
Abstract
Persistence of hepatitis B virus (HBV) infection leading to chronic infection and its sequalae is responsible for over half a million deaths worldwide. The reason for persistence of chronic hepatitis B (CHB) infection is still not clearly understood. An attempt was made to understand the role of immune regulatory genes in CHB in comparison to spontaneously cleared HBV infection. Relative gene expression of 26 genes involved in innate immunity were studied using Real-Time Polymerase Chain Reaction Array. A total of 679 subjects from three different geographical regions of Northeast India (Assam, Arunachal Pradesh, and Tripura) were included in this case–control study. The cases were subdivided into CHB cases with HBeAg(+)(72), CHB with HBeAg(−)(278), spontaneously cleared controls (88), and healthy controls (228). Overall, 28.3% of the subjects had previous exposure with HBV, while 28.6% had protective antibodies IgG/IgM against HBV. There was a statistically higher number of CHB in men (66.4%) compared to women (33.6%) (p = 0.0001). Proto-oncogene FOS has been found to be moderately upregulated in CHB with HBeAg +ve (2.3-fold) and significantly upregulated (4.1-fold upregulation) in hepatocellular carcinoma. Further, FOXP3 was found to be significantly upregulated (3.0-fold, p = 0.01) in CHB with HBeAg (+) compared to spontaneously cleared HBV infection. In conclusion, CHB with HBeAg positivity was found to have disrupted immune response with upregulation of FOS and FOXP3. Thus, early induction of HBeAg seroconversion with interferon-based therapy or oral nucleos(t)ide analogs along with FOS inhibitors can have important clinical implications in the management of CHB and preventing cirrhosis and HCC.
Introduction
Chronic hepatitis B (CHB) and its sequelae of liver cirrhosis and hepatocellular carcinoma is a major public health problem worldwide (23). The current global burden of hepatitis B virus (HBV) is 360 million chronically infected people and accounts for over half a million deaths from HBV-related liver disease or hepatocellular carcinoma (HCC) (13). In most adults about 75% of HBV infection is subclinical, ∼5% to 10% of adults develop a chronic infection manifested by the persistence of hepatitis B surface antigen (HBsAg) in serum along with the production of other viral antigens and HBV DNA (21).
Children infected during the first few months of life almost invariably develop chronic infection. The chronicity of HBV infection is not yet properly understood. After acute HBV infection, whether the patient will develop CHB or clear the virus is largely dependent on the immune response of the host. In recent years, the role of innate immunity, has come to light on many infectious diseases. Like other viruses, the outcomes of HBV infections and pathogenesis of the associated diseases are determined by virus–host interactions, largely mediated by innate and adaptive immune responses (28). Virus infection elicits a rapid and potent innate immune response in mammalian cells to produce proinflammatory cytokines and chemokines that limit virus replication and coordinate adaptive antiviral immunity (19).
Some studies have demonstrated that HBV carriers have a higher frequency of regulatory T cells (Tregs) in peripheral blood and liver than healthy controls or individuals with resolved infection, indicating that Tregs may contribute; to HBV persistence (19,28). However, other studies have failed to detect any differences in Treg frequencies between asymptomatic carriers (AsCs) and healthy controls (5). Thus, the role of Tregs in the progression of liver disease remains controversial, and little is known about whether their potential roles in viral hepatitis pathogenesis differ according to their functional activities.
The need for the study of the innate immune process involved in HBV pathogenesis and persistence is immense. In India, it is seen that there are pockets of high HBsAg cases especially among isolated tribal communities (17,20). A study from this Institution in 2005 found a high prevalence of HBV infection (>90%) and over 20% HBsAg in an isolated tribal community of Arunachal Pradesh (1). The proposed study area selected is associated with a high prevalence of delta hepatitis virus (35.5% among HBsAg positives) and multiple HBV genotypes (2,3). With this background, this study was therefore designed to understand the role of immune regulatory genes in CHB in comparison to spontaneously cleared HBV infection in HBV endemic region of northeast India.
Methodology
The study was approved by the institute scientific advisory committee and funded by ICMR.
A total of 679 subjects from three different geographical regions of Northeast India (Assam, Arunachal Pradesh, and Tripura) were included in this hospital-based case–control study. The study was conducted from 2015 to 2019. Both male and female subjects attending Assam Medical College & Hospital and nearby hospitals for treatment, from the HBV hyperendemic area of Arunachal Pradesh (Dibang Valley) and subjects attending Agartala Government Medical College, Tripura were enrolled. The subjects were broadly divided into cases and controls based on enrolment criteria. Cases included CHB patients who are HBsAg Positive >6 months and without antiviral treatment for the past 6 months and with no other recent clinical infections 2 weeks prior enrollment (N = 350).
Cases were further subdivided into two categories- Group 1 cases included chronic HBV patients (HBsAg +, HBeAg −, HBcAb IgG +, and HBsAb+) (N = 278), Group 2 cases included chronic wild-type HBV (HBsAg +, HBeAg +, HBcAb IgG +, and HBsAb −) (N = 72). The control group (N = 316) was divided into two categories—Group 3 control included spontaneously cleared HBV: (HBsAg -, HBcAb +, and HBsAb +) (N = 88), and Group 4 control included age and gender-matched healthy controls with normal liver function test and no history of liver disease (HBsAg− and HBcAb −) (N = 228). Group 5-Chronic HBV patients with Hepatocellular carcinoma (HBsAg + with Hepatocellular carcinoma) were also included as a sub-category during the analysis (N = 13).
Written informed consent was taken from all individuals before sample collection. Ethical approval was obtained from the Institutional Ethical Committee of ICMR-RMRC, NE region, Dibrugarh before the commencement of the study.
Venous blood 2.5 mL was collected in a K3EDTA tube and 2.5 mL blood in Paxgene Blood RNA collection tube (cat. no. 762165, BD Bioscience, NJ). All samples were screened for HBsAg and HBcAb, using a commercially available ELISA kit (Monolisa HBsAg ULTRA by BIORAD) and HBcAb ELISA kit (DIA.PRO, Milano, Italy) according to the manufacturer's instructions. All samples found to be negative for HBsAg were further tested for anti-HBs, HBeAg, and anti-HBeAg using HBsAb ELISA Kit (DIA.PRO), HBeAg, and Ab ELISA Kit (DIA.PRO) respectively.
HBV DNA viral load was performed in 292 HBV DNA positive samples using HBV viral load kit (cat. no. P/N: 03500756 190; Roche Diagnostics, Mannheim, Germany) in Cobas TaqMan 48 real-time polymerase chain reaction (PCR) machine (Roche Diagnostics) following manufacturer's protocol. HBV DNA low positive, high positive, and negative control was used with every run. The detection limit of the system is from 6 IU/mL (36 copies/mL) to 1.10E+8 IU/mL.
For the relative gene expression study, a subset of cases and controls were selected through simple randomization. Total RNA from lysed cells was extracted using Paxgene Blood RNA extraction kit (cat, no./ID: 762174; Qiagen, Hilden, Germany) according to the manufacturer's instructions. cDNA was prepared using the first-strand cDNA synthesis kit from Qiagen (cat. no. 205311; QIAGEN AG, Basel, Switzerland). Relative gene expression of 26 genes (FOS, FOXP3, FOXP1, GZMB, CD274, IFNAR2, IFNG, IRF3, IRF7, MYD88, TICAM1, BTLA, CTLA4, CD40LG, CD28, CD40, TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, and TLR10) engaged in innate immunity was performed using custom-designed PCR array plates from SABiosciences (QIAGEN AG).
The customized PCR array for this study was designed to run 3 samples at a time (32 different gene per 96-well PCR plate i.e., 26 genes of interest +3 housekeeping genes +1 reverse transcriptase control, 1 genomic DNA control [HGDC], and 1 positive PCR control [PPC]). A 25–50 ng of cDNA was mixed with 2 × RT2 SYBR Green Mastermix ((Qiagen) and added to the PCR array. Real-time PCR was performed in ABI step one plus real-time PCR machine (ABI stepone plus, ThermoFisher Scientific, Waltham, MA). The thermal cycling condition was initial denaturation at 95°C for 15 min followed by 40 cycles of denaturation at 95°C for 15 sec and annealing at 60°C for 1 min. The amplification was followed by a melt curve at 60°C to 95°C. Data were collected in every increment of temperature of 0.3 sec interval.
Housekeeping genes (ACTB, B2M, and GAPDH) were used to normalize the samples. A PPC, provided by the manufacturer, which consists of a predispensed artificial DNA sequence was used to test the efficiency of the PCR. The Ct-value for the PPC as per manufacturer's should be 20 ± 1 cycle. To minimize plate to plate variations only samples with PPC 20 ± 1 cycle were included in the analysis for relative gene expression study. Relative gene expression analysis was performed online by using the available online platform provided by SABioscience for gene expression studies.
Data collected were subjected to descriptive statistical analysis using the SPSS version 20 (SPSS, Inc., IL). Quantitative variables were summarized using mean and standard deviation (SD) while qualitative variables were summarized in frequencies and proportions. The level of significance was set at 5% (p ≤ 0.05). Relative gene expression fold change was illustrated using histogram plots in Graphpad Prism. version 5.0 for Windows, (GraphPad Software, La Jolla, CA).
Results
Demographic and clinical characteristics of the study subjects
A total of 679 subjects were recruited for the study. The majority of the subjects recruited were from Arunachal Pradesh (317) followed by Assam (186) and Tripura (176). The subjects were screened for HBsAg and accordingly 350 CHB patients and 316 healthy controls were included in the study. Acute HBV cases (13 nos.) were excluded from the analysis.
The overall age of the study subjects (both cases and controls) ranged from 3 to 85 years with an intraquartile age range of 23.5 to 45 years and a mean age of 34 years (SD ±15.0). The mean age of cases was 35 years and controls were 32.6 years. Overall, men accounted for 56.6% of subjects (384/679). Among the CHB cases, there was a statistically higher number of men compared to women (66.4% men compared to 33.6% women; p = 0.0001) whereas among controls there was a higher frequency of women (53.7% compared to 46.3% men).
Among cases, 278 cases were CHB (HBsAg positive and HBeAg negative) and 72 cases were CHB wild-type (HBsAg positive and HBeAg positive). Among controls, 228 were healthy controls (HBsAg negative and HBcAb negative) and 88 were spontaneously cleared group (HBsAg negative, HBcAb positive) (Fig. 1).

Schematic diagram of the study subjects enrolled.
Overall Hepatitis B core antibody (HBcAb) prevalence was found to be 28.3% among the controls. The prevalence of HBcAb increased with age from 20% in the under 15 years age group to 37.9% over the 45 years age group (p = 0.1). HBcAb prevalence was similar between both genders (28.1% vs. 28.5%). However, there was a statistically significant difference in the prevalence of HBcAb among the three states of Northeast India. Arunachal Pradesh had the highest prevalence of 31% followed by Tripura (30.5%) and Assam (8.8%) (p = 0.02).
The protective antibody (HBs Ab >10 mIU/L) against HBV was measured in 298 healthy controls from samples collected from the three states (Assam, Tripura, and Arunachal Pradesh). Overall, 28.6% of the subjects had immunity for HBV infection. The immunity status was highest in Arunachal Pradesh (34.7%) compared to Tripura (18.3%) and Assam (17.6%), which was statistically significant (p = 0.002). This may be due to a higher frequency of natural infection in Arunachal Pradesh. Women had a higher prevalence of HBcAb (30.8%) compared to men (26%). Younger subjects (<15 years) had a higher prevalence of protective antibody (33.2%) that may be due to vaccination against HBV in the younger age group, which has been initiated only recently in these states.
Prevalence of HBeAg was highest among cases from Arunachal Pradesh (37%) compared to Assam (16.3%) and Tripura (21.6%) (p = 0.001). HBeAg had a higher prevalence in the younger age group (43.5% in under 15 years) compared to the older age group (19.5% and 22.9% in 16 to 45 yrs and above 45 years age groups, respectively), which was statistically significant (p = 0.02). HBeAg positivity had a direct correlation with high HBV DNA viral load (p = 0.002).
A total of 223 samples were processed for relative gene expression among which 108 were cases (CHB) and 115 were controls. However, during analysis samples were excluded that did not pass the initial analysis i.e., PPC not within 20 cycles ±1 cycle; cycle difference between PPC and RTC more than 5 cycles, genomic DNA contamination i.e., any samples with HGDC below 35 cycles were excluded from analysis. Therefore, final analysis was performed on 148 samples as 75 samples were excluded from the study that did not meet the above criteria. Among the 148 subjects, 65 were from the cases and 83 from control group.
Relative gene expressions of pattern recognition receptor adaptors (MYD88 and TICAM1), pro-inflammatory cytokines, and Toll-like receptors in CHB compared to spontaneously cleared group (Controls)
It was observed that both PPR adaptors MYD88 and TICAM1 had moderate upregulation (2.2-fold & 2.3-fold) in wild-type CHB with HBeAg positivity compared to HBV spontaneously cleared controls (HBsAg –ve but HBcAb +ve) but was statistically not significant (p = 0.12). However, in CHB with HBeAg –ve group MYD88 and TICAM1 had shown only mild upregulation (∼1.5-fold) (Fig. 2A).
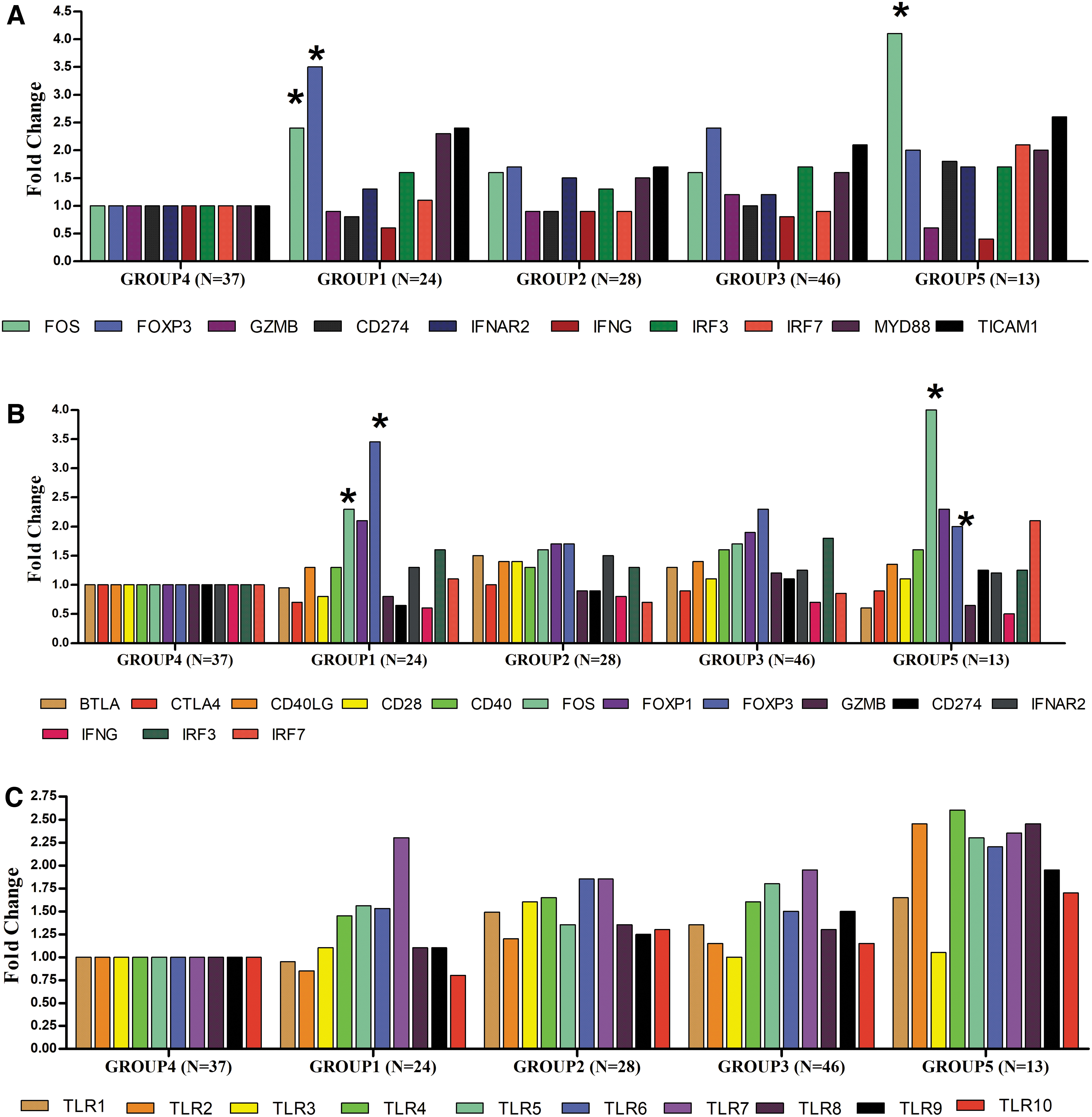
Relative gene expressions among study groups.
Relative gene expression of Tregs markers FOXP3, CD28, and CD40
Differential expression of Regulatory T cells (Tregs) markers and other innate immune genes was observed among wild CHB with HBeAg positivity compared to CHB with negative HBeAg and spontaneously cleared patients. FOXP3 was significantly upregulated in CHB with HBeAg positive status (3.1-fold upregulated, p = 0.01) compared to the spontaneously cleared group. FOXP3 was also found to be upregulated in HCC patients (Fig. 2A, B). There were no significant differences in the levels of the co-stimulatory molecules. However, CD28 expression was downregulated in CHB with HBeAg positivity compared to CHB with HBeAg negative and spontaneously cleared patients (Fig. 2B) (p = 0.1).
Relative gene expression of FOS gene and other co-stimulatory molecules
An interesting finding was that the proto-oncogene FOS gene has been found to be upregulated moderately in CHB with HBeAg +ve (2.3-fold) while in hepatocellular carcinoma cases it was found to be significantly upregulated (4.1-fold upregulation, p = 0.01) (Fig. 2A, B).
Also, CD274 (Programmed Cell Death 1 Ligand 1) was found to be moderately upregulated (1.8-fold) in HCC cases with HBV (p = 0.21). It is known that the interaction of this ligand with its receptor leads to inhibition of T cell activation and cytokine production. As expected IFN-G was found to be downregulated in chronic HBV infection (0.6-fold, p = 0.1).) and also in HCC with HBV (0.5-fold) (p = 0.18).
Relative gene expressions of TLRs in CHB compared to spontaneously cleared group (Controls)
Relative gene expression of Toll-like receptors 1 to 10 (TLR 1 to 10) in PBMC were evaluated among cases and controls (Fig. 2C). It was observed that relative gene expression in PBMC of TLRs was not significantly altered in cases and control (p = 0.14). In CHB with HBeAg positive subjects, TLR7 was found to be upregulated (2.3-fold) but alteration of expression among study groups was not significant (p = 0.08). In the HCC group with HBV, TLR2, TLR4, TLR7, and TLR8 were found to be upregulated above 2.5-fold (Fig. 2C).
Relative gene expression of innate immune genes in CHB with HBeAg(+) compared to spontaneously cleared infection
The genes FOS, FOXP3, MYD88, and TICAM1 were upregulated 2-fold or more, while IFNG was found to be downregulated −1.9-fold (p = 0.03). The most significantly upregulated gene in CHB with HBeAg positive status was FOXP3 (3.1-fold upregulated, p = 0.01) followed by MYD88 (2.2-fold, p = 0.02) (Fig. 3A).

Relative gene expression of TLR genes involved in innate immune response was analyzed between CHB with HBeAg positive status and spontaneously cleared HBV subjects (controls) in PBMC taking B2M as the endogenous control. Among all the Toll-like receptors, TLR2, TLR8, TLR9, and TLR10 were downregulated in CHB with HBeAg to healthy controls and spontaneously cleared group. TLR7 was found to be significantly upregulated (2.8-fold, p = 0.03) in CHB with HBeAg. Also, TLR7 was found to be −1.5-fold downregulated in CHB with HBeAg negative status compared to HBeAg positive status (Fig. 3B).
Discussion
HBV infection progresses to chronic infection or persistence in 5% to 10% of subjects infected with HBV (4). The main reasons for the persistence of HBV infection in adulthood are elusive and multifactorial. In this study, efforts were made to elucidate the contribution of different innate immune genes contributing to the persistence of HBV infection in adults. Patients with CHB exhibit suppression of virus-specific immune responses that is closely associated with viral persistence (25).
Relative gene expression study of 26 innate immune genes revealed that there was an increased expression in wild-type CHB with HBeAg positivity compared to spontaneously cleared or healthy controls. In our study, we have seen that in wild-type CHB with HBeAg positivity individuals are persistently infected with very high viral loads (>20,000 IU/mL). HBeAg positivity has a direct correlation with high HBV DNA viral load (p = 0.002). This is in sync with other studies that have reported that CHB is often characterized by a prolonged immunotolerant, hepatitis B e antigen (HBeAg)-positive phase, where individuals are persistently infected with very high viral loads (6,10).
HBeAg is a nonparticulate version of the HBV nucleocapsid protein and is transcribed, translated, and secreted very early in the HBV replication cycle (10). Although HBeAg is not required for viral replication or infection, it is probably responsible for the immunomodulation of host immune responses during CHB infection (6). There is increasing recognition that the course of HBV infection is influenced by the degree of innate immune response (19).
Tregs constitute a specialized subpopulation of T cells that act to suppress activation of the immune system and thereby maintain immune system homeostasis and tolerance to self-antigens (31). Tregs are characterized by several markers including CD4, CD25, and FOXP3. The constitutive expression of FOXP3 is necessary for the suppressive function of regulatory T cells (Tregs) (24). In our study, FOXP3 was significantly upregulated in CHB with HBeAg positive status (3.1-fold upregulated, p = 0.01) compared to the spontaneously cleared group. FOXP3 was also found to be upregulated in HCC patients. This supports other data that show that FOXP3 regulates tumor progression by expressing not only in Tregs but also in tumor cells of HCC (24,32).
Comparison of FOXP3 expression between HBV positive cirrhosis and HBV positive HCC by Shahera et. al. suggested that upregulation of FOXP3 was more in HBV positive HCC patients, thus supporting that FOXP3 is associated with the incidence and extent of liver damage in HBV patients (22). Further, the upregulation of FOS gene in CHB with HBeAg positivity compared to spontaneously cleared patients was also statistically significant.
FOS gene was significantly upregulated in HCC patients. FOS gene encodes leucine zipper proteins that can dimerize with proteins of the Jun family forming transcription factor complex AP-1. FOS proteins have been implicated as regulators of proliferation, transformation, and differentiation (8). Our findings suggest that FOS gene overexpression is closely correlated with tumor progression in HCC, which is in sync with other reports (33). Therefore, therapeutic targeting of FOS gene expression through selective inhibition in CHB patients may prevent tumor progression and thereby prevent HCC.
Upregulation (2.2-fold and 2.3-fold) of PPR adaptors MYD88 and TICAM1 in wild-type CHB with HBeAg positivity compared to HBV spontaneously cleared controls (HBsAg –ve but HBcAb +ve) was observed. Previous studies have found that overexpression of MyD88 inhibited HBV replication (26). As MyD88 adapter-like (Mal) acts as a bridging adapter for TLR2 and TLR4 recruitment of MyD88 to the signaling complex, our findings are consistent with a role for MyD88 in suppressing HBV replication. Thus, the overexpression of MyD88 circumvented the requirement of Mal for MyD88 recruitment (26). HBeAg can disrupt the Mal: MyD88 signaling complex, which is consistent with an inhibitory role for HBeAg in disrupting TLR: MyD88 signaling (30).
Several studies suggest that the expression and function of TLRs are impaired during chronic HBV infection. In our study, we have observed that TLR2, TLR8, TLR9, and TLR10 were downregulated in CHB with HBeAg to healthy controls and spontaneously cleared group. The reduction of TLR2 expression was reported in hepatocytes among liver biopsies and PBMCs isolated from patients with CHB (14). Interestingly, HBeAg was found to contribute to reducing TLR2 expression and impaired induction of TNF-α in hepatic cell lines. After being tested with the TLR2 ligand, another study confirmed that the expression of TLR2 on PBMCs of CHB patients was reduced with impaired cytokine production. The reduced expression of TLR9 was also found in plasmacytoid DCs (pDCs) of CHB patients, which correlate with IFN-α secretion impairment after ex vivo stimulation with TLR9 ligands (27,29).
The decreased expression of TLR3 on PBMCs, hepatic KCs, and hepatocytes were also found in CHB patients when compared with healthy controls (14).
Among all the Toll-like receptors, TLR7 was found to be significantly upregulated (2.8-fold, p = 0.03) in CHB with HBeAg positivity. TLR7 signaling pathway plays key roles against HBV, and accordingly, HBV uses some mechanisms to inhibit the pathway. The upregulation of TLR7 in HBeAg cases is difficult to explain. However, it has been reported by others that TLR7 upregulation plays a role in the serological clearance of HBV infection. Also, TLR7 was found to be −1.5-fold downregulated in CHB with HBeAg negative status compared to HBeAg positive status. Studies have suggested that by using TLR7 agonists (Imidazoquinolines, Guanosine Analogues, and GS-9620) TLR7 pathway can be amplified and HBV clearance can be induced (9). Further studies on the association of TLR7 and HBeAg positivity and viral persistence need to be explored.
It has recently been proposed that HBV establishes chronic infection via evasion of the innate immune response, which may subsequently limit maturation of the adaptive immune system for the efficient clearance of HBV (18). A study conducted by Milich and Liang and Milich et al. using mouse models illustrated that upon initial exposure to HBV, HBeAg may act through depletion or anergy of HBeAg and HBcAg-specific TH1 cells, tolerizing them as a means of establishing chronic infection (15,16). As HBeAg is implicated in the subversion of innate immune responses, our findings may contribute to the identification of therapeutic drug targets to modify the host's immune response to counteract the immunomodulatory properties of HBeAg observed in chronic HBV infection (11).
Conclusion
In this study, CHB with HBeAg positivity was found to have disrupted immune response. Thus, early induction of HBeAg seroconversion with interferon-based therapy or oral nucleos(t)ide analogs along with FOS and FOXP3 inhibitors may have an important clinical implication in the management of CHB and preventing cirrhosis and HCC. HBeAg seroconversion is an important phase in immune clearance and treatment of CHB (7,12). Therefore, personalized therapeutic regimes can be designed targeting HBeAg seroconversion based on early response and individual response.
Footnotes
Acknowledgments
The authors sincerely acknowledge all the study participants for consenting to the study. The authors also thank the Director, ICMR-Regional Medical Research Centre, NE Region, Assam, India.
Authors' Contributions
B.B. was responsible for conceptualizing and design of the study, data generation, analysis, and article preparation. M.D.S. was involved in analysis and article preparation. T.M., P.J.B., and D.B. was responsible for clinical phenotyping and diagnosis of cases. C.K.B. was involved in data generation and article preparation. H.K. was involved in study design, overall coordination of the study. All the authors were actively involved in article preparation.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The current study was funded by ICMR under the project titled “Study of Host Innate Immunity and Hepatitis B persistence.” Grant No. NER/21/2012-ECD-1.