
Abstract
The initial immune response to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) includes an interferon-dependent antiviral response. A late and uncontrolled inflammatory response characterized by high activity of proinflammatory cytokines and the recruitment of neutrophils and macrophages develops in predisposed individuals and is potentially harmful in some cases. Interleukin (IL)-17 is one of the many cytokines released during coronavirus disease 2019 (COVID-19). IL-17 is crucial in recruiting and activating neutrophils, cells that can migrate to the lung, and are heavily involved in the pathogenesis of COVID-19. During the infection T helper 17 (Th17) cells and IL-17-related pathways are associated with a worse outcome of the disease. All these have practical consequences considering that some drugs with therapeutic targets related to the Th17 response may have a beneficial effect on patients with SARS-CoV-2 infection. Herein, we present the arguments underlying our assumption that blocking the IL-23/IL-17 axis using targeted biological therapies as well as drugs that act indirectly on this pathway such as convalescent plasma therapy and colchicine may be good therapeutic options.
Introduction
The coronavirus disease 2019 (COVID-19), the result of an infection with the new severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has rapidly spread worldwide. In December 2019, the new virus emerged in Wuhan (China) and because of its rapid worldwide spread, the World Health Organization (WHO) declared the COVID-19 outbreak a pandemic in March 2020. SARS-CoV-2 shares genetic information and similar clinical manifestations with SARS-CoV-1 and the Middle East respiratory syndrome coronavirus (MERS-CoV) (71). They belong to the β-coronavirus cluster and the nucleotide sequence homology between SARS-CoV-2 and SARS-CoV-1 is 79.5% (41). There is also high homology with bat coronaviruses. Therefore, based on the current evidence, the epidemic started with the animal (bats) to human transmission although the intermediate animal remains to be determined (71).
Knowledge about the immune response to SARS-CoV-2 is very important for not only the design of an effective vaccine against it but also a suitable treatment. There are a few studies of the immune response to SARS-CoV-2, which show a weak initial response from interferon (IFN) that allows viral replication and a late immune response that is characterized by high activity of proinflammatory cytokines (including interleukin [IL]-1, IL-6, tumor necrosis factor [TNF], IP-10, and MCP-1) (36) and the recruitment of neutrophils and macrophages, which generates an aggressive immune response that could be harmful to the host in certain cases (72). This inappropriate response to the virus is the result of a complex interaction between factors related to the virus (e.g., viral load), the host (e.g., genetic polymorphisms), and the environment (66). The molecular mechanisms regulating emerging coronavirus pathogenesis are complex and include virus–host interactions associated with entry, replication, egress, and innate immune control.
It is remarkable that children who are more susceptible than adults to viral infections such as influenza do not show this inappropriate response and have a benign course with a good prognosis during a SARS-CoV-2 infection in most cases (28). Nevertheless, new concerns about a novel severe Kawasaki-like disease (i.e., a rare acute pediatric vasculitis, with coronary artery aneurysms as its main complication) in children that is related to COVID-19 could change the understanding of this disease in this age group (89). Moreover, immunocompromised patients are not at increased risk of severe complications compared with the general population (21).
The T helper 17 (Th17) cells and IL-17-related pathways are not effectively controlling intracellular microorganisms such as a viral infection and induce recruitment of neutrophils and thus cause more tissue damage (34). However, there are evidence that supports a neutrophil role in the progression of the SARS-CoV-2 infection (40,96). Therefore, we present arguments that could explain the role of Th17 in COVID-19 and the possible drugs that could have a therapeutic role in the treatment of this infection by inhibiting the IL-23/IL-17 axis.
Innate and Adaptive Immune Response Against SARS-CoV-2
Innate immune response
Once the SARS-CoV-2 interacts with the angiotensin-converting enzyme 2 (ACE2) receptor through its Spike S1 protein to mediate infection of the target cell, it is internalized in an endosomal vesicle and then stripped, and its ssRNA passes to the cytosol where it begins its replication and expression and then assembles new viral particles (23). At this point, both pattern recognition receptors and endosomal Toll-like receptors (TLR) 3 and 7 and cytosolic receptors (retinoic-acid inducible gene I, melanoma differentiation-associated gene 5) recognize the ssRNA. As a result, they activate the downstream signaling cascade, that is, NF-κB, interferon regulatory factor (IRF) 3, and IRF7, and their nuclear translocation. The entry of these transcription factors into the nucleus induces the transcription of genes for alfa and beta IFN and proinflammatory cytokines (23). In fact, Zhang et al. (99), through a microengineered human alveolus chip infected by SARS-CoV-2, developed transcriptional analyses and showed a broad innate immune response and antiviral responses, including defense response to the virus, IFN-I signaling pathway, and cytokine-mediated signaling pathway.
IFNs are important for controlling viral replication, activating natural killer (NK) cells, and increasing the presentation of viral peptides in HLA class I to CD8 T lymphocytes. Yan et al. (70,87,95) analyzed subsets cells using flow cytometry in convalescent patients with COVID-19. They found that NK cell counts were significantly higher in patients with COVID-19 than in control patients. Emphasizing the high expression of NK cells is important in innate immune response against SARS-CoV-2.
Increase in monocyte number (70,87) (i.e., CD14++CD16+), HLA-DR expression (70), and activation of innate immune responses with a clear increase in type I IFN and proinflammatory cytokines (84,87) in a time-dependent immune response (84) or depending on the severity of the COVID-19 cases (70,84) has been demonstrated. Some of these findings have been confirmed in postmortem lung biopsies (74,78) or through a COVID-engine inferential statistics platform (78).
Of interest, a pronounced IFN response but attenuated activation of other innate immune pathways, including TLR, IL, and chemokine signaling has been shown in an upper airway host transcriptional response analysis developed in 93 patients with COVID-19 by Mick et al. (55). In addition, they showed that some elements of the IFN response to SARS-CoV-2 were attenuated compared with other viruses' genes, such as IRF7 and OASL. Strikingly, the IL-1 and NLRP3 inflammasome pathways were markedly less responsive to SARS-CoV-2, commensurate with a signature of diminished neutrophil and macrophage recruitment.
Neutrophils play an important role as the first line of innate immune defense as viral infection by interaction with other immune cell populations, virus internalization and killing mechanism, cytokines release, degranulation, oxidative burst, and neutrophil extracellular traps (NETs) (58). SARS-CoV-2 was observed in high amount in blood of granulocyte myeloid-derived suppressor cells, defined as HLA-DR+CD14−CD15+CD11b+CD33+ and immature forms of CD10− neutrophils (CD15+CD66b+CD10−). Both cells were more pronounced in the severe COVID-19 cases than in healthy donors (67). SARS-CoV-2 infection induces neutrophil accumulation, this induces reactive oxygen species release, such as superoxide radicals and H2O2, leading to oxidative stress that contributes to the cytokine storm and blood clots formation and this contributes to acute respiratory distress syndrome (ARDS) (13,42). Decreased expression of the antioxidant enzyme superoxide dismutase 3 was observed in lung of patients with SARS-CoV-2 infection (1). The activation of neutrophils also induces NET production. Elevated level of NETs was observed in COVID-19 patients and this level was associated with disease severity (86). The formation of NETs is associated with blood clots in COVID-19 (56).
The diverse findings in the type of innate immune response may be related to the interference of the SARS-CoV-2 in the Janus kinase (JAK)-STAT pathway (10,17) (key component in the IFN response pathway), the mevalonate pathway (33), and mitochondria antiviral signaling (80), which in turn impact the activity patterns of the inflammasome and are under study.
Adaptive immune response
Dendritic cells (DCs) located in the lung interstitium recognize and phagocyte viral particles and/or lysed cell debris from the viral infection and target regional lymph nodes to present viral peptides to both CD4 and CD8 T lymphocytes playing a role of link between innate and adaptive immunity. Viral components also travel to the lymph node along with cytokines such as IFN and IL-12 that are produced by macrophages and DCs. In the lymph nodes, CD4 T lymphocytes are activated by DCs in the presence of IL-12, which differentiates into Th1 effector cells. These produce IL-2 and IFN gamma that acts on CD8 T lymphocytes and B lymphocytes to assist in their differentiation into cytotoxic cells and immunoglobulin M (IgM)-producing plasmocytes (37,77,88). These CD8 T lymphocytes collaborate to decrease viral replication and lysate the virus-infected cells in the lung tissue. Antibodies, first IgM and then immunoglobulin G, assist in the removal of extracellular viral components and the destruction of infected cells by recognizing viral antigens on the cell surface and inducing NK cell activity (23,72). In this context, DCs and endothelial cells incite a Th17 response that begins to control the response pattern. However, this Th17 response, far from controlling the infection, is prolonged and increasingly reinforced, thus leading to the catastrophic picture of the cytokine storm described in patients with severe disease (83).
The analysis of neutralizing antibody against SARS-CoV-2 shows the complexity of the immune response to COVID-19 (38). Some studies highlight that not only a neutralizing antibody but also cellular immunity (39) has a role in the elimination of SARS-CoV-2. Other studies have shown the importance of neutralizing humoral immunity on disease progression (32). The evaluation of the role of the humoral response in the pathogenesis of SARS-CoV-2 is in continuous development (44).
Empirical Data: Th17 Response to SARS-CoV-2
Transforming growth factor-β (TGF-β), IL-1β, IL-6, and IL-18 induces a Th17 response and increase neutrophil production and recruitment. In SARS-CoV-2, the neutrophils induce a Th17 response via nitric oxide synthases (67) These effector lymphocytes reach the lung tissue and produce IL-17, IL-21, and IL-22, which increase production by the local cells of antimicrobial peptides, IL-8, and collagen. IL-8 recruits more neutrophils and these produce more tissue damage. This Th17 response does not effectively control intracellular microorganisms such as a viral infection and induce recruitment of neutrophils and thus causes more tissue damage (34).
Qin et al. observed high levels of neutrophils in 452 patients with COVID-19 along with severe infections compared with patients with nonsevere infections (p < 0.001) (73). Recently, immunological response to the Th17 phenotype was seen in patients with moderate and severe SARS-CoV-2 (7). Some cytokines that are involved in the Th17 immune response such as IL-2, IL-6, IL-10, granulocyte-macrophage colony-stimulating factor (GM-CSF), IP-10, MCP1, MIP1A, and TNF-α have been reported to be elevated in some studies although mainly in severe cases (18,36). In fact, Xiong et al. (93) described the transcriptional signatures of host inflammatory responses to SARS-CoV-2 by transcriptome sequencing of the RNAs isolated from bronchoalveolar lavage fluid (BALF) and peripheral blood mononuclear cell (PBMC) specimens of COVID-19 patients. Distinct host inflammatory cytokine profiles of SARS-CoV-2 in patients were described. For PBMC samples, certain cytokine-related pathways enriched downregulated genes such as “IL-17 signaling pathway” and “TLR signaling pathway,” and so on, and for BALF samples, an excessive cytokine release such as CXCL10/IP-10 (a product of the IL-23/IL-17 axis inflammation) was found. Finally, Sadeghi et al. (79) evaluated the frequency of Th17 cells in severe SARS-CoV2 patients by flow cytometry to examine the inflammatory responses of these cells. Based on the results, they found a significant increase in the number of Th17 cells, the expression levels of related factors (RORγt, IL-17, and IL-23), and the secretion levels of IL-17 and IL-23 cytokines in 40 COVID-19 patients compared with controls (79).
Another relevant agent in the immunological response to SARS-CoV-2 is inflammasome. Recently, the role of the SARS-CoV-1 unique domain, which is also found in SARS-CoV-2 (75% identity), has been highlighted given that modulated NLRP3 inflammasome-dependent CXCL10-mediated pulmonary inflammation with significantly upregulated expression of CXCL10 in human lung epithelial cells has resulted in severe inflammatory responses and diffuse alveolar damage symptoms (16). This is important given the crucial role that the nuclear receptor, RORγt, plays in the development of Th17 cells of the adaptive immune system and the NLRP3 inflammasome (8).
Th17 Response to Other Viral Diseases
These high levels of proinflammatory cytokines have been seen in SARS-CoV-1 and MERS-CoV (50,91). This highlights the fact that patients with severe SARS-CoV-1 express high serum levels of TGF-β, a cytokine that induces the development of the Th17 lineage (47). In addition, there is evidence of a mechanism whereby the SARS-CoV-1 nucleocapsid protein modulates TGF-β signaling to block apoptosis of SARS-CoV-1-infected host cells and, at the same time, facilitate tissue fibrosis (101). This is important given the striking relationship between TGF-β and the contribution of Th17 cells to fibrosis in different hepatic and lung murine models (69).
SARS-CoV-1 has been associated with inflammasome activation (15) by calcium ion transportation that leads to IL-1β overproduction (63), and by direct caspase-1 activation or through an enhanced potassium efflux (98). Ahn et al. (3) identified significantly dampened activation of the NLRP3 inflammasome in bat primary immune cells compared with human or mouse counterparts. The inflammasome activation induces IL-1β secretion and the activation of a Th17 response (25).
Furthermore, in a human in vitro model for investigating genome-wide host responses to SARS-CoV infection, cytokine production together with lung trafficking chemokines such as IL-8 and IL-17 (lung directed and highly expressed), possibly activated through the TLR9 signaling pathway, was demonstrated. Representation of selected immune-related genes upregulated during the first 12-h postinfection, including Th17 pathway genes, was also demonstrated (62).
In addition, elevated levels of IL-17, Th17 mediators, and IL-17-responsive cytokines were found in serum samples from patients infected with swine-origin H1N1 influenza virus isolated in China during the 2009 pandemic. IL-17 deficiency or treatment with monoclonal antibodies against IL-17 ameliorated acute lung injury induced by the H1N1 virus in the mice model. These results suggest that IL-17 plays an important role in H1N1-influenza-induced acute lung injury (45).
It has been reported that Th17 and Treg cells could determine the nature of the immune response in the respiratory syncytial virus (RSV) infection and, therefore, disease severity. In RSV-infected mice, Th17 cells were found to be 2.3 times higher in number than in the uninfected group, whereas the number of Tregs was 0.7 times lower than that of the control group. This showed that the imbalance of Treg/Th17 cells will lead to immune dysfunction and the progression of infectious diseases as well as an inflammatory response. Leptin, tachykinin, substance P, and some domains of RSV proteins are described as elements that facilitate Th17 cell differentiation (51,90). Furthermore, in some animal models in RSV, digoxin has been identified as the one that selectively inhibits IL-17 production by antagonizing its transcription factor (RORγt), which is the most specific transcription factor for Th17 cells. Thus, it demonstrates that digoxin reduced the signs of clinical illness after an RSV infection and reduced lung pathology (53). Th17 cells are involved in the occurrence and progression of ARDS of different etiologies. Some transcriptional factors (i.e., histone acetyltransferase p300) in cells from patients with ARDS interact with and stabilize the RORγt, a key factor in the exaggerated Th17 response. This shows that the p300 inhibitor caused a decrease in the mRNA and protein levels of RORγt. As a result, IL-17 declines, and this reduces lung tissue inflammation and lung injury score (53).
A similar case has been depicted in patients with chronic hepatitis B and in those with chronic hepatitis C. The number of both Treg and Th17 cells was elevated although the increase in Th17 cell frequency was more pronounced, and thus, the ratio of Th17/Treg was higher in hepatitis B virus-infected patients than in uninfected individuals (90). In addition, in hepatitis C virus (HCV)/human immunodeficiency virus (HIV) coinfected subjects, HCV viremia is associated with an inflammatory CD4 Th17 phenotype in the absence of an HIV control (26). Moreover, increased circulating Th17 cells correlated inversely with CD4 T cell percentages and absolute counts and directly with CD4 T cell immune activation in HIV discordant patients (impaired restoration of CD4 T cell counts despite prolonged suppression of plasma viremia) (85). Finally, a similar association has been highlighted in other viral diseases. IL-17 has been shown to play a significant role as a proinflammatory cytokine dealing with cytomegalovirus in patients with liver and other organ transplants (2,29). Finally, IL-17A-producing cells may influence the pathophysiology of chronic active Epstein-Barr virus (65).
Classification of COVID-19 Stages
Recently, Siddiqi and Mehra (81) suggested dividing the course of the disease into three clinical stages. Each clinical stage is correlated with the severity of the illness. The first stage (i.e., mild−early infection) is characterized by an antiviral response; the second (i.e., moderate−pulmonary involvement without/with hypoxia), by a decrease in the antiviral response and an increase in the proinflammatory response with viral multiplication and inflammation located in the lungs as a norm; and the third (i.e., severe−systemic hyperinflammation), by a severe inflammatory response. Taking the clinical experience accumulated to date into account, a fourth phase could be added. This represents those convalescent patients who develop late complications (e.g., organizing pneumonia), and those who develop chronic sequelae (e.g., pulmonary fibrosis). For details see Figure 1.
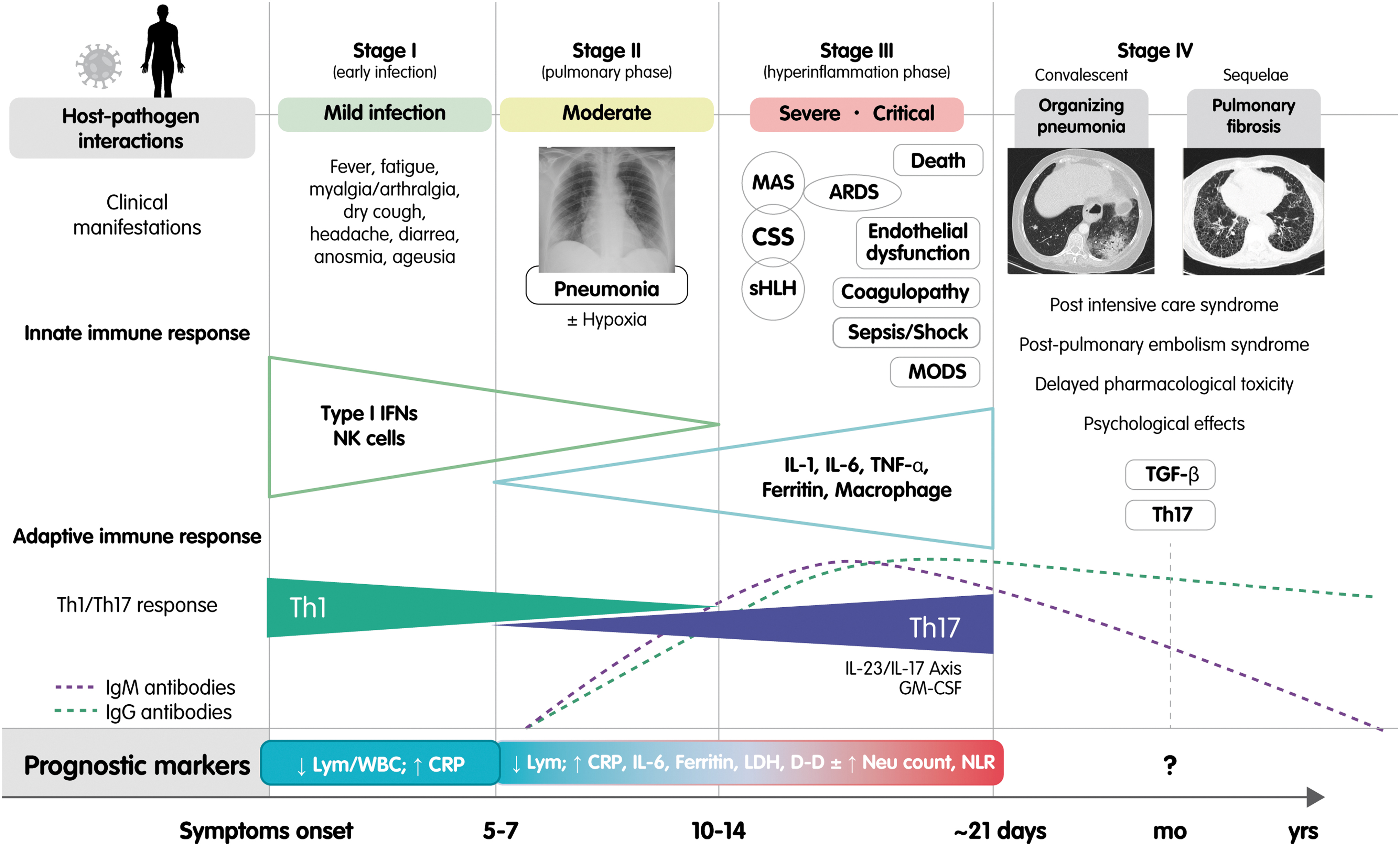
Classification of coronavirus disease 2019 stages. Adapted from the initial clinical–therapeutic staging proposal by Siddiqi and Mehra (81). This figure shows a correlation between the stages of the disease, the severity, the main clinical manifestations, and the innate and humoral immune response. The classical and more recently described prognostic markers are also depicted. The first stage is characterized by an antiviral response to severe acute respiratory syndrome coronavirus 2, the second by a decrease in the antiviral response and an increase in the proinflammatory response with viral multiplication and inflammation located in the lung, the third by an inappropriate and exaggerated Th17-driven inflammatory response, and the fourth by late complications associated with the infection and its treatment. The role of the neutrophil is crucial in the polarization of the Th17 response. ARDS, acute respiratory distress syndrome; CRP, C-reactive protein; CSS, cytokine storm syndrome; D-D,
In the context of a Th1 polarized adaptive response toward SARS-CoV-2, available data show that patients, particularly those with severe disease, display increased plasma levels of IL-17 and TNF-α (36,94). Therefore, an enhanced polarization during phase II toward Th17 cells and IL-17-related pathways seems to be associated with a worse outcome of the disease (27). This change is owing to several mechanisms including the interference of viral proteins in the antiviral response of the host cells, that is, the inhibition of the production of IFN and the consequent increase in proinflammatory cytokines and the involvement of neutrophils. Several abnormal hematological parameters have been reported in COVID-19 patients including lymphopenia, neutrophilia, and elevated levels of
To date, several authors have proposed neutrophil count and a high NLR as prognostic markers in COVID-19 patients (40,96). NLR is an independent risk factor for in-hospital mortality for COVID-19 patients, especially for men (48). A recent meta-analysis (43) that included 828 patients demonstrated that the NLR values increased significantly in COVID-19 patients with severe disease. Furthermore, the China Expert Medical Treatment Group for COVID-19 developed a risk score to predict development of critical illness in a cohort of 1,590 patients. They identified 10 independent predictors (including NLR) and developed a risk score (COVID-GRAM) that predicts the development of critical illness (46).
Phase III of the disease is the result of a defective antiviral and inflammatory response to SARS-CoV-2 in a susceptible host. A proinflammatory background is favored in part by the Th17 polarization. The “cytokine storm” is both ineffective with respect to the pathogen and detrimental for the body and eventually leads to multisystem complications such as secondary hemophagocytic lymphohistiocytosis, ARDS, endothelial dysfunction, coagulopathy, multiple organ dysfunction syndrome, and finally to death (14,35,97).
Therapeutic Implications
The natural history model of acute COVID-19 is important because it not only makes the correlation of the immune response with the clinical course of the disease possible but also directs the available treatments based on the stage of the disease. Thus, antiviral drugs are recommended mainly during stage I, and anti-inflammatory drugs should be used from stage II to III (100).
In this context, it is worth noting that the action of drugs involved in the Th17 response may be relevant in the treatment of SARS-CoV-2. Blocking the IL-23/IL-17 axis using targeted biological drugs that are already available and used to treat different inflammatory and immune disorders may be a good therapeutic option (76). There are also drugs that act indirectly on this pathway such as convalescent plasma therapy, Colchicine, and JAK inhibitors.
In the face of the COVID-19 pandemic, there are limited data about whether patients with inflammatory diseases will be more susceptible to infection or disease. In recent reports, some patients with SARS-CoV-2 treated with IL-23/IL-17 axis inhibitors (e.g., guselkumab and secukinumab) have shown a favorable outcome even in the presence of associated risk factors for severe disease (6,27,54). IL-23 does not seem to be essential for controlling virus clearance but may play a role in the deleterious hyperinflammatory state associated with severe symptoms (6). A clinical trial studying the use of Ixekizumab (an IL-17A inhibitor) associated with antiviral therapy is currently ongoing in China as a possible treatment for COVID-19 (19). One of the possible explanations for this behavior comes from studies of patients with inflammatory bowel disease in which medical therapy (including ustekinumab, an anti-IL-12/23 inhibitor) has been associated with a lower expression of ACE2 in the ileum and colon (12). Mavrilimumab (61) and gimsilumab (59), monoclonal antibodies against GM-CSF, are currently undergoing clinical trials in patients with severe COVID-19 pneumonia and hyper-inflammation (92). These drugs could be a promising therapeutic target in the near future.
Some studies have shown that intravenous immunoglobulin (IVIg) reduces the proliferation of Th17 cells and decreases the production of IL-17A, IL-17F, IL-21, and CCL20. In addition, IVIg appeared to modulate the Th17/Treg ratio that is associated with some immune-mediated diseases. These studies point to one of the plausible mechanisms in the treatment of COVID-19 with convalescent plasma (75).
Colchicine is a molecule that is commonly used to treat several autoinflammatory diseases including gout, familial Mediterranean fever, and Behçet's disease as well as cardiac conditions, for example, viral pericardial syndromes (20,24). Its usefulness is largely explained by its pleiotropic anti-inflammatory and immunomodulatory properties. Colchicine decreases the production of proinflammatory cytokines promoted by IL-17 such as IL-1, IL-6, and TNF-α as well as chemotaxis of inflammatory cells, for example, neutrophils and monocytes, to the inflammation sites. Recently, Colchicine has also been associated with disrupting NLRP3 inflammasome activation, thereby suppressing subsequent release of IL-1β and IL-18 (52).
There is other basic research that demonstrates the effect of Colchicine in RSV replication and suppression of secondary airway inflammation given the fact that Colchicine facilitates the expression of IFN-α and IFN-β1 and regulates anti-oxidative factor production (4,49). In addition, there is a wide range of preclinical and clinical literature on the inhibiting effect of Colchicine on viral diseases such as adenovirus (11), herpes simplex virus type 1 (9), Epstein-Barr Virus (64), hepatitis virus (5,30), and so on. Recently a patient with cardiac tamponade secondary to COVID-19 was treated with Colchicine in addition to corticosteroids and antimalarials with a good clinical response (22). Therefore, some researchers thought it would be wise to test Colchicine in the context of COVID-19 (24,68).
Five patients (age 38–61 years) under treatment with Colchicine for iatrogenic allogenosis 1–3 weeks before testing positive for COVID-19 has been reported recently. Despite the presence of several comorbidities, including arterial hypertension, and type 2 diabetes, they developed mild symptoms (headache, cough without dyspnea, and arthralgias). It should be noted that some close contacts presented severe symptoms and three of them died (57).
Since March 26, 2020, 13 studies have been registered at
Randomized Controlled Trials Registered in
Data updated as of May 22, 2020.
BID, twice a day; BNP, brain natriuretic peptide; CKD, chronic kidney disease; COVID-19, coronavirus disease 2019; CRP, C-reactive protein; CT, computed tomography; ESR, erythrocyte sedimentation rate; GFR, glomerular filtration rate; ICU, intensive care unit; IL, interleukin; IMV, invasive mechanical ventilation; LDH, lactate dehydrogenase; LPV/r, lopinavir/ritonavir; LVEF, left ventricular ejection fraction; NEWS, national early warning score; QD, one a day; PCR, polymerase chain reaction; p.o., per os; SOC, standard of care; SOFA, sequential organ failure assessment; TID, three times a day; WHO, World Health Organization.
The use of JAK inhibitors to target IL-6 and other cytokines (e.g., IL-17) with JAK-dependent signaling is one way to restrain the excessive level of cytokine signaling in COVID-19 patients (82). Wu and Yang (92) demonstrated that fedratinib (a JAK2 inhibitor) can suppress the production of several Th17 signature cytokines in a murine model. Therefore, this is promising and could prevent the deteriorating outcomes of Th17-associated cytokine storms in COVID-19. Finally, an open prospective randomized trial has recently been started with mild and severe COVID-19 patients that will be randomized into colchicine, ruxolitinib, secukinumab, and control groups (60). We hope that the results of these studies may be available soon.
Conclusions
An extensive amount of information has been collected over the past two decades on the molecular immune pathogenesis of coronavirus infections based on understanding the SARS-CoV-1 and MERS-CoV during those epidemics. This has been helpful in the current SARS-CoV-2 pandemic but much remains to be learned about the potential pathogenesis mechanism of SARS-CoV-2. Gaining knowledge on this topic will allow for better insight into the different clinical phases and the development of new relevant therapeutic strategies for controlling it. In the meantime, until a new, effective vaccine is designed and/or neutralizing antibodies that can stop viral spread are induced, the mechanisms of action of the drugs used to address similar pathological conditions such as those that dysregulate IL-17 signaling need to be rethought. This way, pilot clinical trials can be carried out that will quickly show us their potential therapeutic efficacy on this viral pathogen that is taking so many lives daily and destabilizing the world economy. A novel, additional strategy for dealing with SARS-CoV-2 could be to use biological drugs that are already available and are able to block the IL-17 pathway both directly and indirectly.
Author's Contributions
All authors contributed to the conception of the study, wrote the article, and approved the article before submission.
Footnotes
Acknowledgments
The authors recognize Cecile Dunn and Cristhian Ramírez-Parra for their technical support concerning the English language and the Internal call for improving English scientific manuscripts from the Fundación Universitaria de Ciencias de la Salud (FUCS). The authors also recognize Jeyson Arismendy for the help.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.