
Abstract
Immune system plays dual roles during human papilloma virus (HPV) infections, from defense against the virus to induction or stimulation of the HPV-related cancers. It appears that various differences within the immune-related genes and the functions of the immunological parameters of the patients are the main factors responsible for the roles played by immune system during HPV infections. Toll-like receptors (TLRs) play key roles in the recognition of viruses and activation of immune responses. The molecules also can alter the target cell intracellular signaling and may participate in the transformation of the infected cells. TLR9 is the unique intracellular member of TLRs that recognize foreign DNA, including viral DNA. Thus, TLR9 may play significant roles in the defense against HPV and its related cancers. This review article discusses TLR9 antiviral and pathological roles during HPV infection.
Introduction
Human papilloma viruses (HPVs) are small non-enveloped viruses that contain a double-stranded DNA genome (23). The virus is the cause of a wide range of diseases, such as benign warts and invasive cancers, for example cervical cancer. Accordingly, these viruses are the most predominant causes of cervical cancer, a main prevalent tumor in women (23). Additionally, investigations proved that HPVs also participate in the pathobiology of other human-related cancers, such as anal, head, and neck cancers (3,8,34). It has been reported that there are more than 200 HPV genotypes, however, the high-risk types, including HPV genotype-16 (HPV16), 18, 31, and 45, are the causes of ∼80% of cervical cancers (58).
Although the high-risk genotypes are associated with higher chance of cervical cancers, they do not induce the malignancies in some people (76). It appears that the immune system plays key roles in the determination of the outcome of HPV infections. Accordingly, the suitable innate and adaptive immunity to the HPV viruses can be associated with suppression of viral functions (24). The main differences in the innate immune responses among humans that lead to eradication or persistent HPV infections are yet to be clarified.
Toll-like receptors (TLRs) are the main clusters of the innate immune receptors, which are known as pathogen recognition receptors (PRRs) (1,49). The molecules have similar structures, but recognize different ligands and induce various innate immunity responses (1). The molecules can induce some intracellular signaling pathways that play dual roles during cancers, including fighting against the disorders and inducing the malignancies (6,50). Therefore, the roles played by TLRs during HPV-related cancers may be controversial, hence the receptors may be considered as future targets for immunotherapy against HPV-related cancers. Among the TLRs, TLR9 is the unique molecule that recognizes the pathogen double-strand DNA (63). Due to the fact that HPV contains DNA as its genome, it can be recognized by TLR9, thus the receptor might be considered as a main plausible target for interaction between innate immunity and HPV infections, and HPV-related malignancies. This review article collected recent information regarding the roles played by TLR9 against HPV viruses and also its roles in the HPV-related cancers.
Human Papilloma Viruses
As mentioned previously, HPV is a small, non-enveloped, epitheliotropic and double-stranded DNA virus. It usually infects the genital tract to induce cell proliferation, and then leads to cervical cancers (3). However, some types of HPV can induce other anogenital, oropharyngeal, and skin cancers (3).
HPV, like other papillomaviruses, has a common genetic structure, which is distinct from the polyomaviruses (26). Accordingly, HPV has a double-stranded circular DNA genome with approximately eight open-reading frames, a non-enveloped icosahedral capsid and is ∼8 kilobase pairs in size (9). HPV contains genes transcribed late (L-genes), including L1 and L2 and encodes capsid with two structural proteins, the major (L1) and minor (L2) basic proteins (9). The L2 protein plays crucial roles in either papillomavirus assembly or progression of the infection while the majority of capsids are made by L1 protein (70).
However, most genotypes encode eight major proteins, six of which are located in the “early” and two in the “late” region (9). The “early” proteins play key roles in the genome replication and transcription and also participate in the host cell cycle, signaling and apoptosis. The six proteins in the “early” region also play significant roles in the regulation of immune responses and the infected cell structural modification. Additionally, the high-risk types of HPV can produce E6 and E7 oncoproteins that play key roles against immune responses and also cell transformation (26).
High-risk genital HPVs, in particular HPVs 16 and 18, express the oncoprotein to promote the disruption of normal cell-cycle control through targeting the retinoblastoma family and p53 (5,22). The oncoproteins also induce telomerase activities that contribute to the development of malignancies (48). According to the functions, HPVs are categorized into high (16,18), intermediate (31,33,35,39,45,51,52,56,58,59,68), and low (6,11,26,40,42,43,44,53,54,55,62,66) risk viruses that participate in the induction of the cervical squamous carcinoma (21). As mentioned, HPV uses oncoproteins E6 and E7 to induce malignancies in some infected cells, however, it appears that the virus uses complicated mechanisms to induce the transforming cells. Moreover, interfering with the PRR signaling appears to be another main mechanism used by the virus.
TLR9
TLR9 is the main receptor for unmethylated CpG-DNA, the DNA for microbes (29). 3p21.3 is the location for TLR9 gene and the related-encoded protein is highly conserved. Like other TLRs, the N-terminal of TLR9 consists of leucine-rich repeats (LRRs) and it is an extracytoplasmic domain. The LRRs are followed by a hydrophobic transmembrane and a toll/interleukin-1 receptor (TIR) domain, which is in the cytoplasm. TLR9 is localized in the lysosome, endosome, and endoplasmic reticulum, as in the intracellular membrane organelles (41). TLR9 recognizes B/K-type and A/D-type, two subtypes of CpG-oligodeoxy nucleotides, as the main ligands (39). Additionally, it has been reported that the DNA of viruses contains unmethylated CpG motifs, thus, it can be recognized by TLR9 (38,77). Moreover, there are some documents that proved TLR9 can also detect the DNA without CpG motifs (35), hence, TLR9 is able to recognize DNA either with or without CpG motifs.
TLR9/ligands interactions lead to the activation of TIR domain and then, the related-adaptor protein, myeloid differentiation primary response protein 88 (MyD88) (62). Activation of MyD88 can start the related intracellular signaling that contains interleukin-1 receptor associated kinase-1 (IRAK-1), IRAK-4, tumor necrosis factor receptor associated factor 3 (TRAF3), TRAF6, transforming growth factor b-activated kinase 1 (TAK1), mitogen-activated protein kinase, activator protein 1 (AP-1), interferon regulatory factor 7 (IRF7), and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) (12,42,66). AP-1, IRF7, and NF-κB are the main transcription factors transcripted from several genes that participate in the defense against viral infections and also cell transformation to malignancies (69).
HPVs and TLRs9
It has been reported that the expression of TLRs is different in the HPV positive and negative patients and also between the patients with and without HPV-related cancers (11). Therefore, it appears that the roles played by TLR9 during HPV infections are important and need to be discussed.
TLRs9 Plays Key Roles Against HPVs
Based on the fact that TLR9 is a main member of PRRs, its roles against HPV, a DNA virus, are plausible. Accordingly, the in vitro and in vivo investigations proved the roles played by TLR9 against HPV. For example, an in vitro investigation proved the roles played by TLR9 against HPV and reported that TLR9 is highly expressed in HPV-infected HaCaT cells (44). Consequently, the study revealed that TLR9 via upregulation of type-I interferons can improve the immunity (44). Hasan et al., reported that E6 and E7 oncoproteins suppress the TLR9-dependent cytokine secretion by human keratinocytes (27). HPV-6 and 18 oncoproteins can suppress the pro-inflammatory cytokines synthesis, too (36,51). E6 and E7 oncoproteins also downregulate the expression of TLR9 in the HPV-infected uterine cervical lesions (25).
However, the study revealed that TLR9 expression gradually increased in parallel with the histopathological grade (25). Another study demonstrated that E7 from HPV-16 promotes nuclear complex formation in the human epithelial cells, which consists of estrogen receptor 1 (ESR1) and nuclear factor kappa B (NF-κB) NFKB1/RELA (p50/p65) dimers to target the TLR9 promoter (28). In parallel with the study, Pacini et al., reported that HPV38 downregulates the expression of TLR9 in the human foreskin keratinocytes (HFK) (55). Additionally, p65 interaction with histone deacetylase 1 (HDAC1) leads to histone modifications at site B (28). The phenomenon results in suppression of TLR9 transcription. A study by Ying et al. demonstrated that HPV11 transfection into HaCaT cells was associated with downregulation of TLR9 and upregulation of NF-κB p65 expression (74).
Interestingly, administration of NF-κB p65 siRNA significantly reduced expression of NF-κB p65 and rescued the expression of TLR9 in the cell lines (74). Thus, based on the in vivo studies, it appears that HPV downregulates TLR9 in the indirect format, which is illustrated in the Figure 1. Additionally, the studies in the persistent patients proved the results. For instance, Cannella et al. showed that TLR9 is expressed in the HPV-positive cervical cells (11). Upregulation of TLR9 protein during HPV infections have also been documented by several investigations (2,10). In HPV-infected women with positive antibodies to E6, higher expression of TLR9 was significantly associated with clearance of HPV after periods of persistence (60). Additionally, Tang et al., suggested that TLR9 downstream signaling pathway cannot be fully activated in Verruca vulgaris, a complication after HPV infections, leading to difficulty of HPV removal (67).

Important roles played by TLR9 against HPV. The figure illustrates that TLR9 via recognition of HPV-DNA activates MyD88 dependent pathways that results in activation of MAPK pathway and NF-κB, AP-1, and IRF7 transcription factors. The pathways lead to transcription from immune related genes against HPV. To overcome the immune responses, HPV produces E6 and E7 proteins to suppress TLR9 signaling and also downregulates TLR9 expression in ESR1 and NF-κB (p50/p65) dimers dependent manner. Variations in the TLR9 genes may also affect expression of TLR9 and defected immunity against HPV. AP-1, activator protein 1; ESR1, estrogen receptor 1; HPV, human papilloma virus; IRF7, interferon regulatory factor 7; NF-κB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; MyD88, myeloid differentiation primary response protein 88.
Downregulation of TLR9 by appropriate antagonists also increases the proliferation of HPV in the human (75). Thus, it is plausible to hypothesize that TLR9 improves immune responses against HPV via recognition of its genome. In agreement with this hypothesis, the in vitro investigations confirmed that HPV needs to suppress TLR9 expression and functions to be proliferated. Collectively, it appears that TLR9 significantly participates against HPV infections. In addition to the viral factors, the genetic of the host is another important factor for determination of HPV infection outcomes. A study by Pandey et al., demonstrated that TLR9 single nucleotide polymorphisms rs187084, rs352140, and rs352139 were associated with decreased HPV16 copy numbers (56).
Chauhan et al., showed that TLR9 rs187084 TC genotypes are higher in HPV-related cervicitis patients compared to controls (13). The significant relations between genotype TT in the −1486 locus of TLR9 with HPV infection were confirmed by Mexican researchers (46). However, the rs5743836 polymorphism within promoter region of TLR9 is not associated with HPV infections (53). Collectively, it may be hypothesized that two important factors, including HPV-related factors for inhibiting the TLR9-related immune responses and the host genetics that affect expression and functions of TLR9, determine the outcome of HPV infections by high-risk genotypes that may be eradicated or persistent. However, due to the fact that the investigations regarding the roles of the host factors are limited in the HPV infected patients, more studies are needed to determine the roles in the outcome of HPV infections by high-risk genotypes.
TLRs9 Is a Key Molecule in the HPVs-Related Cancers
Although TLR9 plays key roles against HPVs, its overexpression in the cells with persistent HPV infections was reported by Cannella et al. (11). Previous investigations proved that persistent HPV infections are the most important factors for induction of HPV-related cancers (17). Thus, overexpression of TLR9 may be in response to persistent HPV infections and considered as a key factor for induction of the related malignancies. In parallel with the hypothesis, it was documented that the expression of TLR9 in head and neck squamous cell carcinoma (HNSCC) can be associated with HPV status and high TLR9 expression can be related to poor outcomes (71). Daskalopoulos et al., demonstrated that expression of TLR9 and its related transcription factor (NF-κB) increased in oral tongue squamous cell carcinoma and its microenvironment (16).
Persistent overexpression of TLR9 during the progression of cervical intraepithelial neoplasia (CIN) has also been documented in the in vivo conditions (20,44). It appears that HPV uses TLR9 to induce or stimulate the malignancies. Consequently, some studies revealed that TLR9 is more expressed in the HPV-infected oropharyngeal squamous cell carcinoma (OPSCC) when compared to the HPV-negative OPSCC patients (33,68). Upregulation of TLR9 in the HPV-related squamous intraepithelial cervical lesion and uterine cervical neoplasm in the Mexican female population were demonstrated by Martínez-Campos et al. (46). Another study showed that TLR9 expressions were higher in the HPV16-positive cervical cancer when compared to HPV16-positive nonmalignant patients and in the latest group when compared to HPV-negative individuals (61).
The results were also confirmed by Hao et al. on the HPV-infected uterine cervical lesions (25). They showed that TLR9 expression had a positive correlation with the histopathological grade in the following order: chronic cervicitis< low-grade of CIN< medium-grade of CIN < high-grade of CIN < cervical squamous cell carcinomas. Ilmarinen et al. showed that the protein levels of TLR9 were significantly lower in the cytoplasm of laryngeal tissue cells in the laryngeal papillomas when compared to laryngeal squamous cell carcinoma (30). Therefore, overexpression of TLR9 is a main characteristic of tumors during HPV infections. Baruah et al., revealed that TLR9 helps progression of HNSCC via upregulation of PD-ligand 1 (PD-L1) and PD-L2 in the tumor tissues (7). PD-L1 and PD-L2 play crucial roles in suppressing the immune responses against tumor cells (32,37). Thus, it may be hypothesized that overexpression of TLR9 may be associated with altered intracellular signaling, and then, expression of cancer-related molecules.
Additionally, previous investigations revealed that immunotherapy against programmed cell death protein-1 (PD1) can be considered to suppress some cancers (43). A study by Cheng et al., showed that in situ immunization by TLR9 agonist significantly enhances anti-PD1 immunotherapy (15). Another investigation proved the roles played by TLR9 against HPV-related cancers and revealed that liposomal TLR9 agonist modulates functions of dendritic cells and then modulates tumor microenvironment and enhances antitumor immunity (64). In parallel with the studies, ectopic expression of TLR9 in HPV38 E6/E7 cells leads to an accumulation of the cell cycle inhibitors p21 (WAF1) and p27 (Kip1) and also declines CDK2-associated kinase activity, which results in inhibition of cellular proliferation of HFK (55). Domingos-Pereira et al., also proved the hypothesis and revealed that using TLR9 ligands was associated with regression of HPV-associated genital tumor in a mice model (18). Beneficial roles played by TLR9 ligands, as adjuvants, during immunotherapy against HPV-associated genital tumors have been documented by several investigations (14,45,52,65,72).
Therefore, based on the results it appears that only overexpression of TLR9 is not the unique factor to determine TLR9 functions. Due to the above investigations on the known TLR9 ligands, it may be hypothesized that existence of other unknown ligands in the tumor microenvironment determines the intracellular signaling of TLR9, which may be modulated by HPV oncoproteins, such as E6 and E7.
Additionally, the host genetic variations might be considered as another reason for the controversy. Associations of TC, CC genotypes of TLR9 rs187084 with the increased risk of cervical cancer in the HPV positive patients have been reported in the Indian population (57). A study on Chinese population revealed that rs352140-GA + AA genotypes within TLR9 gene were associated with the highest cervical cancer risk when compared to the non-HPV infected participants with rs352140-GG genotype (31). Another study on the Chinese population confirmed the results and revealed that rs352140-GA genotype was significantly distributed in the patients suffering from cervical cancer compared to the controls (40). Based on the limited investigations on the roles played by host genetic variations, we are unable to have a confirmed conclusion, however, due to the results on the Chinese population, TLR9 rs352140-GA genotype may be considered as a risk factor, which needs more evaluations. Figure 2 illustrates the main mechanisms played by TLR9 during HPV-related cancers.
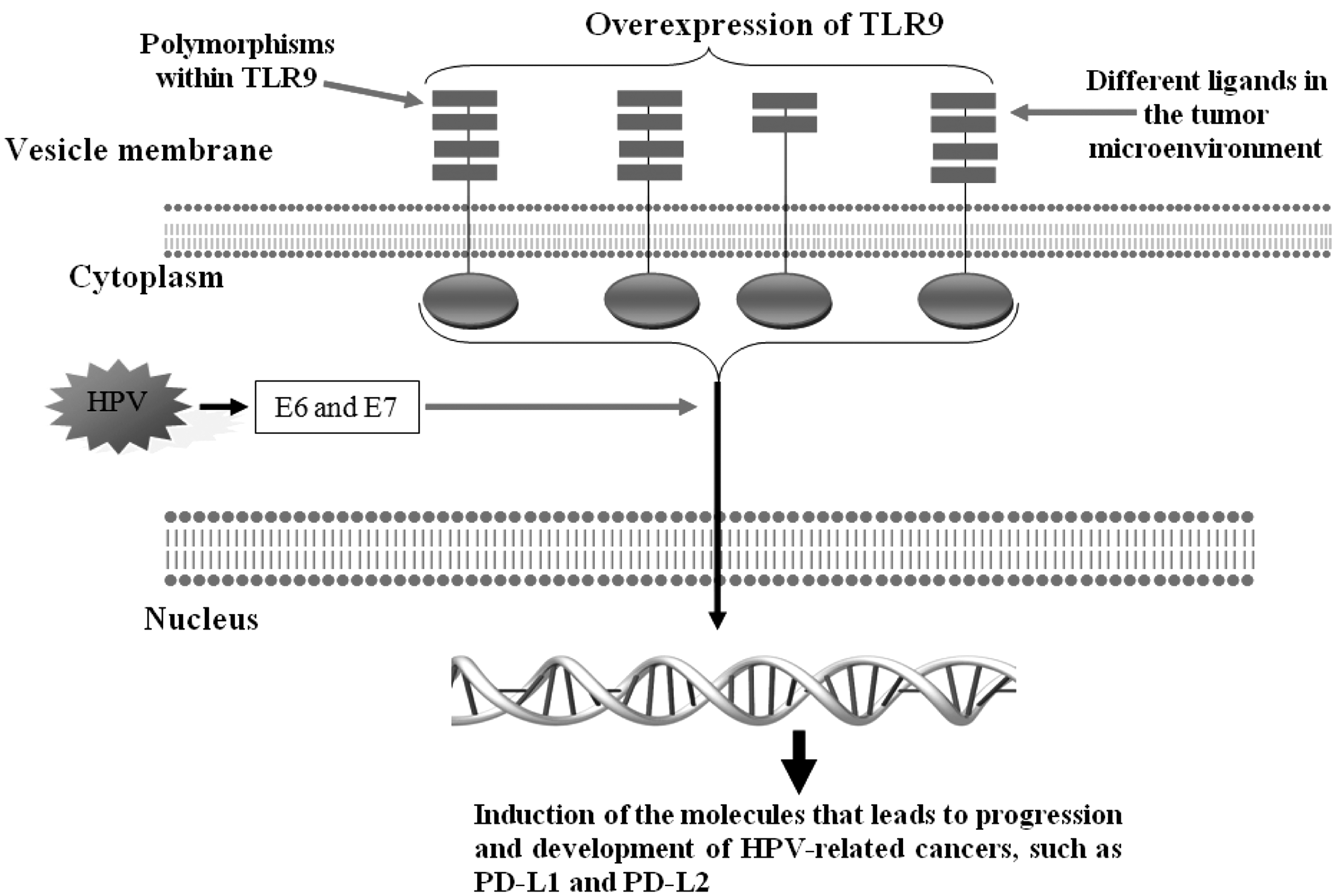
The roles played by TLR9 during HPV-related cancers. The figure illustrates that overexpression of TLR9 and the altered ligands in the tumor microenvironments may be the most important factors to alter intracellular signaling pathways that results in transcription from the molecules that participate in the development of HPV-related cancers. HPV produced E6 and E7 oncoproteins may be another factor to alter intracellular signaling pathways. Variations in the TLR9 genes may be another factor affecting expression and functions of TLR9.
Immunotherapy
Due to the important data presented in previous sections, it seems that TLR9 can be considered as a crucial target for immunotherapy. For example, using the well-known ligands may be considered to alter the functions of TLR9 in the microenvironment of HPV-related tumors. Accordingly, an investigation in murine models revealed that vaccination with synthetic long peptide in association with CpG induce robust HPV E7-specific cytotoxic T cell responses and HPV-related tumor eradication (47). Similar investigation by Domingos-Pereira et al., demonstrated that vaccination with HPV E7 protein in association with CpG induces efficient regression of genital HPV16 tumors (19). The potential roles played by TLR9 in the vaccination strategies have also been documented by Allen et al. (4). Additionally, using intravaginal TLR9 agonists, as adjuvant, increased local immune responses against HPV and its associated genital tumors (18). Thus, it appears that using the potential ligands for TLR9 can be considered to alter the functions of TLR9 in the tumor microenvironment. Another investigation showed that using combination immunotherapy can enhance the anti-HPV TLR9 ligand therapy. Accordingly, using TLR agonists and PD-1 blockade simultaneously can activate tumor-associated macrophages and enhance tumor-specific adaptive immune responses against HPV-related HNSCC (59).
In addition to appropriate ligands, other stimulators such as photodynamic therapy may be considered to alter the functions of TLR9 against HPV. In agreement with the hypothesis, Xie et al., reported that photodynamic therapy led to activation of local immunity via upregulation of TLR9 in the HPV positive genital warts (73). However, this is not simple, and the viral factors may interfere with the interventions. For example, a study by Pacini et al., revealed that UV therapy leads to upregulation of TLR9 and improvement of immune responses (54). They revealed that E6 and E7 oncoproteins from beta HPV38 significantly suppress UV-activated TLR9 expression. The investigators demonstrated that the E6 and E7 oncoproteins inhibit TLR9 expression via preventing the p53 and c-Jun recruitment to the TLR9 promoter (54). Thus, future studies need to be directed to suppress the effects of E6 and E7 oncoproteins in association with UV therapy.
Conclusion
Due to the previous investigations, it may be concluded that TLR9 is an important molecule that participates in the recognition of HPV, hence the virus uses some mechanisms to escape from TLR9-related immunity. TLR functions may be associated with pathological roles and may induce HPV-related cancers in some situations. For example, overexpression of TLR9, other unknown ligands in the tumor microenvironment interaction with TLR9, and host genetic variations may be associated with altered intracellular signaling and then, expression of cancer-related molecules. More in vivo investigations could clear the main mechanisms directing TLR9 to defend against HPV or induction of HPV-related cancers.
Footnotes
Authors' Contributions
B.D.-M. and A.K. are from Islamic Azad University, Kerman Branch (Kerman, Iran); M.K.A. is from Rafsanjan Univeristy of Medical Sciences (Rafsanjan, Iran); and B.K. is from Islamic Azad University, Baft Branch (Baft, Iran), all where education and research are the primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Islamic Azad University of Kerman, Iran (Grant No: 3165.99) and Rafsanjan University of Medical Sciences granted this project.