
Abstract
Respiratory RNA viruses are a major cause of acute lower respiratory tract infections and contribute substantially to hospitalization among infants, elderly, and immunocompromised. Complete viral clearance from acute infections is not always achieved, leading to persistence. Certain chronic respiratory diseases like asthma and chronic obstructive pulmonary disease have been associated with persistent infection by human respiratory syncytial virus and human rhinovirus, but it is still not clear whether RNA viruses really establish long-term infections as it has been recognized for DNA viruses as human bocavirus and adenoviruses. Herein, we summarize evidence of RNA virus persistence in the human respiratory tract, as well as in some animal models, to highlight how long-term infections might be related to development and/or maintenance of chronic respiratory symptoms.
Introduction
Respiratory diseases are a relevant public health problem by their substantial contribution to morbidity and mortality mainly in children and elderly people worldwide. World Health Organization reported that lower respiratory tract infections produced 2.6 millions of deaths in 2019 and were ranked as the fourth leading cause of death before COVID-19 pandemic (WHO, 2020). Community-acquired pneumonia has mostly viral or bacterial etiology; nevertheless, 45 − 75% of children and 15 − 54% of adults requiring hospitalization are affected by respiratory viruses (Jain et al, 2015; Pavia, 2013). Bronchiolitis is another lower tract inflammatory disease that affects about one in nine children during their first year of life, although incidence is higher in the 3- to 6-month age group, and viruses are also the most common cause (Petat et al, 2021; Rivera-Sepulveda and Garcia-Rivera, 2017).
Viruses associated with human respiratory tract infections are human respiratory syncytial virus (HRSV), human rhinovirus (HRV), influenza A and B, human metapneumovirus (HMPV), parainfluenza virus (PIV), endemic coronaviruses (CoV-229-E, CoV-OC43, CoV-NL63, and CoV-HKU1), bocavirus (BoV), and adenovirus (ADV), of which HRSV has been identified in up to 45% and 88% of infants with moderate-to-severe bronchiolitis living in tropical and temperate countries, respectively (Miller et al, 2013; Petat et al, 2021; Rivera-Sepulveda and Garcia-Rivera, 2017). Likewise, HRV is associated with up to 34% of cases of bronchiolitis, and it is codetected in 38% of infants with HRSV (Miller et al, 2013; Petat et al, 2021).
Interestingly, it has been documented that infants hospitalized for acute viral bronchiolitis at around 2 − 6 months of age have significantly increased risk to develop recurrent asthma-like symptoms during their first decade of life (Miller et al, 2013; Pérez-Yarza et al, 2007; Petat et al, 2021; Wu et al, 2008).
Studies to evaluate the relationship between viral bronchiolitis and development of asthma-like illness have not concluded whether it is causal (e.g., by epithelial damage, long-term viral infection, chronic inflammation, and altered activation of airway nerves) (Fig. 1), or if bronchiolitis is a triggering factor that unveils a genetic predisposition for atopy, although they might be nonmutually exclusive events (Pérez-Yarza et al, 2007; Petat et al, 2021).

Chronic respiratory diseases like wheezing and asthma have been associated with severe acute viral infections early in life, which alter the epithelial barrier triggering exposure and remodeling of airway nerves, leading to increased risk of bronchoconstriction. Aberrant chronic inflammation induced by viral exposure early in life and/or viral persistence might also contribute to inflammation and airway hyperresponsiveness.
DNA viruses such as ADV and BoV normally establish asymptomatic persistence in the human respiratory tract (Garnett et al, 2009; Xu et al, 2021), but it is still not clear whether respiratory RNA viruses establish long-term infections in subjects without underlying medical conditions or their possible role in chronic airway hyperreactivity (AHR). Current COVID-19 pandemic has contributed with many clinical and fundamental studies describing long-term coronavirus shedding (Choudhary et al, 2022; Goubet et al, 2021; Tarhini et al, 2021). Accordingly, in this review we summarize evidence of RNA virus persistence in the human respiratory tract (excluding coronaviruses), as well as in some animal models, to underline how long-term infections might elicit development of chronic respiratory symptoms.
Persistent Viral Infections
A viral infection is a dynamic event that progresses in acute manner when the virus replicates actively eliciting its own transmission to new hosts, but then it is cleared by activity of both innate and adaptive immune responses. Alternatively, other viruses establish persistent infections (normally after an acute process) through effective mechanisms to evade the host immune response and by molecular strategies that modulate their replication to avoid significant tissue damage. In such a way, it has established a metastable virus−host equilibrium, with an immune system adjusted to the continuous presence of viral antigens (e.g., nucleic acids), still contributing to limit viral replication, but modulated to circumvent local and systemic damage (Kane and Golovkina, 2010; Virgin et al, 2009). This is mainly true for human DNA viruses like those from the Herpesviridae and Adenoviridae families, since infection by viruses like hepatitis B and hepatitis C is in fluctuating dynamics with destruction waves of infected cells mediated at least by cytolytic lymphocytes that entail progressive hepatic damage (Iannacone and Guidotti, 2022; Rios et al, 2021).
Persistent viral infections may be classified as chronic and latent. In the former, virus assembly or expression of viral antigens occurs constantly. In contrast, during latency the viral genome is dormant without expression of viral genes or with expression of few essential genes for assurance of intracellular permanence (Fig. 2), although reactivation may occur transitory under some physiological circumstances to allow new virion assembly and spread (e.g., immunosuppression, hormonal imbalance, pregnancy, chemotherapy, and aging) (Stoeger and Adler, 2019). Retrovirus and many DNA–virus genomes encode molecular mechanisms or latency programs for successful persistence evading apoptosis and the host immune response, whereas replication of the viral genome and its segregation is secured to warrant long-term infection (Cohen, 2020; Ta et al, 2022; Weidner-Glunde and Kruminis-Kaszkiel, 2020).

Viral persistence may be chronic or latent. Chronic viral infections may progress as productive when assembly of viral particles is constant, or as nonproductive when infectious virus is not detected, despite the genome remaining intracellular and it is possible to detect expression of some proteins. During latency the viral genome is not able to produce a viral progeny since it is dormant, although few essential genes might be expressed to secure intracellular permanence and inheritance to daughter cells if the host is not a terminally differentiated cell.
Until now, measles, hepatitis C, and some enteroviruses have been recognized as RNA viruses with capacity to infect persistently their human hosts (Chen et al, 2020; Griffin, 2020; Rios et al, 2021). However, there is growing evidence that supports this same ability that exists in other RNA viruses, such as Ebola, Zika, HRV, and Chikungunya (Aid et al, 2017; Engelmann et al, 2017; Hawman et al, 2016; Kurscheidt et al, 2019; Racine and Kobinger, 2017; Thorson et al, 2021). Understanding mechanisms of viral persistence is essential to monitor this type of infection and its possible relationship with human recurrent or chronic diseases.
Up to now, demonstration of a persistent infection in humans is not easy task since longitudinal studies are not straightforward, mainly when sampling is invasive, and subjects do not commit for long periods of time. Animal models are outstanding study tools, but they are not natural hosts for human viruses, thus extrapolating information must be careful.
In next sections, we present evidence regarding persistence of RNA virus in respiratory tissues, and its possible association with chronic disease.
Viral Persistence in Respiratory Human Tissues
Persistence in immunocompromised hosts
The potential of respiratory RNA viruses to establish long-term infections in the respiratory tract has been demonstrated in immunocompromised individuals, mainly by influenza, HRV, HRSV, PIV, HMPV, and CoV (Hijano et al, 2018; Maziarz et al, 2010). Hematological patients are particularly at higher risk to develop life-threatening lower respiratory tract infections and long viral shedding, which frequently are associated with leukopenia and hypogammaglobulinemia (Srinivasan et al, 2011; Tabatabai et al, 2018).
In a study, hematological adult patients hospitalized for stem cell transplantation were screened by reverse transcriptase-quantitative polymerase chain reaction (RT-qPCR) for influenza, PIV, and HRSV on admission. From 111 virus-infected patients, 13, 40, and 64 were positive for PIV, influenza, and HRSV, respectively (6 patients showed coinfections). Only 67% of these patients received transplant, and follow-up was performed under respiratory symptom development by consecutive nasal swabs. Long-term infection (>30 days) was identified in 20 transplanted subjects with median viral RNA detection of 80 days (range 35 − 334 days). HRSV represented 80% of chronic infections and showed the longest persistence, whereas PIV and influenza were detected up to 36 and 48 days, respectively (Lehners et al, 2016).
Another prospective study in an adult cohort that underwent stem cell transplantation (n = 21) showed HMPV infection and persistence in 16 asymptomatic/subclinical subjects during 26 to 94 days. According to virus isolation in cell culture, only two patients displayed replication competent virus in two consecutive nasopharyngeal aspirates obtained 12 and 56 days apart, suggesting that a nonproductive persistence was predominant in this group (Maurizia Debiaggi et al, 2006). Even though several long-term respiratory infections appear to progress in absence of virus assembly (nonproductive infections), the few subjects detected with replication competent virus might be contagious for a long time. In fact, prolonged viral shedding is considered a risk for nosocomial spreading and is a major challenge for infection control management in health care settings with transplant recipients and oncologic patients (Lehners et al, 2016; Weinstock et al, 2003).
Influenza A virus also establishes long-term infections (range of 14 − 275 days), mainly in patients with lymphopenia (Gooskens et al, 2009), and antiviral specific treatment (e.g., amantadine, rimantadine, and oseltamivir) is a risk factor for arising of resistant mutants, which could have IC50 values up to 7000-fold higher than the IC50 of pretreatment isolates (Boivin et al, 2002; Gooskens et al, 2009). Interestingly, influenza A virus persistence often associates with detection of replication competent virus, including those drug-resistant variants, increasing risk of transmission (Boivin et al, 2002; Weinstock et al, 2003). In this case, viral clearance has been achieved by lymphocyte reconstitution (Gooskens et al, 2009).
HRV from the genus Enterovirus is the leading cause of common cold, and normal mean time of viral shedding is for 2 weeks in general population. However, it is also associated with lower respiratory tract infections, persistence in immunocompromised and exacerbations of asthma, and chronic obstructive pulmonary disease (COPD) (Peltola et al, 2013; Piralla et al, 2012). To distinguish among virus persistence and reinfection, it has been proposed that it should exist in a very high nucleotide sequence identity between specific viral genes or full genomes isolated from consecutive samples taken at different time intervals from the same subject. Another criterion is to find the same viral genotype in subsequent samples (Ammerman et al, 2020; Dijkema et al, 2016; Engelmann et al, 2017; Gooskens et al, 2009; Tang, 2010). Such principles have been applied to evaluate persistence of viruses like HRV.
A prospective study that included mostly immunocompromised subjects disclosed a median time of HRV shedding of 92 days (range 50 − 455 days) in almost 10% of them. In this group, respiratory samples taken 45 days or longer apart were sequenced to compare the viral genome region VP4/VP2. As sequence divergence was <10% it was assumed as long-term infection by the same viral genotype (Engelmann et al, 2017). In contrast, pediatric lung transplant recipients develop respiratory virus infections principally within the first 6 months post-transplant, and HRV has been found as the leading etiology with capacity to persist also in nearly 10% of subjects, according to virus genotype determination (Ammerman et al, 2020). Long-term infection by this same virus has been described up to for 5 months in patients with hypogammaglobulinemia by sequencing of the genomic 5′ noncoding region (Kainulainen et al, 2010) and in stem cell transplant recipients in accordance with nucleotide sequence of the VP1 gene (Canducci et al, 2013); thus, HRV is one of the best identified virus with capability to persist in the respiratory tract.
Normally, immune pressure appears not to be a driving force for viral evolution in immunocompromised hosts (Tabatabai et al, 2018), but antiviral drugs (as mentioned for influenza virus) and physiological changes that affect susceptibility to infection (e.g., by changes in receptor expression) may be drivers of emergence of new variants. Greninger et al (2021), described persistence of PIV-3 over 98 and 278 days in two patients with leukemia and stem cell transplant. Viral loads evaluated by RT-qPCR displayed cycle thresholds usually under 25, indicating continuous replication of the viral RNA. Interestingly, virus genome sequencing revealed many individual mutations that appeared after day 70.
One mutation (H552Q) in the envelope glycoprotein hemagglutinin neuraminidase (HN) was of particular interest since it conferred increased avidity for the virus receptor (sialic acid) and was associated with increased fusogenic capacity, suggesting that infectious process was improved mainly by cell-to-cell spreading. However, it was hypothesized that this property might reduce human-to-human transmission. Emergence of such mutation was related to patient pharmacological treatment with DAS181, which is a recombinant neuraminidase that cleaves sialic acid from host tissues preventing viral entry (Greninger et al, 2021). The H552Q mutation in HN had been previously identified in a cell-culture adapted PIV-3, which was low infectious in a rat model and in human airway epithelial cells. Possibly H552Q mutation reduces stability of the prefusion complex formed by HN and the fusion glycoprotein (F). Sialic acid enriched tissues (such as airway epithelium) may induce premature triggering of F because of the increased avidity of HN by its receptor, thus, reducing PIV-3 infection efficiency (Palmer et al, 2012).
In addition to PIV, HRSV has shown very long periods of persistence in immunocompromised adults and children, with evidence indicating that it is not caused by reinfections. Tabatabai et al (2018) evaluated the genetic variability of the HRSV G gene from hematological patients (n = 16) persistently infected with RSV A, subgroups NA1 and ON1. Interestingly, most patients displayed viruses with identical G gene sequence over the course of infection (12/16). Nevertheless, two patients that experienced severe immune suppression and the longest persistence (142 and 333 days) harbored viruses with mutations leading to premature stop codons. These patients repeatedly received intravenous immunoglobulin, and after immune reconstitution, viruses with stop codons in G gene decreased until clearance.
Accordingly, an unsuccessful immune response promotes development of viral variants at low frequency because of the lack of selective pressure, as long as immune therapies or drugs are not administered (Tabatabai et al, 2018). Adaptations in long-term immunosuppressive therapies in transplant recipients (allowing residual immunity) to prevent or reduce severe diseases by infectious agents might play a role in intrahost viral evolution during persistent infections, as described for HRV in lung transplant recipients (Tapparel et al, 2011), making genomic surveillance an ideal tool to track emergence of new viral variants in immune suppressed groups.
Persistence in nonimmunocompromised hosts
Understanding relevance of respiratory virus persistence in immunocompromised patients has been essential for clinical management at the level of prophylaxis and treatment. However, it is still lacking information about clinical and epidemiological implications of virus persistence in the respiratory tract of nonimmunocompromised individuals since it might be a risk factor for chronic airway inflammation.
Adenotonsillar hypertrophy is a common affection in the upper respiratory tract of pediatric patients, which has diverse proposed etiologies such as allergies, gastroesophageal reflux disease, as well as bacterial and viral infections (Kim et al, 2018; Soderholm et al, 2018). Obstructive sleep apnea (OSA) has a prevalence of 1–3% in children, and adenotonsillar hypertrophy is a factor that increases its incidence (Wang et al, 2019).
In a study, respiratory viruses like HRSV, HMPV, ADV, CoV-NL63, HRV, PIV-3, and influenza A and B were evaluated in hypertrophic tonsils extracted by surgery from children with OSA (n = 34). Every patient had suffered a lower respiratory tract infection at least 12 months before surgery, and 7/34 subjects had recurrent wheezing or asthma. Analysis by RT-qPCR revealed that 17 patients were positive for respiratory viruses in the following order of frequency: HRV>ADV>HMPV>HRSV>CoV-NL63. In contrast, PIV-3, influenza A, and influenza B were not found. Interestingly, samples were collected in a northern hemisphere country in months from June to September from children without respiratory symptoms or recent acute respiratory disease. As it was not yet season for HRSV or seasonal CoV-NL63, their finding might suggest persistence at least for these two viruses (Nichols et al, 2021; Yeshuroon-Koffler et al, 2015).
Parallel analysis of adenoids, palatine tonsils, and nasopharyngeal secretions from children ≤14 years with diagnosis of tonsillar hypertrophy or recurrent tonsillitis revealed that 97.5% (118/121) had at least one of nine respiratory viruses evaluated, being ADV, Enterovirus, and HRV the most frequent (38 − 47%), followed by BoV, HMPV, and HRSV (15 − 30%). Enterovirus detection in tonsils was statistically associated with higher hypertrophy degrees, and long-term infection was considered as probable, since no respiratory symptoms were detected in virus-positive children. Thus, it was hypothesized that virus persistence contributes to chronic inflammation owing to constant stimulus of the immune system by viral antigens (Proenca-Modena et al, 2012). Interestingly, it has been observed that adenoids function more often as target tissue than tonsils for respiratory viruses and possibly also as viral reservoirs, indicating that respiratory lymphoid tissues are differentially infected; so, they should be independently studied to evaluate prevalence of infectious agents (Faden et al, 2016).
In contrast, Mikola et al (2018) comparatively evaluated transcription of inflammatory and anti-inflammatory cytokines in tonsils from patients with tonsillar hypertrophy (n = 47) and recurrent tonsillitis (n = 42), observing no significant differences between groups, excepting for interleukin-37 (IL-37) that was overexpressed in the former. In addition, respiratory virus infection was more frequent in tonsillar hypertrophy (79% vs. 38%) (Mikola et al, 2018). IL-37 is produced in pro-inflammatory environments by monocytes, macrophages, dendritic cells, and B-lymphocytes to control inflammation and ameliorate tissue damage (Su and Tao, 2021). Therefore, increased levels of IL-37 mRNA in hypertrophic tonsils might reflect a chronic inflammatory state produced by prolonged exposure to antigens; however, further studies are needed to determine utility of this cytokine as a biomarker of long-term infections in lymphoid respiratory tissues.
Up to now, only few studies have performed molecular typing in immunocompetent cohorts to discriminate between viral persistence and reinfections.
Brint et al (2017) observed in nonimmunocompromised infants (younger than 6 months) infected by HRSV, a tendency for development of severe bronchiolitis in comparison to older infants, which was associated with higher viral loads and slow virus clearance, in such a way that RT-qPCR remained positive at least for 35 days after symptom onset. Remarkably, viral load rebound was detected in 37% of infants with long-term infection in whom the same HRSV subtype was identified in consecutive samples, suggesting that such rebound was not result of reinfection (Brint et al, 2017).
Similarly, a multicenter cohort of infants under 1 year of age hospitalized with bronchiolitis showed delayed HRSV clearance in 19% of individuals (106/557) 3 weeks after admission. This prolonged infection occurred in the group of younger children (<5 months age), affected by higher viral loads. Sequencing of a hypervariable region of the G gene confirmed infection by the same HRSV subtype (and probably same virus) at beginning and end of the study. Similar findings have been described during HRV infection in children <1 year of age, with persistence and reinfections (determined by genotyping) in 4.5% and 14.5% of cases, respectively (Loeffelholz et al, 2014), indicating that younger infants are prone to undergo longer infections. Interestingly, it was found that respiratory microbiota with abundance of Haemophilus was associated with a higher risk of delayed viral clearance, suggesting that microbiota composition might alter immune response, particularly to clear HRSV (Mansbach et al, 2019).
Prolonged virus infection has also been observed in immunocompetent adults (median age 55 years), through a study designed to discriminate between HRV persistence and reinfection. In that cohort, HRV was detected in 18% of individuals (444/2485) to whom 5% showed prolonged viral shedding (31 − 40 days). According to genotyping, 65% of such prolonged detections corresponded to reinfections and 35% to persistence (nucleotide similarity of 99.9 − 100%). In this case, reinfections were associated with underlying conditions of COPD and asthma, whereas persistence occurred mainly in older patients (median age 72). Thus, long-term respiratory virus infections are more frequent at the extremes of age (Zlateva et al, 2014).
Asthma and COPD are the most common chronic respiratory diseases, and respiratory viruses like ADV, HRV, HRSV, CoV, picornaviruses, and influenza have frequently been associated with worsening of such clinical conditions. It is estimated that 262 million people worldwide suffer from asthma, whereas over 3 millions of deaths are associated with COPD yearly (WHO, 2022). There is practically consensus that viral infection generates frequent and intense exacerbations. Consequently, clinical research on this topic is often based on the hypothesis that respiratory viruses are a target for prevention and/or treatment of the acute condition (Tan et al, 2003). However, virus participation in pathogenesis of asthma and COPD is still not clear since only few prospective studies have been performed.
Marin et al (2000) found ADV, HRV, and CoV genome by nested PCR in asthmatic children (n = 50) who had been asymptomatic at least for 3 weeks before sampling; percentages of virus detection in a total of 88 nasopharyngeal swabs were 78.4%, 32.4%, and 2.7%, respectively. In the control healthy group (n = 20) only one ADV positive sample was detected. Important to consider in this study is that patients were under treatment with inhaled glucocorticoids. Accordingly, virus detection in the asymptomatic period suggested persistent infection, although it might have been driven by glucocorticoid therapy (Marin et al, 2000).
Kling et al and Wood et al (2011), detected HRV infection in more than 40% of adults and children up to 6 weeks after asthma exacerbation; however, it was determined in the last study that long-term infections were caused by different virus genotypes, by sequencing of a VP4/VP2 fragment. These results agreed with another study that described in adult patients with stable asthma, respiratory virus reinfections instead of viral persistence throughout a period of 12-week sampling (Turchiarelli et al, 2011).
In contrast, Falsey et al assessed HRSV persistence in a 1-year longitudinal study in a cohort of adults with a diagnosis of COPD (n = 92), through respiratory sampling during the asymptomatic state and in acute illness. Results showed that HRSV detection occurred normally during acute exacerbation, whereas only 0.9% of subjects were positive during stable COPD (Walsh and Falsey, 2004). In contrast, Wilkinson et al (2006) found in a 2-year longitudinal study (n = 74), 59 subjects with at least one HRSV positive sputum sample in the stable phase, whereas 32.8% of the overall samples (n = 241) collected under same clinical circumstances were virus positive.
Of relevance, patients with ≥50% of HRSV positive samples during stable COPD had significantly accelerated lung function decline and augmented levels of IL-8, IL-6, and myeloperoxidase, suggesting that this virus might participate in pathogenesis of this chronic disease by contributing to epithelium inflammation and subsequent deterioration in lung function (Wilkinson et al, 2006). Besides, an in vitro study described in both A546 epithelial cells and human tracheobronchial cells that HRSV infection augmented TLR-3 expression and its localization in plasma membrane, altering epithelial sensitivity to dsRNA, along with increased levels of IL-8 (Groskreutz et al, 2006), suggesting that this virus sensitizes airway epithelial cells to subsequent viral antigens, possibly contributing to COPD and asthma exacerbations.
Such discrepancies between studies to determine persistence and role of RNA virus in asthma and COPD could be reassessed, as mentioned before, through prospective studies implemented with genotyping and whole-genome sequencing.
Evidence of Respiratory RNA Virus Persistence in Animal Models
A variety of viral infections affecting humans have been studied in diverse animal models, although mouse has become the most useful tool, due to its ease of handling, has low maintenance requirements, but mainly because there exists a deep knowledge of its anatomy, physiology, and immunology; besides that, its complete genome has been sequenced. Thus, similarities and differences between mice and humans can be considered in both modeling of human diseases and data interpretation (Rydell-Törmänen and Johnson, 2019).
In this regard, HRSV persistence in female Balb/c mice has been studied that is associated with a pathological state. Mice intranasally infected with ∼8 log10 PFU/mL of HRSV showed at 42 days postinfection (dpi) undetectable levels of viral RNA in bronchoalveolar lavages, even though whole-lung tissues exhibited ∼6.5 log10 RNA copies/mL, suggesting a lack of virus assembly and/or release despite active viral genome replication. Immune response assessed by cytokines was predominantly Th1, with an increase of interferon (IFN)-γ and AHR that positively correlated with viral RNA copy numbers in lung tissue (p < 0.01; r = 0.8) (Estripeaut et al, 2008). In a more recent study, 2.0 log10 RNA copies/mL was recovered from mice lung tissue up to 60 days after initial infection with 5.5 log10 PFU of HRSV.
Viral load was associated with AHR increase and airway inflammation mainly mediated by lymphocytes and macrophages, although infiltrating cells gradually decreased with time. IFN-γ significantly augmented at 7 dpi, but normal levels were restored at day 60. Of relevance, levels of neurotrophins like nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) significantly increased gradually during chronic infection, from day 14 to 60 (Zang et al, 2015). Immune cells and respiratory epithelium produce such neurotrophins; both are augmented during human asthma and COPD, possibly playing a role in eosinophil activity, airway smooth muscle contractility, and lung tissue remodeling (Liu et al, 2021; Prakash and Martin, 2014). Analysis of NGF and BDNF during evident respiratory virus persistence in humans might help to understand pathophysiology of chronic respiratory diseases.
Regarding HMPV, it also establishes chronic infections in mice. Liu et al (2009) showed lung viral titers in a biphasic pattern with a first peak at 7 dpi and a second peak by day 28 postinfection that remained high up to day 42. Afterward, viral load declined but persisted at least up to day 56 (200 PFU/g lung tissue). Besides epithelial cells, PGP9.5+ neuronal cells that innervate the lungs were positive for HMPV G protein and genomic RNA along 42 days (Liu et al, 2009); however, both viral products were undetected at 77 dpi. Interestingly, treatment with dexamethasone at this last time reactivated G protein and viral RNA detection, whereas virus titers showed ∼100 and ∼600 PFU/g in lung tissue at days 7 and 13 post-treatment, respectively. Authors suggested that HMPV persisted at very low-to-undetectable levels in neuronal fibers and treatment with dexamethasone induced immunosuppression allowing virus reactivation and reinfection of respiratory epithelium (Liu et al, 2009).
In contrast, rat model is another convenient tool for studying human diseases (Szpirer, 2020). HRSV vertical transmission was observed in pregnant female rats, after intratracheal infection at day 12 of gestation. Virus was detected in 67% of rat fetuses, whereas 4/10, 2/12, and 2/8, newborns, weanlings (2 weeks of age), and adults (10 weeks of age), respectively, were also virus positive. HRSV isolation from fetal tissues was achieved by coculture with HEp-2 cells, confirming vertical transmission and persistence. Gene transcripts of NGF and its receptor TrkA were found augmented 2.2- and 6.1-fold, respectively. In addition, prenatal exposure to HRSV was associated with significantly increased postnatal respiratory smooth muscle contraction upon infection with the same virus, along with twofold increased levels of acetylcholine, in comparison to subjects nonexposed to HRSV in utero. Therefore, in the rat model, fetal exposure to HRSV promotes a persistent infection (possibly by immune tolerance) and dysregulation of neurotropic pathways leading to AHR by exposure to postnatal respiratory stimuli (Piedimonte et al, 2013).
Besides infection of experimental animals with human viruses, animal viruses can be used to understand dynamics of infection and mechanisms of pathogenesis in their natural host. Porcine reproductive and respiratory syndrome virus (PRRSV) is an interesting model to study not only acute infections but also mechanisms of persistence. It is an enveloped positive-sense single-stranded RNA virus that causes late-term reproductive failure and pneumonia in pigs, generating economic losses to the swine industry. Acute infection and virus clearance from serum is typically overcome within 21 to 42 dpi, although in some subjects PRRSV can be detected in tonsils until 250 dpi, indicating that long-term infection is established in lymphoid tissues (Dong et al, 2021).
Hees et al (2019) described that PRRSV persistence in tonsils was associated with high viral loads in serum and some host genes related to susceptibility of infection. They demonstrated in pigs experimentally infected with two PRRSV isolates, NVSL and KS06, a direct correlation between the levels of viremia and quantity of virus in tonsils (Hess et al, 2019). In a most recent study, transcriptome data from tonsils of pigs infected with the KS06 isolate showed augmented transcription of innate immune response genes at 42 dpi in comparison to tonsils from pigs infected with the highly virulent NVSL isolate. In this case, the elevated viremia by KS06 correlated with recruitment of basophils, B cells, and CD8+ central memory T cells, achieving a more efficient viral clearance than that observed in NVSL infection.
Furthermore, 80.9% of virus-positive tonsils were obtained at 42 dpi from nonviremic NVSL infected pigs. A single nucleotide polymorphism in the putative PRRSV resistance-gene GBP5 (guanylate-binding protein 5) showed that the AB genotype was associated with stronger immune response in pigs infected with KS06 than in those infected with NVSL. The study suggested that the NVSL strain inhibits or evades the host immune response and persists in tonsils longer than the KS06 strain (Dong et al, 2021).
During RNA virus replication dsRNA is formed as a replicative intermediate, although some positive-stranded RNA viruses maintain persistence through this nucleic acid structure, in absence of active replication (Klepper et al, 2017). Guo et al (2018) recovered PRRSV RNA mainly in germinal centers of tracheobronchial lymph node (TBLN) and tonsils of pigs infected for 52 dpi that did not show viremia at that moment (5 log10 copies/mg in tonsils and 4 log10 copies/mg in TBLN). Pattern of gene expression in TBLNs was similar between mock and infected pigs along 52 days, excepting for IRF-7 that was upregulated. The plus-strand to minus-strand RNA ratio in TBLNs at 10 dpi was 50.9, while at 52 dpi was 10.81, which correlated with limited viral replication, suggesting that formation of dsRNA is a mechanism that mediates viral latency without induction of strong immune response, revealing a novel mechanism for PRRSV to escape the antiviral response (Guo et al, 2018).
High-throughput sequencing (HTS) combined with metagenomic approaches has identified members of the Picornaviridae family in tonsils of specific pathogen-free pigs (Blomström et al, 2018). In Serogriesen, Sweden, a herd was declared free of swine Influenza A, PRRSV, pseudorabies virus, porcine epidemic diarrhea virus, African swine fever virus, Japanese encephalitis virus, foot and mouth disease virus, rabies virus, classical swine fever virus, swine vesicular disease virus, and transmissible gastroenteritis virus. However, HTS identified more than 87% of pigs positive for Pasivirus A and 25% positive for both Posavirus 1 and Teschovirus. Thus, viral persistence occurs in low to high percentages of apparently healthy animals, making them reservoirs that potentially could transmit the pathogen to the whole herd during reactivation, leading to potential outbreaks (Blomström et al, 2018).
Concluding Remarks
Complete viral clearance from acute infections is not always achieved, leading to persistence in low-to-high percentages of individuals in a population. Viruses without animal reservoirs need to maintain continuous transmission in the human population to be preserved as specie. In the case of seasonal viruses, as many that infect the respiratory tract, possibly establish persistent infections in certain individuals that function at least temporary as reservoirs, undergoing periodic reactivations to transmit the virus to new human hosts. Viral, host, and environmental factors regulate outcome of acute infections and consequences in health. However, our knowledge about which and how these factors interact to induce persistence and its relationship with disease is still partial (Oldstone, 2009). In fact, there exists debate about which viruses maintain long-term infections, principally with respect to those with RNA genomes; outstanding confirmed cases of persistent RNA viruses as measles, hepatitis C, Ebola, and human immunodeficiency virus indicate that this is not an unusual event.
Inoculation of susceptible cell lines with samples obtained consecutively from the same individual to determine presence of infectious viruses has been a tool to evaluate long-term infections. Nevertheless, once adaptive immunity is established, neutralizing antibodies and low viral loads might mask virus presence, making molecular techniques higher valuable and sensible tools to determine persistence, at least of the viral genomes (Fig. 3). Therefore, it is not surprising that many viral infections suspected as persistent have not yet been confirmed, as those affecting the respiratory tract.
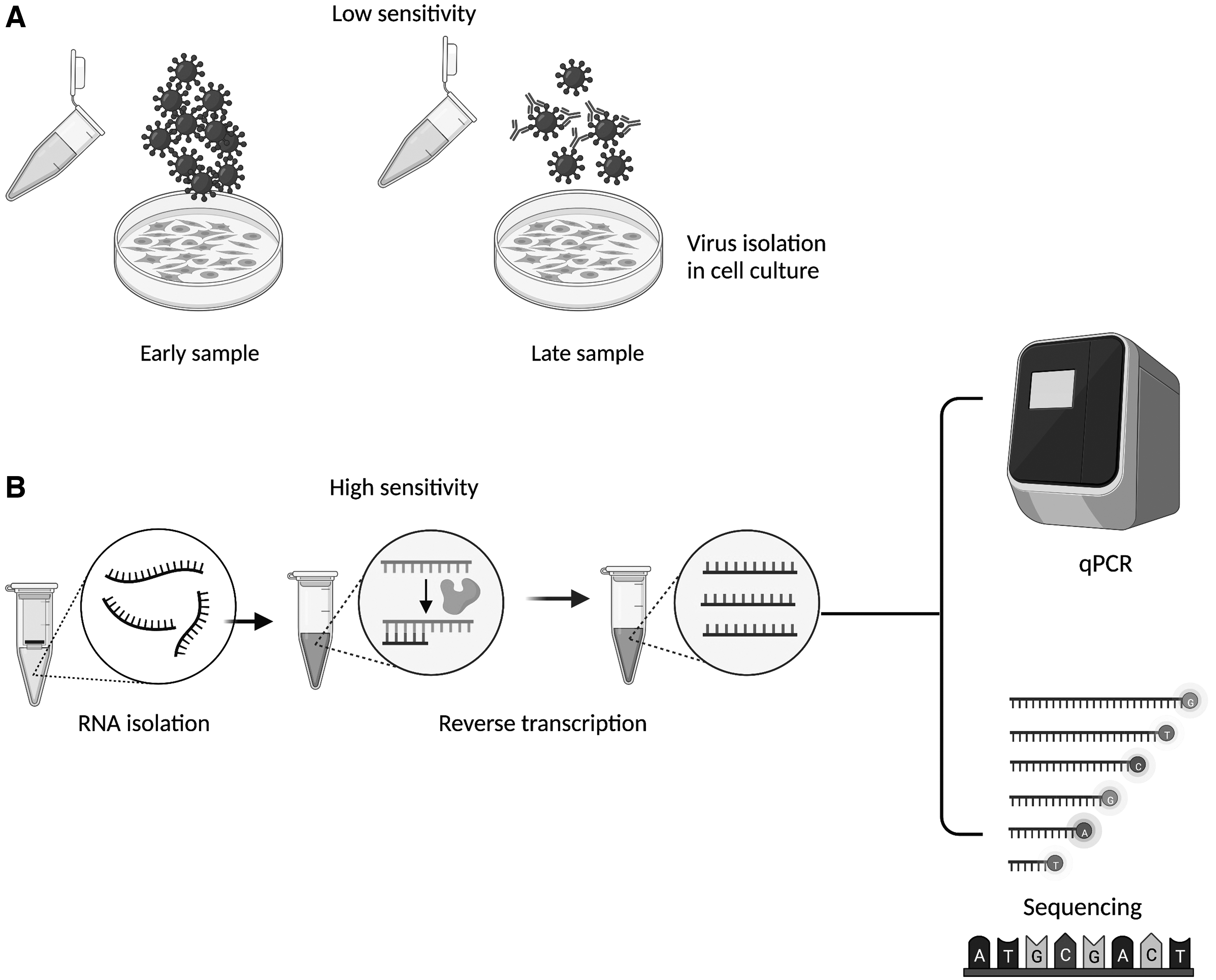
Mechanisms associated with persistent infections entail a greater gene complexity than that in acute infections to succeed in regulation of virus replication and in evasion of host immunity. Under such conditions, it would be ideal to identify particular changes in expression and/or sequence of the viral genome to be used as molecular markers of persistence. In addition, an integrative study considering host genetic background (e.g., simple nucleotide polymorphisms), transcriptomics, and proteomics might result in an integrative network describing how these variables affect health and disease. Thus, multiomics data should be concurrently used and integrated to understand virus–host relationship and map risk pathways to viral persistence and disease.
Footnotes
Acknowledgment
The authors thank Ana Flisser Steinbruch for her support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grant IA205521 from Programa de Apoyo a Proyectos de Investigacion e Innovacion Tecnologica (PAPIIT), Direccion General de Asuntos del Personal Academico (DGAPA), and Faculty of Medicine, Universidad Nacional Autonoma de Mexico (UNAM), Mexico.