
Abstract
Eosinophils participate in the immune response against many pathogens, including viruses. Since mouse eosinophils are susceptible to influenza A virus infection and possess antiviral activity, we evaluated the expression of sialic acid residues in human eosinophils and their response against influenza virus in vitro. We demonstrated that human eosinophils express α2,6- and α2,3-linked sialic acid, and drastically reduced influenza virus titer. After influenza virus exposure, eosinophils upregulated retinoic acid-inducible gene I (RIG-I) mRNA expression, but no other pattern recognition receptors. Finally, high concentrations of interleukin-8 (IL-8) were found in influenza virus-exposed eosinophil cultures. These data suggest that human eosinophils possess antiviral activity and may play a role in the innate immune response to influenza virus.
Introduction
Influenza A virus (IAV) causes seasonal epidemics that result in ∼3–5 million cases of severe respiratory illness and 290,000–650,000 deaths worldwide per year (14), as well as periodic pandemics that may even induce higher morbidity and mortality (13). Influenza viruses are enveloped negative-sense single-stranded RNA viruses that belong to the Orthomyxoviridae family (28). Influenza hemagglutinin (HA) binds to sialic acid residues expressed on host cells' surface, thereby mediating virus entry. Avian IAVs preferentially bind to α2,3-linked sialic acid (SAα2,3), whereas human IAVs preferentially bind to α2,6-linked sialic acid (SAα2,6) (18).
Influenza, among other respiratory viral infections, triggers asthma exacerbations (26). TH2-high endotype is the most prominent asthma and is characterized by eosinophilic inflammation in the airways (1). Eosinophils participate in asthma pathogenesis through eosinophil-derived toxic mediators, including basic proteins, cytokines, and chemokines (16). Since respiratory viruses are associated with asthma exacerbations, several studies have evaluated the role of eosinophils against these pathogens (9). Eosinophils express pattern recognition receptors (PRRs) that bind to viral molecules, including toll-like receptor (TLR)3 (23), TLR7, TLR9 (38), retinoic acid-inducible gene I (RIG-I), and melanoma differentiation-associated gene-5 (MDA-5) (20). They also possess proteins with antiviral activity, such as eosinophil cationic protein (ECP) and eosinophil-derived neurotoxin (EDN) (29), and preformed typical type-1 cytokines, such as interleukin (IL)-2, IL-12, and interferon (IFN)-γ (4). Moreover, eosinophils are able to present viral peptides to virus-specific T CD4+ (10) and T CD8+ cells (31).
During the 2009 influenza pandemic, it was reported that patients with asthma had less severe outcomes related to viral infection compared with patients with other comorbidities (36). Samarasinghe et al. demonstrated that mouse eosinophils are susceptible to IAV infection and respond to virus through degranulation, being able to induce CD8+ T cell antigen-specific proliferation (31). More recently, Sabogal Piñeros et al. showed that human eosinophils are able to capture and inactivate influenza virus in vitro (30).
Despite the evidence of an antiviral role of eosinophils against respiratory viruses recently reviewed (9), there is a lack of data about the involved mechanisms. The aim of this study was to evaluate the presence of sialic acid in human eosinophils and their response to IAV exposure. We found that human eosinophils express SAα2,6 and SAα2,3. IAV-exposed eosinophils reduced the viral titer, upregulated RIG-I expression, and secreted IL-8.
Methods
Cells and virus
MDCK cells (NBL-2) (ATCC, Manassas, VA) were grown in Eagle's minimum essential medium supplemented with 10% fetal bovine serum (FBS). A549 cells (ATCC) were grown in F12K medium containing 10% FBS and 2 mM
Healthy human eosinophil isolation and culture
The Bioethics Committee of the School of Medicine (Universidad Autónoma de Nuevo León, Monterrey, México) approved the study, and all donors provided written informed consent (IN18-00003). Eosinophils were isolated from freshly drawn peripheral blood collected from nonatopic healthy donors (eosinophils <350 cells/μL). A leukocyte-rich plasma layer was obtained by adding 6% Dextran. Red blood cells were lysed by hypotonic shock. Eosinophils were isolated by density centrifugation using Ficoll-Paque (GE Healthcare, Uppsala, Sweden) and negative selection (Miltenyi Biotec, Auburn, CA). Eosinophil purity was confirmed by cytospin, Wright staining (>99%), and flow cytometry using antibodies anti-Siglec-8 (allophycocyanin [APC]; Biolegend, San Diego, CA) and anti-CCR3 (fluorescein isothiocyanate [FITC]; Biolegend, San Diego, CA). Viability (>95%) was assessed by trypan blue exclusion.
Eosinophils (600,000 cells/well) were cultured in 12-well plates and incubated with IAV (multiplicity of infection [MOI] 0.1 or 1), R848 (10 mM; Novus Biologicals, Centennial, CO), or free medium for 1 h. After washing to remove unbound virus, cells were maintained in fresh medium (Roswell Park Memorial Institute [RPMI] 1640 medium, 10% FBS, 2 mM
Determination of sialic acid residues
MDCK, A549, and eosinophils were stained for SAα2,6 and SAα2,3 determination and assessed by fluorescence microscopy and flow cytometry as previously described with some modifications (32). In brief, detached MDCK and A549 cells or eosinophils were fixed in 2% formaldehyde for 30 min at 4°C and washed with flow buffer (1% bovine serum albumin in PBS). The cells were then stained with 20 μg/mL biotinylated Maackia amurensis lectin-II (MAA II; Vector Laboratories, Burlingame, CA) to detect SAα2,3 or 20 μg/mL fluorescein Sambucus nigra lectin (SNA; Vector Laboratories) for 1 h at 4°C to identify SAα2,6. For sialic acid α-2,3, a second staining with APC-conjugated streptavidin (BD biosciences, Mountain View, CA) diluted 1:250 in flow buffer was performed for 1 h at 4°C. Cells were washed with flow buffer, acquired on an FACS Canto II flow cytometer, and analyzed using FlowJo version 10 for Windows (Treestar, Ashland, OR). For fluorescence microscopy, the cells were further stained with 4′,6-diamidino-2-phenylindole (DAPI) to counter-stain cell nuclei and then observed using a fluorescence microscope (Thermo Fisher Scientific, Carlsbad, CA).
Virus titer
Eosinophils (850,000 cells/well) were cultured in 12-well plates and incubated in fresh medium (RPMI, 10% FBS, 2 mM
Viability assay
Cell viability of cultured eosinophils exposed or not to IAV was analyzed using Calcein Blue AM (BD biosciences) by flow cytometry following manufacturer's protocol.
Gene expression
Total RNA was isolated using RNeasy Mini kit (QIAGEN, Valencia, CA) and stored at −80°C until use. The concentration of RNA in each sample was measured by spectrophotometry and was reverse transcribed with M-MLV (New England BioLabs, Ipswich, MA) and random hexamers (Agilent Technologies, La Jolla, CA). Real-time polymerase chain reaction (PCR) was performed using iQ SYBR Green Supermix or IQ Supermix (Biorad, Hercules, CA), specific primers (Alpha DNA, Montreal, Quebec, Canada), or Taqman probes (Applied Biosystems, Grand Island, NY) in a CFX96 thermocycler (Biorad) to evaluate relative mRNA expression of TLR3, TLR7, DDX58 (RIG-I), IFNB1, and IFNL1 (IFN-λ1, Hs00601677g1). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used to normalize the expression levels. Primer sequences used were GAPDH F (TGCCAAATATGATGACATCAAGAA), GAPDH R (GGAGTGGGTGTCGCTGTTG) (5), IFNB1 F (CAGCAATTTTCAGTGTCAGAAGC), IFNB1 R (TCATCCTGTCCTTGAGGCAGT) (12), TLR3 F (AAATTAAAGAGTTTTCTCCAGGGTGTT), TLR3 R (ATTCCGAATGCTTGTGTTTGC) (33), TLR7 F (TTGAGAAGCCCTTTCAGAAGTC), TLR7 R (CACCTGACTATAGGCCACATGA) (17), RIG-I F (CCAAGCCAAAGCAGTTTTCAAG), RIG-I R (CACATGGATTCCCCAGTCATG) (33).
Cytokine measurement
IL-8, IL-6, and IFN-γ secretions were measured from culture supernatants by enzyme-linked immunosorbent assays (ELISAs) (Thermo Fisher Scientific, Vienna, Austria) following manufacturer's protocol.
Statistical analysis
Significance was determined using one-way analysis of variance (ANOVA) and Student's t-test or Bonferroni post hoc test or nonparametric Kruskal–Wallis analysis and Dunn post hoc test. Calculations were performed using GraphPad Prism version 4.0 for Windows (San Diego, CA). p-Values <0.05 were considered significant.
Results
First, we determined the purity of the isolated human eosinophils through the expression of Siglec-8 and CCR3 by flow cytometry (Supplementary Fig. S1B). Isolated cells were also morphologically identified by Wright staining (Supplementary Fig. S1C). The purity obtained in all experiments was >99%.
Since influenza virus binds to sialic acid residues expressed on cell surfaces, we determined the presence of SAα2,6 and SAα2,3 on human eosinophils. It is reported that both MDCK and A549 express sialic acid residues on their surface (32), therefore, we used these cell lines as positive controls. Using specific lectins and fluorescence microscopy, we found that human eosinophils express SAα2,6 and SAα2,3 on their surface (Fig. 1A, B), as cells showed fluorescent staining similarly to that observed in MDCK and A549 cells (Fig. 1C–F). Presence of SAα2,6 and SAα2,3 on eosinophil surface was confirmed by flow cytometry (Fig. 1G). Because it is reported that human eosinophils reduce virus infectivity (6), we investigated whether human eosinophils exert antiviral activity against IAV. We exposed purified eosinophils to IAV (MOI 1) for 24 h and assessed viral titer. We found that viral titer diminished drastically after eosinophil exposure (Fig. 1H). To rule out the possibility that the decrease in viral titer was due to nonviable eosinophils, we performed viability assays at 24 h. The eosinophils cultured alone presented 65.9% of viability that is expected due to the short life span of eosinophils from blood (Supplementary Fig. S2A). In contrast, eosinophils exposed to IAV presented 92.3% of viability (Supplementary Fig. S2B), which suggests that eosinophils are activated by IAV.

Human eosinophils express sialic acid residues and reduce viral titer. Freshly isolated eosinophils
Eosinophils express a great number of PRRs (19), including receptors that recognize viruses (9). Since mouse eosinophils upregulate some of these PRRs after IAV infection (21), we evaluated whether human eosinophils had a similar behavior. Eosinophils upregulated gene expression of DDX58 (RIG-I) after IAV (MOI 0.1) infection and the increment was statistically significant using a higher viral dose (MOI 1) (Fig. 2A). TLR3, TLR7, and IFNB1 (IFN-β) expression levels were not upregulated after IAV exposure (Fig. 2B–D). IFNL1 (IFN-λ1) was not expressed by peripheral blood eosinophils (data not shown).
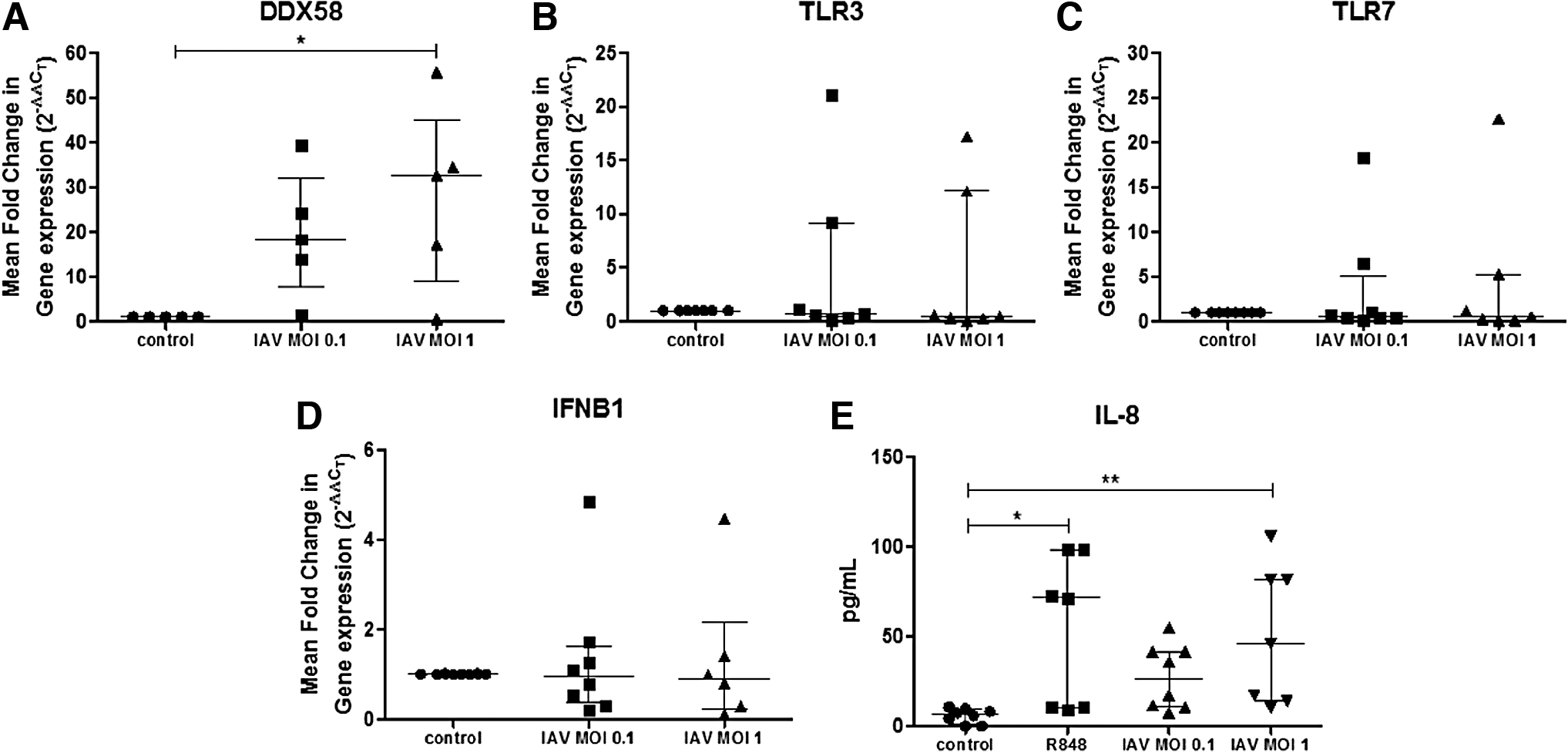
Human eosinophils upregulate the gene expression of RIG-I and secrete IL-8 after IAV exposure. Human eosinophils were isolated and stimulated with IAV (MOI 0.1 or MOI 1) for 24 h followed by RNA extraction and RTqPCR. Data are represented as the median and IQR of n = 5–8 donors/group. Statistical significance determined by one-way ANOVA and Bonferroni post hoc test (*p < 0.05)
Eosinophils respond to viral infection through cytokine release. Because TLR7 ligands activate eosinophils (25) and induce IL-8 secretion (22), we used R848, a synthetic TLR7 and TLR8 ligand, as positive control. As expected, eosinophils stimulated with R848 secreted IL-8 (Fig. 2E). Interestingly, IAV-exposed eosinophils also secreted IL-8. Finally, we evaluated IL-6 and IFN-γ amount in supernatants, but eosinophils did not secrete detectable levels of these cytokines after IAV exposure (data not shown).
Discussion
IAV HA protein binds to sialic acid present on target cell surfaces and favors virus attach and entry to the cells. IAV that infects humans binds preferentially to SAα2,6, whereas avian IAV infects preferentially cells expressing SAα2,3 linkages (18). In this study, we demonstrate that human eosinophils express both SAα2,6 and SAα2,3, thus IAV may bind to eosinophils through these receptors. Interestingly, it has been reported that neutrophils do not express SAα2,6 or SAα2,3; nevertheless, they are susceptible to IAV infection (40).
In our experiments, eosinophils were able to decrease the viral titer. Similar results have been observed with parainfluenza virus (6). A recent study showed that mouse eosinophils are able to decrease IAV infectivity, and this effect was mediated by EDN and ECP combined (34). These proteins are RNases with antiviral activity against ssRNA viruses, such as respiratory syncytial virus (RSV) (29). A similar mechanism could be occurring in human eosinophils. We also observed that eosinophils exposed to IAV maintained their viability in contrast with eosinophils cultured alone. It has been demonstrated that TLR7 ligands, such as R848, are able to activate eosinophils preventing cell death (25). It is possible that eosinophils are recognizing IAV through TLR7, thus being activated and inducing cell survival.
Human eosinophils express PRRs that recognize different viral molecules; among them, TLR7 is the most abundant (19). These cells respond through TLR7 signaling by the expression of the activation marker CD11b, the release of superoxide (25), and the secretion of IL-8 (22). In our study, eosinophils responded to R848, a TLR7 and TLR8 ligand, secreting IL-8. In a similar way, human eosinophils incubated with IAV also secreted IL-8. IL-8 is a potent human neutrophil chemokine that is mainly secreted by epithelial cells after IAV infection (35). Although eosinophils normally do not express the IL-8 receptor (27), activated eosinophils from atopic donors respond to IL-8 with chemotaxis (37). It is possible that in an asthmatic scenario, lung eosinophils could respond to IAV secreting IL-8 and recruiting even more eosinophils to the site of infection.
It has been shown that IL-6 is released by mouse eosinophils after pneumonia of mice virus (PVM) infection and by human eosinophils after RSV infection (7). Human eosinophils incubated with IAV also secrete IL-6 (30). However, in our experimental conditions, IL-6 was not detected in supernatants of cultured eosinophils after IAV exposure. A possible explanation for this difference may be related to the methodology used in both studies, including dose of infection, time of incubation, and the IAV strain used. Eosinophils contain preformed IFN-γ and other type-1 cytokines within their granules, which can be released after stimulation (3). In our study, IAV-exposed eosinophils did not release IFN-γ. Our results are in agreement with a previous report studying the eosinophil response to PVM infection, showing absence of IFN-γ secretion (7). Nevertheless, eosinophils pulsed with IAV peptides induce the production of this cytokine by IAV-specific T CD8+ cells, which suggests T cell regulation by eosinophils (31). The secreted IFN-γ by T CD8+ cells could activate eosinophil to further respond to IAV, as IFN-γ increases the expression of TLR7 and TLR8 (6,25), and potentiates TLR7-induced nitric oxide production by eosinophils (6).
We demonstrate that human eosinophils incubated with IAV upregulate RIG-I mRNA expression, as found in mouse eosinophils by other authors (21). RIG-I induces an antiviral response against RNA viruses, such as IAV (2,15). Its activation induces types I and III IFN and inflammatory cytokine production (39), and it has been proposed as the primary PRR for IAV recognition (2). Whether the overexpression of RIG-I in eosinophils has a biological significance or not has to be further investigated, since stimulation of this receptor by a synthetic ligand did not activate these cells (20). Although the expression of TLR3 and TLR7 was not increased after IAV exposure in our study, their constitutive expression in human eosinophils suggests a participation in viral infections. Unlike intestinal eosinophils (11), peripheral blood eosinophils did not express IFN-λ1. Furthermore, IFN-β1 was not upregulated after IAV incubation. All together, these results suggest that the antiviral role of eosinophils do not require IFNs. This could be related to the diminished production of both IFN-β1 and IFN-λ1 by bronchial epithelial cells after coculture with eosinophils (24). Moreover, several observations suggest that asthmatic patients have an impaired innate immunity against respiratory viruses because of a reduction of types I and III IFNs production (8).
In summary, sialic acid residues are present in human eosinophils from healthy individuals, suggesting a potential ligand for IAV entrance to these cells. IAV-exposed eosinophils are able to reduce the viral titer. Besides, human eosinophils incubated with IAV pmd09 induce IL-8 secretion and RIG-I gene upregulation. These results strongly suggest an important role of eosinophils in IAV infection, and their relevance in clinical conditions such as asthma.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by the Consejo Nacional de Ciencia y Tecnología (CONACYT) Ciencia Basica grant 253330, and the Programa de Apoyo a la Investigación Científica y Tecnológica (PAICYT) grant SA805-19. Armando S. Flores-Torres received a PhD scholarship from CONACYT (558124).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.