
Abstract
Since the end of 2019, the emergence of novel coronavirus disease 2019 (COVID-19) caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has accelerated the research on host immune responses toward the coronaviruses. When there is no approved drug or vaccine to use against these culprits, host immunity is the major strategy to fight such infections. Type I interferons are an integral part of the host innate immune system and define one of the first lines of innate immune defense against viral infections. The in vitro antiviral role of type I IFNs against Middle East respiratory syndrome coronavirus (MERS-CoV) and SARS-CoV (severe acute respiratory syndrome coronavirus) is well established. Moreover, the involvement of type I IFNs in disease pathology has also been reported. In this study, we have reviewed the protective and the immunopathogenic role of type I IFNs in the pathogenesis of MERS-CoV, SARS-CoV, and SARS-CoV-2. This review will also enlighten the potential implications of type I IFNs for the treatment of COVID-19 when used in combination with IFN-γ.
Introduction
Type I Interferons (IFNs) are considered as one of the first lines of host innate immune defense against viral infections (46). Type I IFN family includes a group of structurally similar cytokines and consists of IFN-α (13 different subtypes), IFN-β, IFN-ɛ, IFN-κ, IFN-ω, IFN-δ, IFN-ζ, and IFN-τ (71). Among these, IFN-α, IFN-β, IFN-ɛ, IFN-κ, and IFN-ω exist in humans, whereas IFN-δ and IFN-τ have been described only for pigs and cattle, respectively, and do not have human homologs (68). Type I IFNs can induce a direct antiviral response within infected and the neighboring cells via the upregulation of molecules that can antagonize virus replication (57). Other than antiviral messengers, IFNs possess a wide range of other biological activities, that is, inhibition of cell proliferation, regulation of apoptosis, and immunomodulation (34,45).
Based on their antiviral activity, type I IFNs are an important therapeutic option for the treatment of viral infectious diseases. For example, IFN-α and IFN-β are part of the standard treatment for chronic hepatitis B and C virus infections (29,47,95). Similarly, IFN-τ is effective in inhibiting HIV infection (14). In addition to IFNα/β, the distinct antiviral activities and potential clinical implications of other type I IFNs (i.e., IFN-ɛ, IFN-ω, IFN-κ, IFN-δ, IFN-τ, and IFN-ζ) have also been reported earlier. However, individual antiviral and immunomodulatory properties of IFN-ɛ, IFN-ω, IFN-κ, IFN-δ, IFN-τ, and IFN-ζ have not been investigated as widely as IFN-α and IFN-β (51).
In this review, we summarize the pivotal role of type I IFNs in MERS-CoV, SARS-CoV, and SARS-CoV-2 infections. Importantly, based on the previous studies and our current understanding of the knowledge, we propose the potential therapeutic implications of synergistic use of type I and type II IFNs against COVID-19.
Type I IFNs Induction and Signaling
Microbial pathogen-associated molecular patterns are sensed by the host pattern recognition receptors resulting in the induction of type I IFNs. Generally, there are two pathways for the induction of type I IFNs in response to RNA viruses, that is, intracellular pathway and the endosomal pathway (94). The intracellular pathway of type I IFNs induction involves the by-products of virus replication such as double-stranded RNA (dsRNA) or 5′ triphosphorylated single-stranded RNA (ssRNA) leading to the activation of the transcription factor IRF-3 (IFN regulatory factor 3). Both TBK-1 and IKKɛ that are stimulated by the RIG-I (RNA sensing molecule) and MDA5 (dsRNA sensing molecule), phosphorylate IRF-3. Optimum activation of the IFN-β promoter requires the cooperative action of NF-κB and AP-1 transcription factors (35). Viral dsRNA is also sensed by the PKR, which in turn activates NF-κB (25), whereas AP-1 is activated by the stress-responsive kinase Jun (26). The endosomal pathway of type I IFNs involves viral ssRNA and dsRNA sensing via TLR7/8 and TLR3, respectively, followed by the activation of IRF-7, IRF-3, and the transcription of IFN-α/β (35,94,97,104).
Type I IFNs (IFN-α and IFN-β) bind to the same heterodimeric receptor (IFNAR1/IFNAR2) followed by the activation of the protein tyrosine kinases, that is, JAK1 (Janus kinase 1) and TYK2 (tyrosine kinase 2). These protein tyrosine kinases (JAK1/TYK2) then phosphorylate the STAT1 (signal transducer and activator of transcription 1) and STAT2 resulting in the dimerization and subsequently the intranuclear translocation of the phosphorylated STAT1 and STAT2 (40,49,87). Inside the nucleus, STAT1/STAT2 dimer assembles with IRF9 (IFN-regulatory factor 9) and gives rise to a trimolecular complex called ISGF3 (IFN-stimulated gene factor 3), which on binding to its cognate IFN-stimulated response elements stimulates the transcription of several hundred IFN-stimulated genes (ISGs). These ISGs encode proteins that inhibit the viruses by blocking the viral transcription, translation, and replication, and degrading the viral nucleic acids (40).
Protective Effect of Type I IFNs Against MERS-CoV, SARS-CoV, and SARS-CoV-2
For the control of coronaviruses, the IFN response is considered very pivotal. Several animal coronaviruses have shown sensitivity to type I IFNs (94). Both SARS-CoV and mouse hepatitis virus (MHV) produce an adequate amount of viral dsRNA; an important inducer of type I IFNs (94,98,100). Previously, several studies have suggested that exogenous administration of IFN-α/β can inhibit the growth of SARS-CoV (15,33,86,91,107). Many in vitro and in vivo studies have demonstrated the use of IFN-I treatment against MERS-CoV and SARS-CoV (90). For example, in response to IFN-β1b therapy, an amelioration in the lung pathology and reduction in viral load have been reported in marmosets infected with MERS-CoV (8). Similarly, a combination of ribavirin with IFN-β significantly inhibited SARS-CoV replication in animal and human cell lines (62). Furthermore, combination therapy with IFN-α2b and ribavirin also resulted in better clinical outcome in macaques infected with MERS-CoV (19). Administration of IFN-α before SARS-CoV challenge reduced the virus location and pulmonary damage in macaques (30) demonstrating protective results in response to IFN-I therapy in animal studies of SARS-CoV infection.
As discussed earlier, many in vitro and in vivo studies have shown a relative success against MERS-CoV and SARS-CoV; however, the results of IFN-I therapy in human studies are still inconclusive and unsatisfactory (90). For example, IFN-I therapy along with steroids proved efficient in ameliorating the lung abnormalities in human patients infected with SARS-CoV (54). Similarly, an improved survival proportion was also observed in MERS-CoV patients at 14 days, but not at 28 days after diagnosis followed by IFN-α2a/ribavirin administration (65).
No therapeutic effect of IFN-α/ribavirin therapy has been reported in a study conducted by Al-Tawfiq et al. (1). This difference in survival rate in response to IFN-α/ribavirin therapy might be due to state of illness and time of diagnosis as all the patients in the later study (1) were critically ill at the time of initiation of IFN therapy. Similarly, a combination of IFNα2a/ribavirin therapy failed to decrease mortality after MERS infection (2). In a recent study, the administration of IFN-β in combination with lopinavir-ritonavir improved the pulmonary functions but failed to reduce the viral load and lung pathology (82). The reason for inconsistency in results in human studies could be attributed to many factors such as IFN-I subtypes and timing of IFN administration. Although beneficial effect of IFN-α against coronaviruses has also been documented previously (13,54,93), IFN-β was shown to inhibit the coronaviruses replication more effectively in comparison with IFN-α (78,90).
Further evidence regarding the superior antiviral role of IFN-β among IFN-I subtypes has also been reported against MERS-CoV (9,17,32) and SARS-CoV (33). The timing of IFN therapy could be another important factor in driving the clinical outcome of the infection as IFN therapy before infection resulted in a better antiviral response in animal models of SARS-CoV (30). In addition to this, in vivo studies in mice demonstrated that IFN-I therapy shortly after virus infection gave positive effects as compared with late administration that failed to limit to virus replication (12).
The important role of type I IFNs in response to coronaviruses is also exhibited by viral evasion of the IFN pathway and decreased ISG production during the disease. Previous studies have shown that type I IFNs were not induced in fibroblast cells (nonspecialized cells lacking TLRs) that were infected with either SARS-CoV or MHV. However, treatment of these cells with polyinosinic:polycytidylic acid resulted in IFN induction (98,109). Thus, it looks that coronavirus macromolecules appear to be invisible to immune sensors (i.e., RIG-I, MDA5, and TLR3) in these cells partly because dsRNA, a potent stimulator of the innate immune system, is buried in double-membrane vesicles (DMVs) (67,98). DMVs could form a protective microenvironment that prevents virus from being detected by host cell sensors (28,85). Moreover, Nsp1 also inhibits type I IFN signaling in both SARS-CoV- and MHV-infected cells, in part by inhibiting STAT1 phosphorylation thereby impairing a proper innate immune response (99,111).
Besides Nsp1, the inhibition of type I IFNs induction by other viral proteins of coronaviruses has been reviewed by Perlman and Netland (67). A few studies have suggested a downregulation of IFN and ISGs expression in peripheral blood mononuclear cells derived from the SARS patients (72,105). Similarly, downregulation of RIG-I, MDA5, and IRF-3/7 was also reported in an individual infected with MERS-CoV (fatal outcome) as compared with the recovering patient (20). Despite the inhibition of type I IFN induction by coronaviruses, IFN-α/β have been detected in infected mice and humans (6,73). A significant IFN production by MHV- and SARS-CoV-infected plasmacytoid dendritic cells has also been reported by (7). Therefore, the important role of type I IFNs against coronaviruses cannot be ignored in case of coronaviruses infections.
Several knockout mice models have highlighted the importance of type I IFNs in controlling the coronaviruses infections. It was observed that mice deficient in STAT1 were more susceptible to SARS-CoV-induced pathology of various organs (23,36). Mice lacking type I IFN receptors (IFNAR1 or IFNLR1) showed an enhanced replication of SARS-CoV and MHV (7,61). Moreover, mice lacking both type I and type III IFN receptors (double knockout) or STAT1 showed a significantly higher titer of a mouse-adapted strain of SARS-CoV (rMA15) as compared with the wild-type mice (55). Surprisingly, the disease outcome of both IFNAR1−/−, IFNLR1−/− mice (23), and double-knockout (IFNAR1−/−/IFNLR1−/−) mice (55) was quite similar as compared with wild-type mice infected with SARS-CoV. However, decreased survival and clinical signs were observed in STAT1-deficient mice (STAT1−/−) (23,55). These findings suggest the possible involvement of the IFN-receptors-independent signaling mechanism via STAT1 to SARS-CoV-associated pathology. An increased pathology in the lungs was associated with SARS-CoV infection in mice lacking MyD88, TLR3, or TRIF (81,96).
A recent report (not peer-reviewed yet) have suggested a substantial in vitro anti-SARS-CoV-2 activity of IFN-α as its treatment resulted in a decrease in virus titer several folds (53). In vitro studies have revealed that SARS-CoV-2 is more sensitive to type I IFN treatment as compared with the SARS-CoV (56). This difference in sensitivity to IFN could be due to the Orf6 and Orf3b proteins. Both of these proteins of SARS-CoV act as IFN-antagonist by inhibiting the intranuclear translocation of STAT1 and disruption of IRF3, respectively (22,42). However, in the case of SARS-CoV-2, both of these proteins are truncated suggesting a possible loss in anti-IFN activity of SARS-CoV-2 (53). The beneficial effect of type I IFN therapy was also reported in a study conducted by Hung et al. (39). In this study, early treatment of patients (with mild to moderate COVID-19) with triple antiviral drugs (IFNβ-1b, lopinavir-ritonavir, and ribavirin) was found to be effective in alleviating symptoms and shortening the duration of viral shedding in comparison with lopinavir-ritonavir alone (39). In a study, treatment with IFN-α2b with or without arbidol substantially reduced the duration of detectable virus in the upper respiratory tract of COVID-19 patient (110). The therapeutic effectiveness of IFN-α2b against COVID-19 has also been reported; however, the results are not peer-reviewed yet (66).
Immunopathogenic Role of Type I IFNs in MERS-CoV, SARS-CoV, and SARS-CoV-2 Infection
Type I IFNs have been implicated in several noninfectious pathological conditions. Innate immune responses, particularly proinflammatory cytokines, have long been considered as possible crime partners in the outcome of disease severity (64). The involvement of type I IFNs in disease severity and pathology is controversial and might be misleading. An increase in IFN-α level was observed in MERS patients with severe disease but not in patients having a mild disease. However, the disease outcome could not be correlated with the serum level of IFN-α as this increase in serum IFNα level during the clinical course of the disease may be due to the exogenously injected pegylated IFN-α2a in this study (41).
Some animal models have also suggested the role of IFN-α in disease outcome. For example, infection of MA15 (a mouse-adapted SARS coronavirus) into BALB/c mice resulted in pronounced signs of disease and substantial weight loss. About 85% of the mice died 8 days postinfection. A delayed expression of proinflammatory cytokines along with IFN-β was observed in these severely ill mice. In contrast, mild to moderate disease severity and weight loss were observed in BALB/c mice lacking type I IFN receptor (IFNAR−/−) (11). From these findings, we can conclude that type I IFNs are at least partly involved in SARS- and MERS-induced pathology. Clinical findings of patients infected with SARS-CoV-2 have indicated a correlation between inflammatory cytokines and high mortality (108). Also, recent reports in Lancet showed ARDS (acute respiratory distress syndrome) as one of the main causes of deaths in patients infected with SARS-CoV-2 (38,103). A higher mortality rate was also observed in a recent study conducted by Wu et al., in which 41.8% of patients (out of 201 patients) developed ARDS, and of those, 52.4% died (102). ARDS is an immunopathological event for MERS, SARS, SARS-CoV-2 infections (103) and developed due to excessive release of proinflammatory cytokines, including IFN-α, and chemokines, such as CCL2, CCL3, CCL5, CXCL8, and CXCL9 (52).
Future Potential of Synergistic Use of Type I and Type II IFNs Against SARS-CoV-2
Synergistic effect of type I IFNs and IFN-γ on virus replication
Type I IFNs (IFN-α and IFN-β) and type II IFN (IFN-γ) are important components of the host immune response to viral infections. IFN-α and IFN-β are produced by most of the cells as a direct response to viral infection, whereas IFN-γ is synthesized almost exclusively by activated natural killer (NK) cells and activated T cells in response to virus-infected cells (70). The observation of a synergistic effect between type I and type II IFNs is not novel, as similar observations have been reported in other viral infections. Several studies have shown that IFN-γ synergizes with IFN-α/β to inhibit the replication of both RNA and DNA viruses (16,21,27,50,79,80,83). Both in vitro and in vivo trials have proved the synergistic effect of IFN-γ with IFN-α/β against herpes simplex virus 1 (HSV-1) (74).
The synergistic effect of IFN-γ with IFN-α has also been demonstrated against HSV-2 replication in Vero cells. Coadministration of IFN-α with IFN-γ resulted in reduction in HSV-2 plaque formation, replication, and the onset of viral cytopathic effects (69). Coadministration of IFN-α and IFN-γ increases the antiviral effect in a model of MHV type 2 infection (24). A potent antiviral synergy was also reported against the hepatitis C virus (HCV) followed by simultaneous administration of type I/II IFNs combinations in HCV replicon system (44). A potentially beneficial role of type I/II IFN combination therapy has also been documented against Lassa virus (3). Further evidence regarding the synergistic role of IFN-γ and innate IFNs (IFN-α/β) is provided by in vitro studies against human cytomegalovirus (HCMV). Both IFN-α/β and IFN-γ act synergistically to inhibit the replication of HCMV (75).
More importantly, the synergistic antiviral action of both type I and type II IFN has also been reported against SARS-CoV. Synergistic anti-SARS-CoV activity was observed when 100 U/mL of IFN-γ was coadministered with various concentrations of IFN-β in Vero cells. However, simultaneous administration of IFN-γ (1 U/mL) along with various concentrations of IFN-β failed to produce a synergistic effect, which suggests that a certain level of IFN-γ is required for optimum synergistic activity of both IFN-β and IFN-γ (78). Sainz et al. determined the synergistic effect of IFN-β and IFN-γ against two strains of SARS-CoV, that is, Urbani and HK. A significant inhibition in both strains of SARS-CoV was observed in cell cultures stimulated with 100 U/mL of both IFN-β and IFN-γ compared with the cultures treated with 100 U/mL of IFN-β and IFN-γ, individually (76). A combination of type I and type II IFN therapy resulted in a significant increase in antiviral activity against SARS-CoV infection in Vero cells when compared with individual IFN treatments, which yielded a minor inhibitory effect on SARS-CoV replication (77).
The exact mechanism by which type I and II IFN synergize to inhibit SARS-CoV is yet unclear. A previous study has demonstrated the synergistic interaction between IFN-α and IFN-γ through the induction of one subunit of the transcription factor ISGF3 (48). Activation of type I IFN receptors by type II IFN could be another possible explanation for the underlying synergistic mechanism of both type I and type II IFNs. IFN-γ induced a time- and dose-dependent expression of mRNA for IFNAR-1 and IFNAR-2 in a human hepatoma cell line, HepG2 cells. Moreover, combined treatment of both type I and type II IFN also resulted in significantly higher 2-5A-synthetase activity than that by type I IFNs alone (58). Scagnolari et al. also described a marked increase in transcriptional levels of IFN-induced proteins (i.e., 2′-5′-oligoadenylate synthetase and p56) in Vero cells stimulated with a combination of IFN-α or -β and IFN-γ in comparison with the cells treated with IFN alone, suggesting the possible synergistic mechanism (77). Figure 1 describes the proposed working mechanism of the synergistic effect of type I and II IFN against SARS-CoV-2.
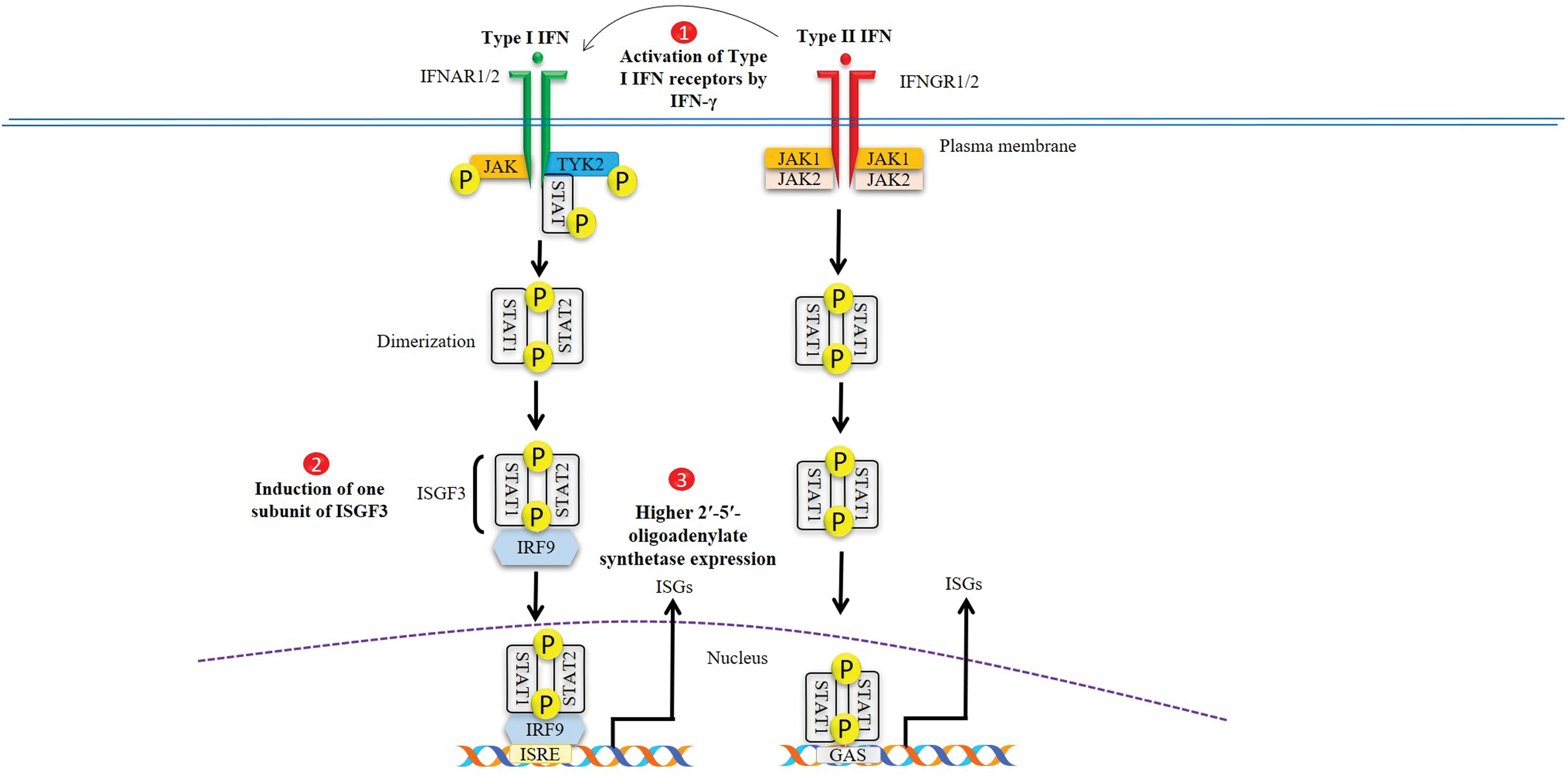
Proposed mechanisms for synergistic antiviral effect of type I/II IFNs on SARS-CoV-2 replication. SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; IFN, interferon. Color images are available online.
Suppressive effect of type I IFNs and IFN-γ on ILC2-induced immunopathology
An optimal antiviral response is accompanied by activation of beneficial immune responses, whereas limiting the potentially damaging responses at the same time. Type I IFNs stimulation has been shown to enhance MHC II expression and presentation of antigens (43,60,84). In addition to this, type I IFNs have been found to restrict the damaging immune responses that can lead to tissue pathology. Upon infection of the airways, ILC2 cells contribute to type 2 immunopathology and loss of airway function (10). Mouse models of allergic inflammation induced by viral lung infections have revealed that ILC2 cells play a critical role in driving allergic lung responses and, therefore, need to be tightly regulated for the control of pulmonary inflammatory pathologies (37). In a model of influenza infection, the absence of the type I IFN receptor resulted in significant virus-induced immunopathology. Duerr et al. demonstrated that this pathology was mediated by an upregulation of type 2 cytokines from unregulated innate lymphoid type 2 cells (ILC2s) (18). Thus, type I IFN suppresses ILC2 function during virus infection.
IFN-γ is a type II IFN and an important component of the antiviral response. Major functions of IFN-γ include an increase in antigen presentation, maturation of dendritic cells, and expression of MHC I and MHC II. IFN-γ is a critical propellant of the Th1 response (46). IFN-γ has similar features with type I IFNs in many ways. For example, alike type I IFN, IFN-γ can also suppress ILC2 proliferation and type 2 cytokine production (18,63). Figure 2 represents a proposed mechanism regarding the possible role of type I and II IFNs in MHC expression and suppression of ILC2-induced immunopathology followed by virus infection. During IAV infection in mice, Stifter et al. demonstrated that a functional interplay is required between both type I and type II IFN signaling to prevent excessive immunopathology (89). Moreover, experiments in STAT1-deficient (a key transcription factor downstream of type I/type II IFN signaling) mice resulted in an increase in lung pathology due to increased cytokine production from ILC2 and ILC3s during RSV infection (88).
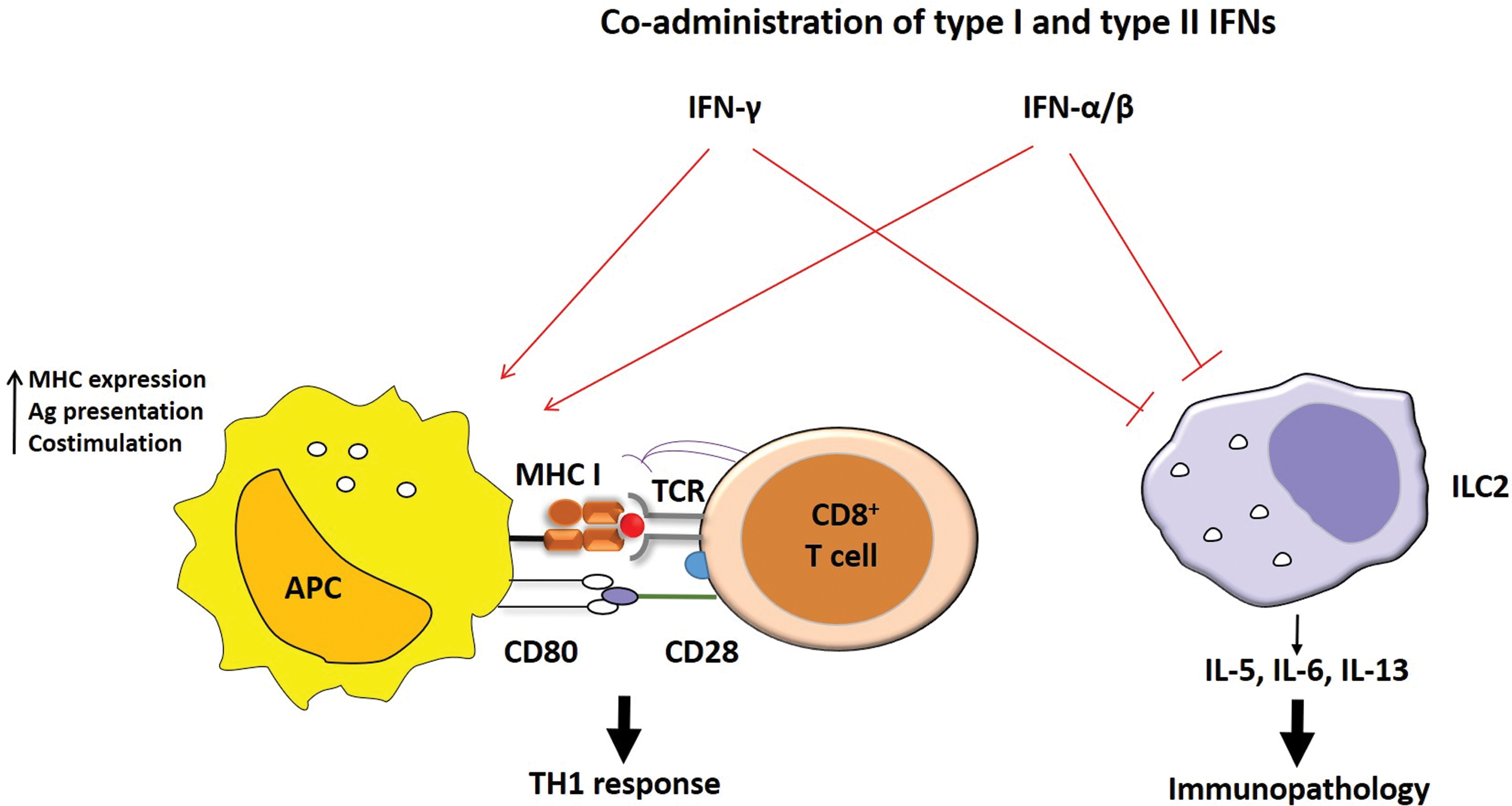
Probable model describing the working mechanism of exogenous coadministration of type I and II IFNs. Type I and II IFNs likely to work in concert with each other to promote APC maturation, upregulation of costimulatory molecules, and antigen processing and presentation to promote Th1 polarization, whereas simultaneously suppressing ILC2-mediated immunopathology. ILC2, group 2 innate lymphoid cells. Color images are available online.
In vivo experiments have revealed the suppression of IL-33-induced ILC2 proliferation followed by IFN-γ administration (18). Similarly, Han et al. demonstrated the inhibition of ILC2 by IFN-γ thus preventing the development of an asthma phenotype in rhinovirus-infected mice (31). However, in a mouse model of influenza infection, IFN-γ production resulted in an increased susceptibility to influenza virus infection. Whereas deficiency of IFN-γ led to increased IL-5 and amphiregulin release from ILC2s resulting in enhanced tissue repair and improved survival. However, this increase in survival was most probably associated with improved tissue integrity due to amphiregulin release from ILC2, not the IL-5 (5). The role of amphiregulin in the maintenance of tissue support and lung repair has been reported previously (59,106). Besides amphiregulin, pulmonary eosinophils can also contribute to tissue repair and enhanced survival (4,92). Other factors such as differences in mouse strains, viral strain, gene deficiency, and/or immunization routes may also result in discrepancies in the aforementioned studies regarding the suppressive role of IFN-γ on ILC2-induced immunopathology.
This contrast in results of cytokines was also observed in some human patients. In the sera of SARS patients, elevated levels of IFN-γ, IL-1, IL-6, and chemokines, that is, CCL2, CXCL10, and IL-8 were detected (101). Importantly, in another study, a reduction in inflammatory biomarker, that is, IL-6 was reported after inhaled IFN-α2b therapy (110). However, the number of individuals in this study was small and all were suffering from moderate COVID-19. Also, the clinical findings of Wong et al. (101) and Zhou et al. (110) are mainly derived from the blood of patients and do not allow the assessment of immune responses within the lung. Many other factors such as time of disease onset, time of admittance to the hospital after disease onset, level of disease severity, different treatment regimens provided to the patients in these studies, and already prevailing comorbidities may have a varying degree of influence on the clinical outcome of cytokine profile. So, before generalization of these results, further studies should be investigated to address these factors.
Concluding Remarks
Knowledge gained from studies on SARS-CoV and MERS-CoV will be valuable for determining the suitability of IFN-I as a treatment strategy in COVID-19. Previous findings of several studies support the antiviral potency of type I IFNs particularly IFN-β against MERS-CoV and SARS-CoV (8,17,32,33,78,82,90). Based on our previous knowledge regarding type I IFNs therapy against MERS-CoV and SARS-CoV, it is proposed that IFN-β could be a potential treatment option against SARS-CoV-2 infection in early phase of the disease, especially when used in combination with antiviral drugs such as lopinavir/ritonavir, remdesivir, or ribavirin.
In summary, our assessment of previous results demonstrates that a combination of type I/II IFNs may be more powerful than the use of either agent alone against SARS-CoV-2 replication. Having established that simultaneous administration of both type I IFNs and IFN-γ were significantly effective in preventing many viruses, including SARS-CoV replication, we speculate that synergistic use of type I/II IFNs may have therapeutic implications against COVID-19. Furthermore, route, timing, duration, and dosage of IFN combination therapy could be the critical parameters underlying the success or failure of this combination therapy against COVID-19. Therefore, immediate attention should be given to determine the role of type I IFNs and IFN-γ as a therapeutic strategy in the treatment of SARS-CoV-2 infections.
As for the role of type I IFNs and IFN-γ in immunopathology is concerned, several studies have shown the suppression of ILC2-mediated immunopathology in response to type I and type II IFN administration. However, we propose that biological role of type I/II IFNs combination therapy in the suppression of ILC2-mediated immunopathology is still poorly understood and, therefore, further assessment of innate immune responses should be done at the local site (within the lungs) instead of patients' blood to validate any decisive conclusion. Notably, understanding how ILC2s are regulated in the context of a viral infection particularly SARS-CoV-2 will be necessary and may provide us the rationale for combined and improved therapeutic approaches to treat COVID-19.
Footnotes
Acknowledgment
The author thanks the Higher Education Commission, Pakistan.
Author Disclosure Statement
All the authors declared no conflict of interest.
Funding Information
No funding was received.