
Abstract
Interleukin (IL)-24 is a multifunction cytokine in infectious diseases and cancers. IL-17-secreting CD4+ T (Th17) and CD8+ T (Tc17) cells promotes pathogenesis of hepatitis B virus (HBV)-associated liver disorders. However, the regulatory role of IL-24 to Th17/Tc17 cell response was not fully elucidated during HBV infection and HBV-hepatocellular carcinoma (HCC). In this study, plasma and peripheral blood mononuclear cells were isolated from 27 chronic hepatitis B (CHB) patients, 42 HBV-HCC patients and 17 normal controls (NC). Liver-infiltrating lymphocytes (LILs) were prepared from tumor and para-tumor tissues of 17 HBV-HCC patients, whereas CD4+ T cells in LILs were purified. LILs were stimulated with recombinant human IL-24. CD3+CD4+IL-17+ Th17 cells and CD3+CD8+IL-17+ Tc17 cells were investigated by flow cytometry. IL-24 level was measured by enzyme-linked immunosorbent assay. Purified CD4+ T cells were polarized for Th17 cells, and the regulatory role of IL-24 to liver-infiltrating Th17 polarization was assessed. There were no significant differences of peripheral Th17 or Tc17 cell percentage among NC, CHB, and HBV-HCC patients. Liver-infiltrating Th17 and Tc17 cell proportion was reduced in tumor tissues compared with para-tumor tissues. In contrast, plasma IL-24 level was increased in CHB and HBV-HCC patients. Exogenous IL-24 stimulation (10 or 100 ng/mL) in vitro downregulated of Th17 frequency and IL-17 secretion in LILs from both para-tumor and tumor tissues without affecting cellular proliferation or Tc17 percentage. Only 100 ng/mL of IL-24 inhibited tumor-infiltrating Th17 polarization, and this process was accompanied by suppression of nuclear factor-κB (NF-κB) and p38 mitogen-activated protein kinase phosphorylation. In conclusion, IL-24 might dampen Th17 cell response through NF-κB pathway in HBV-HCC tumor microenvironment. Elevated IL-24 might enhance anti-tumor immune response in HBV-HCC patients.
Introduction
Hepatocellular carcinoma (HCC) is the six most commonly diagnosed malignancies and the third leading cause of cancer-related death worldwide in 2020 (37). Surgical resection, liver transplantation, and administration of immune checkpoint inhibitors have strongly improved the long-term survival for HCC patients (30). However, majority of HCC patients are already in an advanced stage when clinically diagnosed, which limits the outcome of anti-tumor strategies (7). In China, chronic hepatitis B (CHB) virus (HBV) infection is still the most critical risk factor for HCC (44). Of importance, both chronic HBV infection and HCC patients present immunotolerance, leading to the dysregulation of homeostasis in both circulation and tumor microenvironment (4). However, the pathogenesis and immunoregulation of HBV-associated HCC remains not fully elucidated.
Interleukin-24 (IL-24), which previously known as melanoma differentiation associated gene-7 (MDA-7), belongs to IL-10 cytokine family (41) and plays as a multifunction protein in various diseases, including malignancies, autoimmune disorders, and infections (29). On the one hand, IL-24 has several beneficial effects in variety of human diseases. IL-24 functions as a tumor suppressor cytokine, which induces cancer-specific apoptosis without affecting normal cells (10,28). Of importance, phase I clinical trial data have shown that repeated intratumoral injection of recombinant adenovirus harboring IL-24 in advanced carcinomas was not only safe, but also induce apoptosis in a large percentage of tumor volume with measurable tumoricidal effects in ∼44% of patients (8). Moreover, IL-24 provides protections against tuberculosis (43), Salmonella typhimurium infection (25), and autoimmune diseases (18).
On the other hand, IL-24 also promotes Pseudomonas aeruginosa Keratitis in mouse corneas (33), and plays a pivotal function in the pathogenesis of proinflammatory and allergic skin diseases (32). A recent study by Chong et al. revealed that IL-17A limits Th17 pathogenicity by autocrine induction of IL-24 in experimental autoimmune uveitis (EAU) (6).
Hepatic and peripheral Th17 cells contribute to the pathogenesis of advanced HBV-related diseases, such as liver cirrhosis and HCC (20,21). However, the regulatory role of IL-24 to circulating and tumor-infiltrating IL-17-secreting cells has not been reported previously. Thus, the aim of this study was to investigate the expression of IL-24 and IL-17-producing CD4+ and CD8+ T cells (Th17 and Tc17 cells) in circulation and tumor microenvironment. The modulatory function of exogenous IL-24 to Th17 and Tc17 cells was also assessed in vitro.
Materials and Methods
The study was conformed to the ethical guidelines of the principles of the Declaration of Helsinki. The protocol was approved by the Ethics Committee of Daqing People's Hospital (ECDPH-2017012). Written informed consents were obtained from all enroll subjects or their legal guardians.
Enrolled subjects
Twenty-seven CHB patients were enrolled. Inclusion criteria were as follows: (1) positive for hepatitis B surface antigen (HBsAg) and serum HBV DNA level >6 months; (2) alanine aminotransferase (ALT) level higher than twofold upper limit of normal; and (3) treatment naive for nucleos(t)ide analogues or interferon. Forty-two HBV-related HCC patients were also included. Inclusion criteria were as follows: (1) positive for HBsAg and serum HBV DNA level >6 months; (2) alpha fetoprotein >400 ng/mL; and (3) confirmed for HCC by contrast-enhanced computed tomography or magnetic resonance imaging scan. Seventeen healthy individuals, who received physical examinations in Daqing People's Hospital and negative for HBV markers, were also enrolled as normal control (NC). Exclusive criteria were as follows: (1) coinfection with other hepatovirus or human immunodeficiency virus-1 (HIV-1); (2) afflicted with autoimmune diseases or other malignancies; (3) received chemotherapy, radiotherapy, tyrosine kinase inhibitor therapy, or immunotherapy before baseline sampling; and (4) pregnancy. There were no significant differences of either average age or sex ratio among three groups. Baseline characteristics of studied subjects are given in Table 1.
Baseline Characteristics of Studied Subjects
AFP, alpha fetoprotein; ALT, alanine aminotransferase; Anti-HBc, anti-hepatitis B core antigen; BCLC, Barcelona Clinic Liver Cancer; CHB, chronic hepatitis B; HBeAg, hepatitis B e antigen; HBsAg, hepatitis B surface antigen; HBV, hepatitis B virus; HCC, hepatocellular carcinoma; NC, normal control.
Plasma and peripheral blood mononuclear cells preparation
Anticoagulant peripheral bloods were collected from all studied subjects. Samples were centrifugated at 1,000 g for 10 min at 4°C. Plasma samples were harvested and stored at −80°C. Peripheral blood mononuclear cells (PBMCs) were prepared by using Ficoll–Hypaque (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation and stored at liquid nitrogen until use.
Liver-infiltrating lymphocytes preparation and stimulation
Tumor and para-tumor tissue specimens were harvested from 17 HBV-HCC patients (12 in stage A and 5 in stage B), who received surgery. Tissues were digested in Hank's solution containing 0.05% collagenase type IV and 0.001% DNase I for 3 h. Tissues were then cut into small pieces (∼0.5 mm3) and cultured at 37°C for 1 h. Cells were collected and passed through 70 μm strainer. Cells were washed twice and centrifugated at 50 g for 2 min. The supernatants were harvested.
Liver-infiltrating lymphocytes (LILs) were isolated by using Ficoll–Hypaque (Sigma-Aldrich) density gradient centrifugation. About 4 × 104 of LILs were stimulated with recombinant human IL-24 protein (Catalog No. 1965-IL-025/CF; 10 or 100 ng/mL; R&D Systems, Minneapolis, MN) (50) in the presence of anti-CD3/CD28 (1 μg/mL; eBioscience, Invitrogen, ThermoFisher Scientific, San Diego, CA) for 48 h.
CD4+ T cells purification and Th17 cell polarization
CD4+ T cells were purified from LILs of eight tumor tissues using MagCellect Human CD4+ T Cell Isolation Kit (R&D Systems) with FACS Aria II Flow cytometer (BD Bioscience, San Jose, CA).
For Th17 polarization, purified CD4+ T cells were cultured with anti-CD3/CD28 (1 μg/mL; eBioscience), human interferon-γ antibody (Catalog No. MAB285-100; 10 μg/mL; R&D Systems), anti-IL-4 (Catalog No. MAB204-100; 10 μg/mL; R&D Systems), recombinant human IL-6 protein (Catalog No. 206-IL-010/CF; 10 ng/mL; R&D Systems), recombinant human IL-23 protein (Catalog No. 1290-IL-010/CF; 10 ng/mL; R&D Systems), and recombinant human transforming growth factor-β1 protein (Catalog No. 240-B-002/CF; 1 ng/mL; R&D Systems) for 7 days. Half of the culture medium was replaced with fresh cytokine-containing medium on day 4, and recombinant human IL-24 protein (10 ng/mL; R&D Systems) was also added.
Flow cytometry
Cells were stimulated with phorbol myristate acetate (50 μg/mL) an ionomycin (1 μg/mL), along with brefeldin A (10 μg/mL) for 6 h. Cells were washed twice and stained with anti-CD3-PE (Clone OKT3; Catalog No. 566683; BD Pharmingen, San Jose, CA), anti-CD4 PerCP Cy5.5 (Clone SK3; Catalog No. 566923; BD Pharmingen), and anti-CD8 APC Cy7 (Clone SK1; Catalog No. 557834; BD Pharmingen) for 30 min at 4°C in the dark.
Cells were washed twice, and treated with intracellular fixation and permeabilization buffer (eBioscience, Invitrogen, ThermoFisher Scientific, San Diego, CA) for 15 min, and were then stained with anti-IL-17A-APC (Clone eBio64DEC17; Catalog No. 17-7179-42; eBioscience), antinuclear factor-κB (NF-κB) (pS529)-Alexa Fluor 647 (Clone K10-895.12.50; Catalog No. 558422; BD Phosflow, San Jose, CA), and anti-p38 mitogen-activated protein kinase (MAPK) (pT180/pY182)-PE CF594 (Clone 36/p38; Catalog No. 563569; BD Phosflow) for 30 min at 4°C in the dark. Cells were acquired using CellQuest Pro Software (BD Bioscience) by FACS Aria II Flow Cytometer (BD Bioscience). The flow cytometry data were analyzed using FlowJo Version 10 (TreeStar, Ashland, OR).
Enzyme-linked immunosorbent assay
The cytokine levels, including IL-24, IL-17A, IL-22, and granulocyte-macrophage colony stimulating factor (GM-CSF), were measured by commercial enzyme-linked immunosorbent assay (ELISA) kits (Cusabio, Wuhan, Hubei Province, China).
Cellular proliferation assay
Cellular proliferation was measured by Cell Counting Kit-8 (CCK-8; Beyotime, Wuhan, Hubei Province, China). In brief, cells were seeded into 96-well plate with the volume of 100 μL. Ten microliters of CCK-8 solution was added to each well at the last 4-h stimulation by IL-24. The absorbance was measured at 460 nm. The tested cell number with and without IL-24 stimulated was calculated by standard curve.
Statistical analysis
Data were analyzed using SPSS 23.0 Software (SPSS, Chicago, IL). The Shapiro–Wilk test was used for normal distribution analysis. Variables following normal distributions were given as mean ± standard deviation. Student's t-test, paired t-test, one-way analysis of variance (ANOVA), and Tukey test was used for comparison. Variables, which did not follow normal distributions, were given as median (interquartile range). Kruskal–Wallis test and Dunn's multiple test were used for comparison. All tests were two-tailed. A value of p < 0.05 was considered statistically significant.
Results
Tumor-infiltrating Th17 and Tc17 cells was downregulated in HBV-HCC patients
The representative flow cytometry analyses of peripheral Th17 and Tc17 cells in NC, CHB, and HCC patients are given in Figure 1A. CD3+CD4+IL-17A+ cells were defined as Th17 cells, whereas CD3+CD8+IL-17A+ cells were defined as Tc17 cells (Fig. 1A). There were no significant differences of either peripheral Th17 cell percentage (one-way ANOVA, p = 0.630; Fig. 1B) or circulating Tc17 cell proportion (one-way ANOVA, p = 0.688; Fig. 1C) among NC, CHB, and HBV-HCC patients.
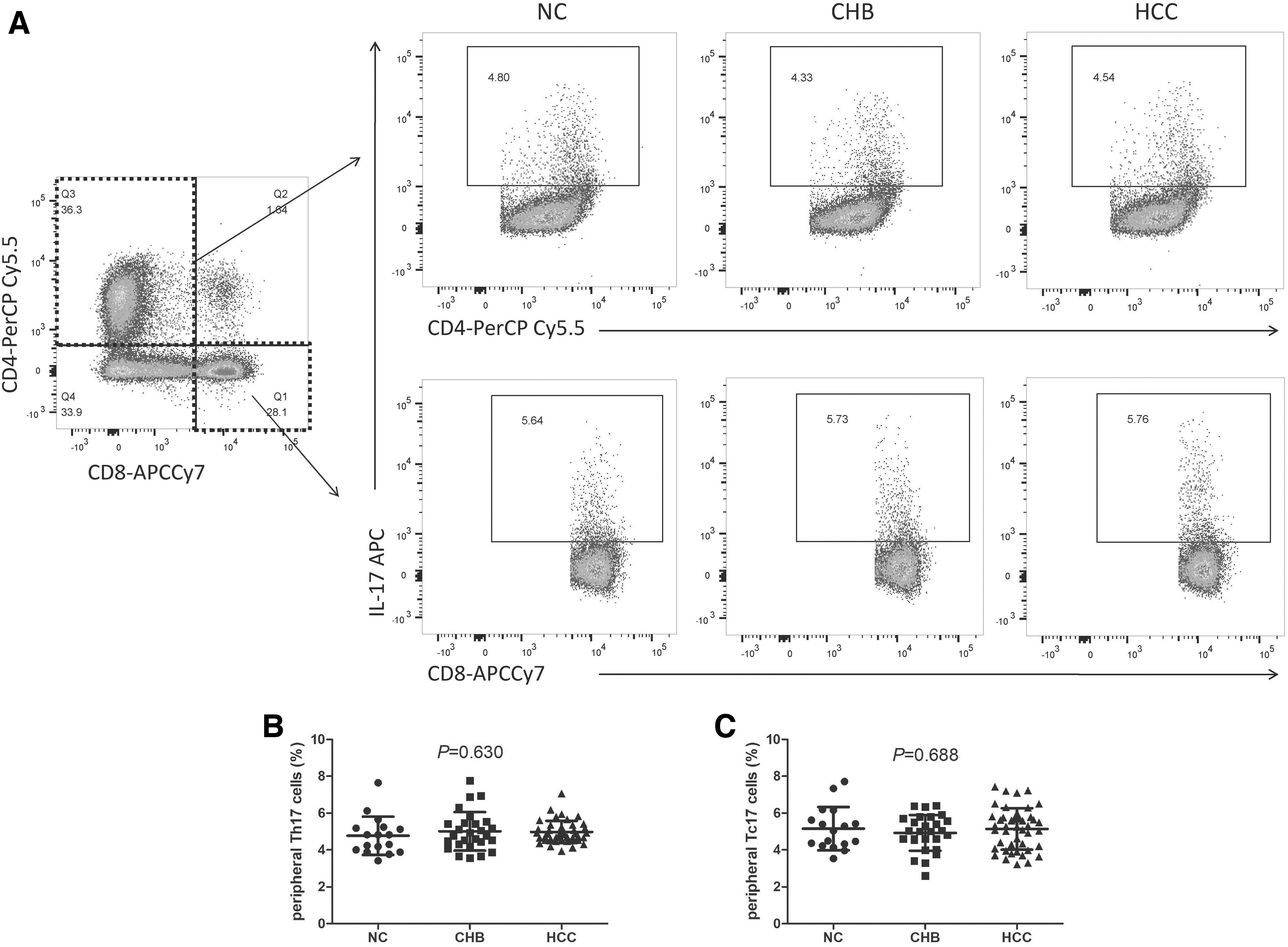
Comparison of peripheral Th17 and Tc17 cell percentage among NC, CHB patients, and HBV-HCC patients.
The representative flow dots for liver-infiltrating Th17 and Tc17 cells in para-tumor and tumor tissue from HBV-HCC patients are given in Figure 2B. The proportion of tumor-infiltrating Th17 cells was robustly downregulated in tumor tissues when compared with liver-infiltrating Th17 cells in para-tumor tissues (0.73% ± 0.12% vs. 1.03% ± 0.08%, Student's t-test, p < 0.0001; Fig. 2B). Tc17 cell frequency was also remarkably reduced in LILs from tumor tissues compared with para-tumor tissues (2.85% ± 0.87% vs. 4.32% ± 0.99%, Student's t-test, p < 0.0001; Fig. 2C).

Comparison of tissue-infiltrating Th17 and Tc17 cell percentage between para-tumor tissue and tumor tissue in HBV-HCC patients.
IL-24 was upregulated and IL-22 was downregulated in HBV-HCC patients
Plasma IL-24 was significantly elevated in both CHB patients (136.5 [75.15, 171.6] pg/mL) and HBV-HCC patients (136.3 [86.78, 219.2] pg/mL) when compared with NC (74.24 [62.47, 96.50] pg/mL) (Dunn's multiple tests, p < 0.05; Fig. 3A). There was no significant difference of IL-24 level between CHB and HBV-HCC patients (Dunn's multiple tests, p = 0.225; Fig. 3A). In contrast, IL-22 was robustly decreased in HBV-HCC patients (254.2 ± 60.76 pg/mL) when compared with NC (303.2 ± 75.28 pg/mL) (Tukey tests, p = 0.011; Fig. 3B). There was no significant difference of IL-22 level between CHB patients (285.8 ± 71.55 pg/mL) and NC (Tukey tests, p = 0.445; Fig. 3B). There was no significant difference of IL-22 level between CHB and HBV-HCC patients (Tukey test, p = 0.234; Fig. 3B).

Comparison of IL-24, IL-22, IL-17, and GM-CSF level among NC, CHB patients, and HBV-HCC patients.
Moreover, there were no remarkable differences of either IL-17 or GM-CSF level among NC, CHB patients, and HBV-HCC patients (one-way ANOVA, p > 0.05; Fig. 3C, D). There were no significant differences of either IL-24 or Th17-related cytokines among HCC patients in different Barcelona Clinic Liver Cancer stages (Kruskal–Wallis test and one-way ANOVA, p > 0.05).
Recombinant human IL-24 suppressed liver-infiltrating Th17 cells in HBV-HCC patients
About 4 × 104 of LILs from para-tumor tissues and tumor tissues were cultured with anti-CD3/CD28 in the presence of recombinant human IL-24 (10 or 100 ng/mL) for 48 h. CCK-8 results showed that IL-24 stimulation did not affect cellular proliferation of LILs from either para-tumor or tumor tissues (one-way ANOVA, p > 0.05; Fig. 4A). Th17 cell percentage in LILs from para-tumor tissues was robustly downregulated in response to 10 ng/mL of IL-24 stimulation (0.95% ± 0.14%) and 100 ng/mL of IL-24 stimulation (0.93% ± 0.12%) (Tukey tests, p < 0.05; Fig. 4B). Th17 cell proportion in tumor-infiltrating lymphocytes was only significantly reduced with 100 ng/mL of IL-24 stimulation (0.64% ± 0.09%; Tukey test, p = 0.027; Fig. 4B), but not remarkably changed with 10 ng/mL of IL-24 stimulation (0.71% ± 0.10%; Tukey test, p = 0.641; Fig. 4B). However, either 10 or 100 ng/mL of IL-24 stimulation did not affect Tc17 cell frequency in LILs from para-tumor or tumor tissues (one-way ANOVA, p > 0.05; Fig. 4C). Th17-related cytokine production in the cultured supernatants was also measured. IL-22 stimulation was comparable among no stimulation, 10 ng/mL of IL-24 and 100 ng/mL of IL-24 stimulation in LILs from para-tumor tissues (Kruskal–Wallis test, p > 0.05; Fig. 4D), whereas 100 ng/mL of IL-24 stimulation significantly reduced IL-22 secretion when compared with no stimulation in tumor-infiltrating lymphocytes (45.53 [38.22–76.39] pg/mL vs. 60.76 [49.07–130.87] pg/mL; Dunn's multiple test, p = 0.027; Fig. 4D). Moreover, IL-24 stimulation (both 10 and 100 ng/mL) notably decreased IL-17 production in LILs from both para-tumor and tumor tissues (Tukey tests, p < 0.05; Fig. 4E). However, there was no statistical difference of GM-CSF expression in the cultured supernatants among no stimulation, 10 ng/mL of IL-24 and 100 ng/mL of IL-24 stimulation in LILs (one-way ANOVA, p > 0.05; Fig. 4F).

Cellular proliferation, Th17 and Tc17 cell percentage, Th17-related cytokine production in response to IL-24 stimulation in LILs from para-tumor and tumor tissues in HBV-HCC patients. LILs were stimulated with recombinant human IL-24 (10 or 100 ng/mL) for 48 h.
Recombinant human IL-24 inhibited tumor-infiltrating Th17 polarization through NF-κB pathway in HBV-HCC patients
CD4+ T cells were purified from tumor-infiltrating lymphocytes of eight HBV-HCC patients, and polarized for Th17 cells for 7 days. Cultured supernatants were harvested at days 2, 4, and 7. IL-24 and Th17-related cytokine expression was measured. IL-24, IL-22, IL-17, and GM-CSF expression in the supernatants on days 4 and 7 was significantly increased compared with day 2 (Tukey tests, p < 0.05; Fig. 5A–D). There were no remarkable differences of IL-24, IL-22, IL-17 or GM-CSF expression between days 4 and 7 (Tukey tests, p > 0.05; Fig. 5A–D).
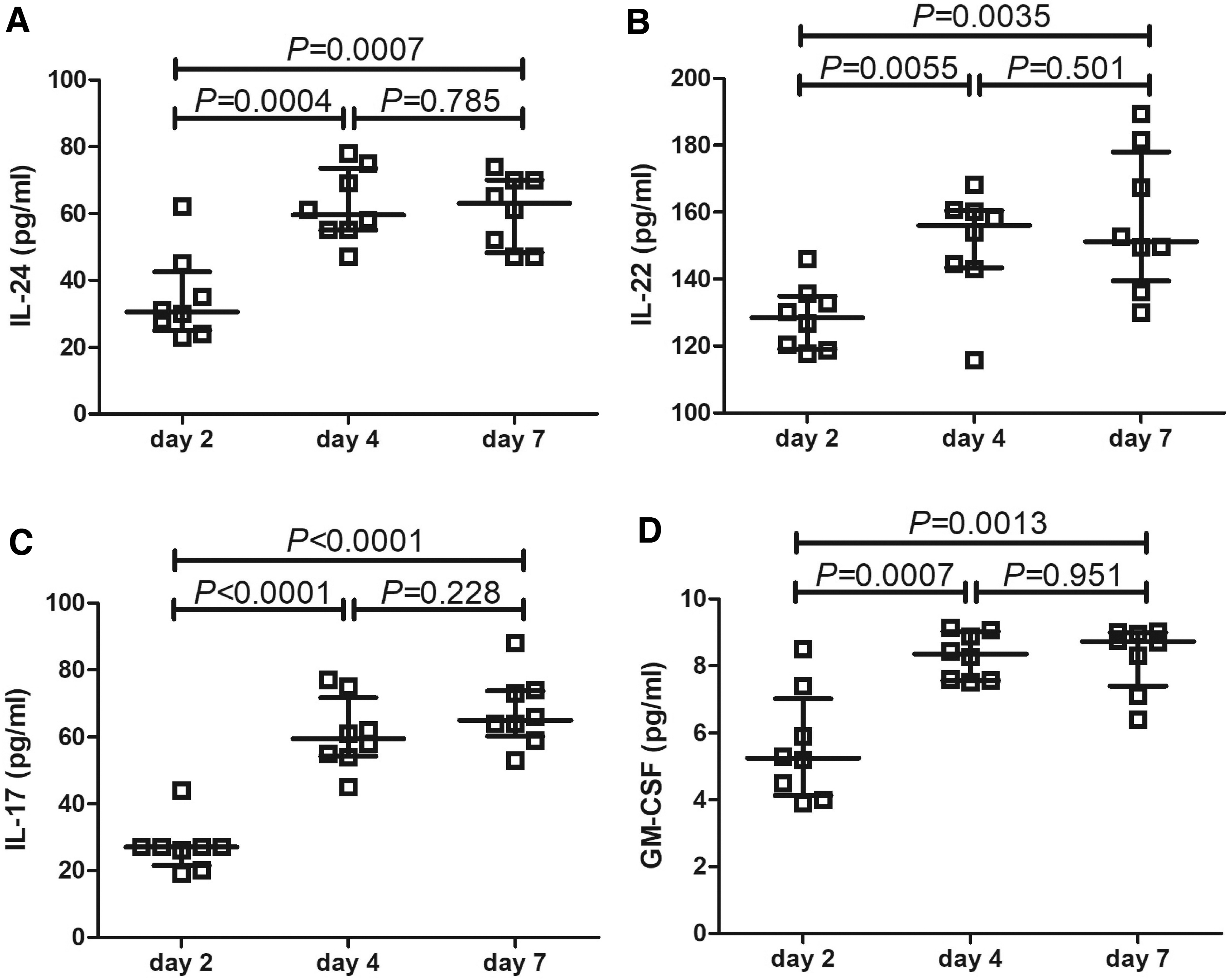
Comparison of IL-24, IL-22, IL-17, and GM-CSF expression in the cultured supernatants on different time points for Th17 polarization. Purified tumor-infiltrating CD4+ T cells were polarized for Th17 cells for 7 days. Cultured supernatants were harvested at days 2, 4, and 7.
On day 4, 100 ng/mL of recombinant human IL-24 was added to the culture medium. Cells were harvested on day 7, and were measured for phosphorylation of NF-κB and p38 MAPK as well as Th17 percentage. The representative flow cytometry analyses of mean fluorescence intensity (MFI) for phosphorylated NF-κB (p-NF-κB) and p-p38 MAPK are given in Figure 6A. Both p-NF-κB and p-p38 MAPK MFI was significantly downregulated in response to IL-24 stimulation (paired t-tests, p < 0.05; Fig. 6B, C). Th17 percentage within CD4+ T cells was also robustly reduced in response to IL-24 stimulation (18.12% ± 2.99% vs. 23.66% ± 5.08%, paired t tests, p = 0.018; Fig. 6D).
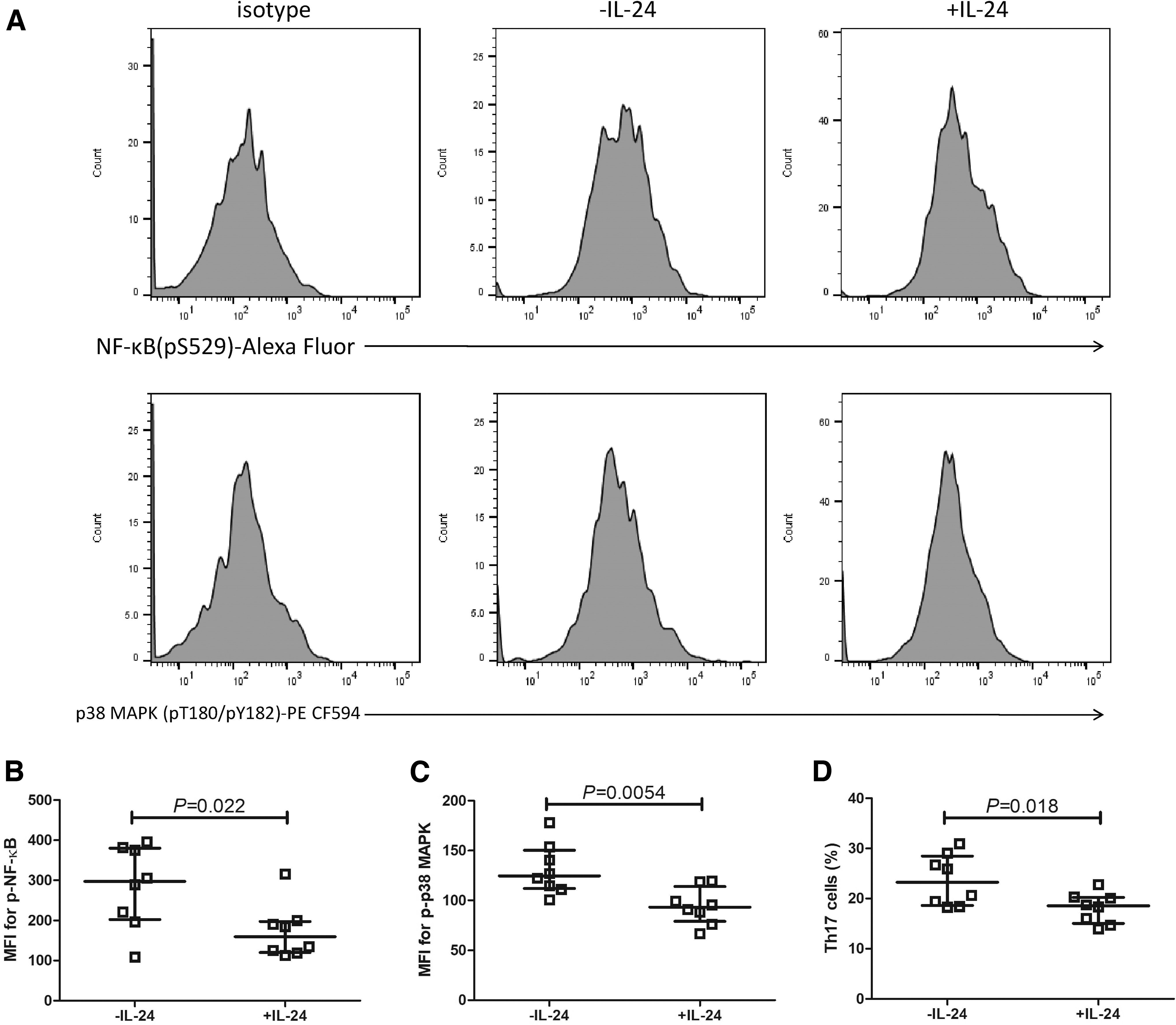
Comparison of NF-κB and p38 MAPK phosphorylation and Th17 cell percentage with and without IL-24 stimulation during Th17 polarization. Purified tumor-infiltrating CD4+ T cells were polarized for Th17 cells for 7 days. On day 4, 100 ng/mL of recombinant human IL-24 was added to the culture medium.
Discussion
IL-24 contributes to viral clearance and tumor rejection both in vitro and in vivo. IL-24 served as an antiviral factor, which inhibited influenza A virus replication through induction of Toll-like receptor 3-dependent apoptosis in vitro (34,42). HIV-1 infection also regulated mRNA and protein expression of IL-24 in PBMCs, leading to increased apoptosis in infected cells (36). Nonstructural protein 3 from hepatitis C virus induced the elevation of IL-24 mRNA expression in hepatic stellate cells (HSCs), resulting in the activation of HSCs and promotion of fibrosis (15).
IL-24 also modulated HCC cell growth and metastasis through various mechanisms, including induction of cell apoptosis and cycling arrest as well as reduction of angiogenesis (40), suppression of tumor adhesion and invasive potential (13), enhancement of cytotoxicity to cytokine-induced killer cells (46), and restoration of chemotherapeutics sensitivity (9). However, few studies focused on the IL-24 expression in HBV-associated diseases.
Our current results revealed robust elevation of plasma IL-24 in CHB and HBV-related HCC patients, indicating potential involvement of IL-24 in the pathogenesis of HBV-related liver disorders. Furthermore, the clinical significance of IL-24 expression in different malignancies was not completely understood. IL-24 level was strongly reduced in both peripheral bloods and tumor tissues in colorectal adenocarcinoma patients (50). Similarly, IL-24 expression was positively correlated with histological differentiation, and negatively correlated with the degree of lymph node involvement in rectal cancer (5).
Low-expression IL-24 in tumor tissue was a potential predictor of poor prognosis for Burkitt lymphoma (24), head and neck squamous cell carcinoma (39), and lung adenocarcinoma (14). However, we did not find statistical correlation of IL-24 expression with HCC staging and did not follow-up for the survival of HCC patients post-therapy. Thus, the clinical significance of elevation in circulating IL-24 in CHB and HBV-HCC patients remains to be investigated further.
Accumulating evidence suggested that Th17 cells and Tc17 cells played important roles in viral infection and cancers. Th17 cells and Tc17 cells contributed to either protective or pathologic immune response, which might be a context-dependent manner owing to the secretion of IL-17 and IL-22 (3,23,31,49). Decreased frequencies of Th17 and Tc17 cells were found in patients with H7N9 avian influenza A virus infection (2). Previous reports revealed that peripheral Th17 cells, but not Tc17 cells, were increased in CHB patients (19,45,49).
However, Zhang et al. showed that HBV infection promoted the recruitment of IL-17-producing T cells, leading to the elevation of Th17 and Tc17 cells in liver and peripheral blood (48). Meanwhile, increased Tc17 frequency might be a predictor for short-term morality in patients with HBV-associated acute-on-chronic liver failure (47). However, few studies focused on the Th17 and Tc17 profile in patients with HBV-related HCC.
Herein, we found that there were no significant differences of circulating Th17 cell and Tc17 cell percentages among NC, CHB patients, and HBV-HCC patients. Plasma IL-22 level was also downregulated in HBV-HCC patients. However, there were no remarkable differences of IL-17 or GM-CSF among groups. Although the current result was not in line with previous reports (11,22,49), it still indicated that peripheral IL-17-secreting T cells might not contribute to either HBV infection or HBV-induced HCC.
In contrast, liver-infiltrating Th17 cells and Tc17 cell proportion was robustly downregulated in tumor tissue compared with para-tumor tissue in HBV-HCC patients. Tumor-infiltrating Th17 and Tc17 distribution was not consistent with the findings in other malignancies, which showed increased Th17 cells and decreased Tc17 cells in thyroid tumor (16) and nonsmall cell lung cancer (35). Our current data suggested that IL-17-secreting CD4+ T cell and CD8+ T cell response might be suppressed in HCC microenvironment. The regulation of tumor-infiltrating Th17 and Tc17 response in HCC is still needed.
The functional Th17 and Tc17 phenotype could be driven by particular cytokines (27). IL-24 could mediate different immune cell types in infections and cancers. IL-24 promoted CD4+ and CD8+ T cell function, especially enhanced cytotoxicity of CD8+ T cells both in vitro and in vivo in colorectal cancer (26,50). IL-24 also promoted anti-tumor activity and persistence of chimeric antigen receptor T cell immunotherapy targeting CD19 in B cell acute lymphoblastic leukemia (12).
In contrast, recombinant IL-24 reduced Th17/Tc17 cell response in filarial infection (1) and pulmonary tuberculosis (17). Of importance, IL-17A induced autocrine IL-24 production, and in turn repressed Th17 cytokine program in EAU (6). We herein reported the modulatory activity of recombinant IL-24 to liver-infiltrating Th17 and Tc17 cells in HBV-HCC patients. It was reported that in vitro activity of IL-24 to T cells was dependent on the concentration. Only higher concentration of IL-24 contributed to T cell response in peripheral blood and tumor microenvironment in colorectal cancer (50). Thus, 10 and 100 ng/mL of recombinant IL-24 were used in our study.
Both concentrations did not affect cellular proliferation and Tc17 response to LILs. About 100 ng/mL of IL-24 suppressed Th17 cell response in both para-tumor and tumor tissues. However, 10 ng/mL of IL-24 only slightly inhibited Th17 cells in para-tumor tissues, not in tumor tissues. Meanwhile, both concentrations could dampen IL-17 secretion by LILs. This might partly be owing to the factor that IL-17 was not only secreted by Th17/Tc17 cells but also produced by other cell types, such as γδT cells and innate lymphoid cells (38).
The responsiveness of Th17 cells in para-tumor and tumor tissue might also be different. Furthermore, IL-24 secretion by tumor-infiltrating CD4+ T cells was also increased during Th17 polarization. Administration of exogenous IL-24 suppressed Th17 polarization, and this process was accompanied by downregulation of NF-κB and p38-MAPK phosphorylation in HBV-HCC tissues. This was consistent with the findings in EAU model (6), confirming the immunosuppressive activity of IL-24 to Th17 cell response.
There were several limitations in this study. First, liver specimens were only obtained from HBV-HCC patients who received surgery. We indeed tried to collect liver tissues from CHB patients who received biopsy. However, the amount of biopsy sample was insufficient for LILs isolation. Second, the current results were based on the in vitro study. Further in vivo animal experiments are still needed to confirm the regulatory role of IL-24 to Th17 cells in HBV-HCC.
Conclusion
Upregulation of peripheral IL-24 was found in patients with chronic HBV infection and HBV-associated HCC. Tumor-infiltrating, but not circulating, Th17 and Tc17 cells were downregulated in HBV-related HCC. IL-24 might dampen Th17 cell response through NF-κB pathway in HBV-HCC tumor microenvironment. Elevated IL-24 might enhance anti-tumor immune response in HBV-HCC patients. The current results indicated novel therapeutic approaches for HBV-induced HCC.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.