023–5,511 bp with size of 489 bp in genome. There were three tail gene location positions of 16,000, 18,500, and 30,000 bp among genomes. The G and C nucleotide composition of phage KM15 was 49.96%. There were 20 genes transcribed in reverse direction and 44 genes transcribed in forward direction among the genomes. Phage KM15 DNA was digested by EcoRI but could not be digested by XhoI, NotI, and HindIII. This was consistent with the results of genome analysis of phage KM15. Genome and experiment result analysis revealed that KM15 was a virulent phage (Figs. 3 and 4).
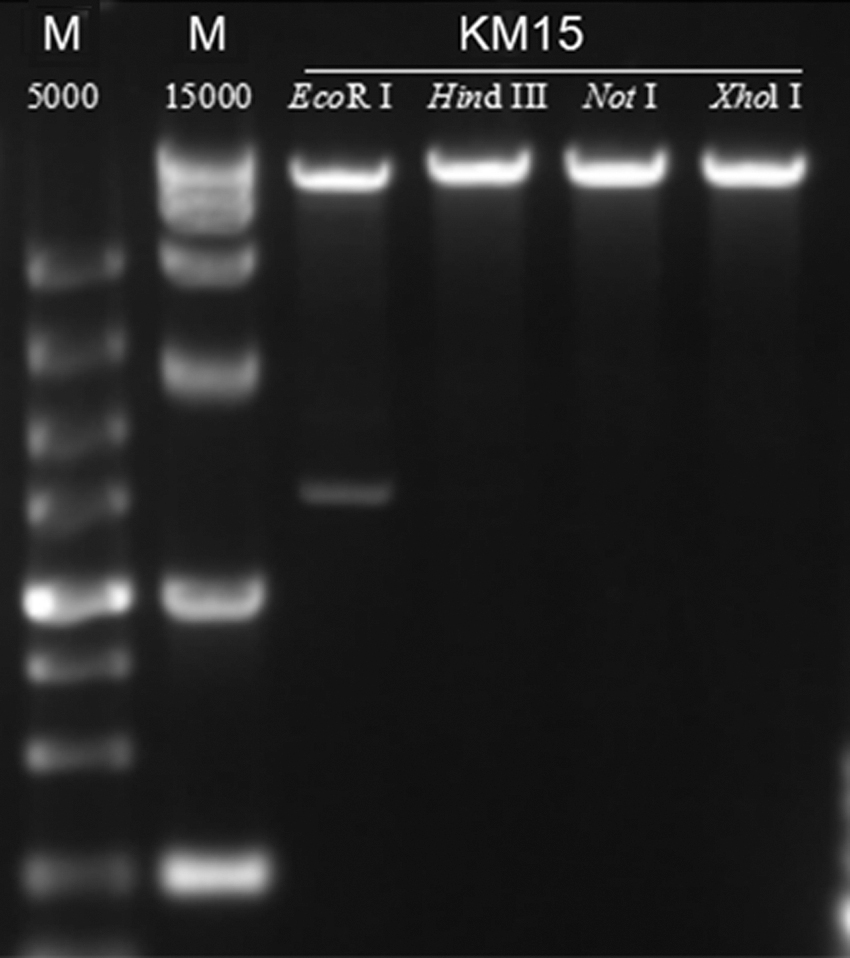
Restriction enzyme digests of phage KM15. Phage KM15 DNA was digested by EcoRI (lane 3), whereas it could not be digested by HindIII, NotI, and XhoI.
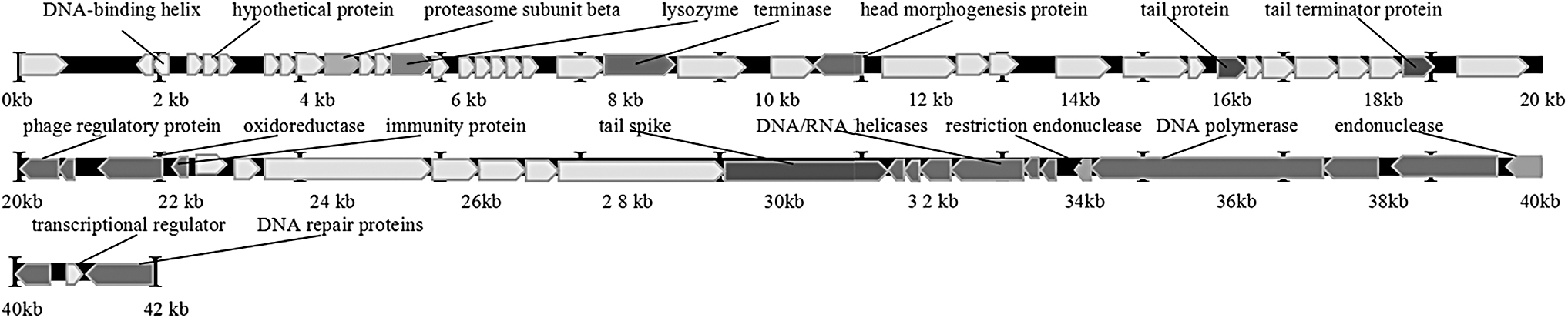
Line map of the phage KM15 genome. In the phage KM15 genome track, lysozyme gene was located at 5 k bp to 5.5 k bp and the arrows represent the transcription direction of open reading frames.
Antimicrobial susceptibility and phage lytic spectrum
Salmonella paratyphi B and Salmonella paratyphi A strains were isolated from clinical patients in the First People's Hospital of Yunnan Province. Upsetting, they were MDR (Table 1). Phage KM15 possesses lytic for many strains of Salmonella paratyphi. The strains of Salmonella paratyphi A JJ9 and Salmonella paratyphi A NA3 exhibited sensitivity to KM15 (Table 2). All strains of Salmonella paratyphi possess resistance to tobramycin, ciprofloxacin, gentamicin, cefoxitin, streptomycin, and penicillin, but are sensitive to macrodantin and kanamycin sulfate.
Antibiotic Resistance of Salmonella Isolates Used in This Study
R, resistant; S, sensitive.
Host Range Analysis of Phages KM15
Compare the antibiofilm effects of phage KM15 and kanamycin sulfate for Salmonella
SEM was used for observing Salmonella paratyphi A NA3 biofilm on round coverslips, which was affected by kanamycin sulfate (10 μg/mL) and phage KM15 (MOI = 0.1). For the group of Salmonella paratyphi A NA3 was inoculated for rate of 4‰, following added kanamycin sulfate (10 μg/mL) and KM15 (MOI=0.1) and culture for 24 hours, kanamycin sulfate has better sterilization effect than phage KM15 among the result of co-culture OD600, scanning electron micrograph and microplate reader OD570 of Salmonella paratyphi A NA3 co-culture biofilm (Figs. 5 and 6). Whereas for the condition of Salmonella paratyphi A NA3 inoculation at the rate of 4‰ and cultured for 12 h, kanamycin sulfate (10 μg/mL) and KM15 (MOI = 0.1) were added and cultured for 12 h; phage KM15 had a better sterilization effect than kanamycin sulfate (10 μg/mL) in the result of OD600 measured coculture, scanning electron micrograph of Salmonella paratyphi A NA3, and microplate reader at OD570 for detector coculture biofilm (Figs. 5 and 6). Moreover, for the condition of Salmonella paratyphi A NA3 inoculation at the rate of 4‰ and cultured for 12 h, then kanamycin sulfate (10 μg/mL) and KM15 (MOI = 0.1) were added and cultured for 12 h, collaboration of phage KM15 and kanamycin sulfate had a better sterilization effect than kanamycin sulfate or KM15 alone, whether in the condition of OD600 measured coculture, scanning electron micrograph of Salmonella paratyphi A NA3, or microplate reader at OD570 for detector coculture biofilm (Figs. 5 and 6).
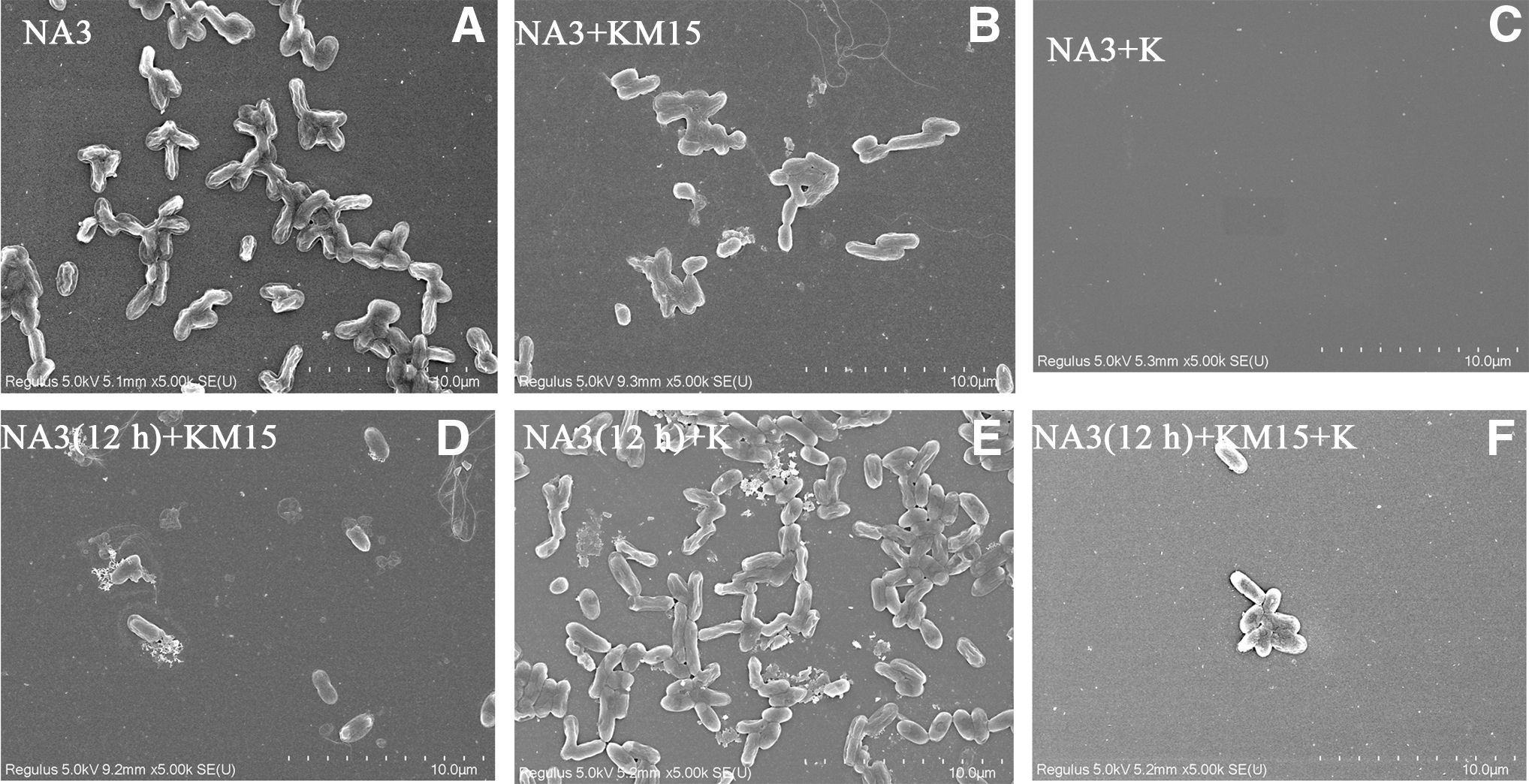
Scanning electron micrograph of Salmonella paratyphi A NA3 biofilm formed on round coverslip.

Effects of phage and kanamycin sulfate (10 μg/mL) on biofilms.
The result of host colony-forming unit indicated that collaboration of phage KM15 and kanamycin sulfate had a better sterilization effect than KM15 or kanamycin sulfate alone (Fig. 7). For another where Salmonella paratyphi A NA3 inoculation at the rate of 4‰ and cultured for 12 h, and then kanamycin sulfate (10 μg/mL) and KM15 (MOI = 0.1) were added and cultured for 12 h, phage KM15 had a better sterilization effect than kanamycin sulfate (Fig. 7).

Effects of phage and kanamycin sulfate (10 μg/mL) on colony-forming unit of Salmonella paratyphi A NA3.
Discussion
Salmonella was a common foodborne pathogen for severe medical problems mainly found in contaminated food (5,31). Salmonella paratyphi B and Salmonella paratyphi A strains used in this study were isolated from clinical patients who accidentally ate Salmonella-contaminated bread in the First People's Hospital of Yunnan Province, China. Overuse of antibiotics increased the severity of Salmonella (25). The main problem of drug resistance was that antibiotic resistance and MDR genes have recombined in the genome of almost all Salmonella strains (26).
The isolated Salmonella phage KM15 belongs to the Siphoviridae family and the genome sizes were 41,869 bp. Corresponding genome sizes of Salmonella phage SPFM was 233–242 kb and Salmonella phage ΦStp1 was 112,149 bp. They all belong to the family of Siphoviridae (38,39).
They have many multiple resistant bacteria and superbacteria with the rising abuse of antibiotics which is leading to serious public health problems. Alternatives for antibiotics were urgently needed. The isolated Salmonella phage KM15 can be an ideal substitute for antibiotics for it has the ability of lytic MDR strains of S. paratyphi Salmonella formation biofilm on different food surfaces, increasing the risk of cross-contamination, which was a serious problem for public health and food industries among poultry products (2,16,36). The exploiting of effective eliminate on biofilm was still challenging in pathogen control (4). Until now there has been no ideal technology for biofilm control, indicating that new control strategies for biofilm are badly needed (37). Here, we have demonstrated the ability of the phage KM15 and kanamycin sulfate for reduced biofilm formation of Salmonella.
The result shows that kanamycin sulfate and phage KM15 can control the formation of Salmonella biofilm, indicating that phage KM15 has the potential for reducing S. paratyphi biofilm. Kanamycin sulfate has a better antibiofilm effect than phage KM15 in low concentrations of bacterial culture (Fig. 5–7). Nonetheless, phage KM15 has a better antibiofilm effect than kanamycin sulfate in high concentrations of bacterial culture, and a collaboration of kanamycin sulfate and phage KM15 has a better antibiofilm effect than KM15 or kanamycin sulfate alone in low and high concentrations of bacteriak culture (Figs. 5–7). In conclusion, we have recovered a new Salmonella phage KM15, which exhibited activity to infect and kill multiple antibiotic tolerances of clinical Salmonella strains. Administration of phage KM15 was effective to reduce the growth and biofilm of Salmonella. These findings could expand our insight on Salmonella phage and offer new potential therapeutic alternatives for clinic and public health.
Footnotes
Ethics Statement
Ethical approval was obtained from the Institutional Ethics Committee (The First People's Hospital of Yunnan Province, Kunming, Yunnan, China). The study protocol was in accordance with the Declaration of Helsinki for Human Research of 1974 (last modified in 2000). Written informed consent was received from each patient before sample collection.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by Yunnan Science and Technology Commission from Yunnan provincial Science and Technology Department and Kunming Medical University, grant number 2018FE001(-115), Health commission of Yunnan province talent program, grant number H-2017027. Sponsored by K.C. Wong Magna Fund in Ningbo University.
The authors gratefully acknowledge the financial support provided by the National Key Research and Development Program of China (2017YFC1601704), Projects 31522044, 31671909, and 31772034 of the National Natural Science Foundation of P.R. China and Program of Jiangsu Key Laboratory of Advanced Food Manufacturing Equipment and Technology (FMZ201904), National First-class Discipline Program of Food Science and Technology (JUFSTR20180205).