
Abstract
To investigate the expression and clinical significance of peripheral blood interleukin (IL)-17A, IL-22, T cell immunoglobulin molecule-3 (Tim-3), and galectin-9 (gal-9) in children with infectious mononucleosis (IM) caused by the Epstein–Barr virus (EBV). Peripheral blood of 54 children with IM (case group) was collected and divided into a liver damage group and a non-liver damage group. During the same period, 20 healthy children were in the control group. IL-17A and IL-22 were measured by enzyme-linked immunosorbent assay. Real-time quantitative polymerase chain reaction was used to measure the mRNA expression of Tim-3 and gal-9. Their correlation with clinical indicators was then analyzed. The IL-17A expression level was higher in the case group than in the control group, while Tim-3, gal-9, and IL-22 were lower than those in the control group. Tim-3 was positively correlated with gal-9, but negatively correlated with IL-17A. Tim-3 and gal-9 were positively correlated with CD4+/CD8+ cells. Conversely, they were negatively correlated with CD3+, CD3+CD8+, white blood cell, lymphocyte (L), alanine transaminase (ALT), aspartate transaminase (AST), glutamyl transpeptidase (GGT), and lactate dehydrogenase (LDH). In the case group, IL-17A was positively correlated with L, GGT, and LDH, but negatively correlated with the natural killer (NK) cell count. IL-17A and IL-22 were positively correlated with CD3+, CD3+CD8+, ALT, and AST, but they were negatively correlated with the ratio of CD4+/CD8+. In the liver damage group, IL-17A, IL-22, CD3+, CD3+CD8+, immunoglobulin A (IgA), IgG, IgM, L, ALT, AST, GGT, LDH, and α-hydroxybutyrate levels were higher than those in the non-liver damage group. However, Tim-3, gal-9, the ratio of CD4+/CD8+, and NK were lower than those in the non-liver damage group. IL-17A, IL-22, Tim-3, and gal-9 are involved in the immune pathogenesis of IM caused by EBV infection in children, which may be related to immune liver injury.
Introduction
In childhood, infectious mononucleosis (IM) was mainly caused by the Epstein–Barr virus (EBV). It is an acute proliferative infectious disease of the mononuclear-macrophage system and associates with immune disorders. EBV infects almost all humans. As the main latent infection site, B lymphocytes are targeted, inducing innate and acquired immune responses (Manet et al., 2021). In the acute phase of IM, the number of regulatory T cells (Tregs) and the immune suppression function are significantly reduced, which is conducive to the activation and proliferation of effector T cells and CD8+ T cells, and the activation and proliferation of cytokines. Increased secretion promotes the clearance of EBV by the host immune system (Wang et al., 2021).
Previous studies have found that there is an imbalance of Treg/T helper (Th)17 in children with EBV infection. During the process of EBV clearance, the proliferation rate of Th17 cells increases, leading to an inflammatory cascade expansion effect (Yang et al., 2021). It is related to autoimmunity. Differentiated Th17 cells can secrete effector cytokines, such as interleukin (IL)-17A, IL-17F, and IL-22. They activate immune cells to produce matrix metallopeptidase, nitrogen monoxide, cytokines, and antimicrobial peptides to clear pathogens (Ding et al., 2022). IL-17A and IL-22, as important signaling molecules in antiviral infection, may play a special role in identifying and attacking invading viruses and pathogens or in self-protection against inflammatory responses (Gong et al., 2021).
T cell immunoglobulin molecule-3 (Tim-3) expression on T cells marks a highly dysfunctional subset of cells characterized by impaired production of antiviral cytokines such as interferon (IFN)-γ and tumor necrosis factor-α and poor proliferative capacity. Low Tim-3 expression on CD4+ T cells contributes to the dysfunctional gal-9-induced apoptosis of CD4+ T cells, thereby producing augmented Th1 and Th17 response (Su et al., 2022). As an inhibitory receptor, Tim-3 binds to gal-9, which mediates cellular calcium influx, suppresses the generation of Th17, and induces immune peripheral tolerance in mice and humans (Wolf et al., 2020). In this study, the expression of IL-17A, IL-22, Tim-3, and gal-9 in peripheral blood of children with IM caused by EBV is determined, and the clinical significance of the Tim-3/gal-9 pathway in IM further explored.
Materials and Methods
Research subjects
A total of 54 children who were hospitalized in our hospital from June 2020 to July 2021, who were confirmed to have IM caused by EBV infection, were enrolled as the case group. They were divided into a liver damage group (28 cases) and a non-liver damage group (26 cases). Twenty healthy children during the same period were enrolled as the control group. (1) The case group was enrolled regarding the diagnosis and treatment principles of major non-neoplastic EBV-related diseases in children (Subspecialty Group of Infectious Diseases, 2016).
(2) Diagnosis of EBV infection (Nowalk and Green, 2016): (1) serological antibody test prompts primary acute EBV infection (meets any of the following three indicators: positive detection of EBV CA IgM and EBV CA IgG antibodies, but negative results for EBV NA IgG antibody and EBV CA IgM antibody, but positive detection of EBV VCA IgG antibody and low affinity; double serum EBV CA IgM antibody titer increased by more than four times); (2) EBV-positive was detected from the patient's serum by polymerase chain reaction (PCR). Inclusion criteria for the liver damage group were as follows: (1) met the IM diagnostic criteria; (2) liver enzyme levels were increased by >twofold. Exclusion criteria were patients with other viral infections within half a year, congenital malformations, severe organ dysfunction, and incomplete clinical data. This study was approved by the hospital's ethics committee and consent was obtained from families. Our study was supported by the Scientific and Technological Development Project (Livelihood Technology) of Suzhou (
Methods
Specimen collection
On the day of admission, venous blood was drawn before the administration of antiviral drugs.
Blood routine test
Two milliliters of patient venous blood was collected in the presence of the anticoagulant ethylenediaminetetraacetic acid (EDTA). The blood routine test was then detected by using the BC-5310 automatic five classification blood cell analyzer (Shenzhen Mindray Biomedical Electronics Co., Ltd.).
Biochemical index and immunoglobulin detection
Collect another 2 mL of patient peripheral blood, use an inert separation gel promoting tube to promote coagulation, and use Siemens ADVIA 2400 automatic biochemical analyzer to detect serum alanine transaminase (ALT), aspartate transaminase (AST), glutamyl transpeptidase (GGT), lactate dehydrogenase (LDH), α-hydroxybutyrate (α-HBDH), and prealbumin (PA). Mindray BS2000 uses immunoturbidimetry to detect the levels of immunoglobulins IgA, IgG, and IgM.
Analysis of peripheral blood lymphocyte subsets
First, use a vacuum ethylenediaminetetraacetic acid anticoagulant tube to extract 2 mL of venous blood. Then, 20 μL of CD3−FITC/CD16+CD56−PE/CD45PerCP-Cy5.5/CD4−PC7/CD19−APC/CD8−APC-Cy7 (Beijing Tong sheng Times Biotechnology Co. Ltd.) fluorescent monoclonal antibody is added to the bottom of the flow tube. Using reverse pipetting technology, draw 50 μL of whole blood to the bottom of the tube, mix well, place at room, and avoid light for 15–25 min. Add 450 μL of hemolytic agent for blood cell analysis to the tube, mix, and place at room for 15 min. Flow cytometry (FACS Canto II, BD) and its automatic analyzer were used to detect CD3+, CD3+CD4+, CD3+CD8+, CD19+CD23+, and CD16+CD56+cell proportion (Fig. 1).
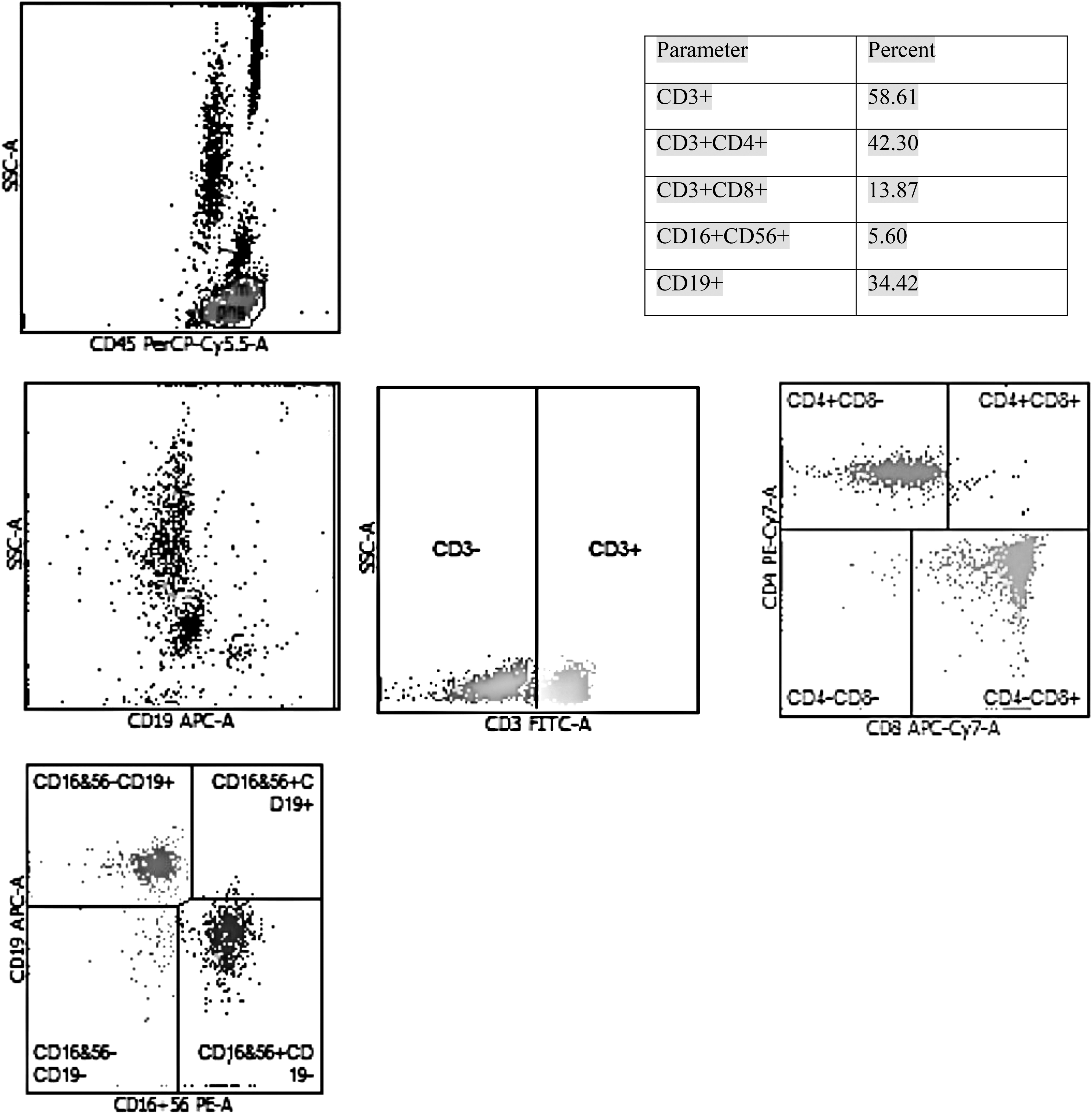
Detection of lymphocyte T cell subsets and NK cells by flow cytometry. Manual gating of lymphocyte region was performed based on high expression of CD45 low SSC. Select and display lymphocyte gate cells, then set up CD3/SSC scatter plots, CD3/CD4 scatter plots, CD3/CD8 scatter plots, CD3/CD16+CD56 scatter plots, and CD3/CD19 scatter plots, respectively. NK, natural killer; SSC, side scatter complexity.
Detection of IL-17A and IL-22
Plasma was separated from venous blood and IL-17A and IL-22 levels were measured by enzyme-linked immunosorbent assay kits (double antibody sandwich method, Xinbosheng Biotechnology Co., Ltd) in a microplate reader.
Analysis of Tim-3 and gal-9 mRNA expression
Peripheral blood samples were centrifuged in a low-temperature, high-speed centrifuge at 1,200 r/min at 4°C for 5 min. Cells were resuspended in 0.5 mL of TRIzol to extract total RNA and then reverse transcribed to synthesize cDNA. A SYBR Green PCR Kit (Novizan Biotechnology) was used to perform real-time PCR with β-actin as the internal reference to measure expression of target mRNA. Expression differences of each gene were compared by the 2−ΔΔCt relative expression method. Primer sequences are listed in Table 1.
Primer Sequences
gal-9, galectin-9; Tim-3, T cell immunoglobulin molecule-3.
Statistical methods
Statistical data were analyzed by SPSS 26.0. Measurement data with a normal distribution are expressed as the mean ± standard deviation. The t-test was used for comparison between groups. Pearson correlation was used for correlation analysis. Data not conforming to a normal distribution are represented by the median (P25–P75). The Mann–Whitney U-test was used for comparison between groups. Spearman correlation was used for correlation analysis. p < 0.05 was considered statistically significant.
Results
Clinical information
There were 54 children in the case group, including 36 males and 18 females, with an average age of 4.48 ± 2.08 years, and 20 children in the control group, including 12 males and 6 females, with an average age of 4.60 ± 2.12 years. There were no significant differences in sex or age between the two groups (p > 0.05). There were 28 children in the liver damage group, including 15 males and 13 females, with an average age of 4.77 ± 1.36 years, and 26 children in the non-liver damage group, including 21 males and 5 females, with an average age of 4.83 ± 1.41 years. There was no significant difference in sex or age between the two groups (p > 0.05).
Comparison of blood routine and biochemical tests between case and control groups
The white blood cell (WBC) count, lymphocyte (L) count, GGT, LDH, ALT, and AST in the case group were higher than those in the control group. The platelet (PLT) count and PA were significantly lower than those in the control group (p < 0.05). There was no significant difference in the neutrophil (N) count or α-HBDH between the two groups (p > 0.05, Table 2).
Comparison of Blood Routine and Biochemical Between Case and Control Groups
α-HBDH, α-hydroxybutyrate; ALT, alanine transaminase; AST, aspartate transaminase; GGT, glutamyl transpeptidase; L, lymphocyte; LDH, lactate dehydrogenase; N, neutrophil; PA, prealbumin; PLT, platelet; WBC, white blood cell.
Comparison of immune-related indicators between the case group and the control group
The absolute counts of CD3+, CD3+CD4+ and CD3+CD8+ cells and IL-17A expression were higher than those in the control group, Tim-3, gal-9, IL-22, and the ratio of CD4+/CD8+ were significantly lower than those in the control group (p < 0.05). There were no significant differences in CD3−CD19+, CD19+CD23+ counts, IgA, IgG, or IgM levels between the two groups (p > 0.05, Table 3).
Comparison of Immune Indicators Between Case and Control Groups
IgA, immunoglobulin A; IL, interleukin; NK, natural killer.
Correlation between Tim-3, gal-9, IL-17A, IL-22, and other indicators in the case group
In the case group, Tim-3 was positively correlated with gal-9 and negatively correlated with IL-17A. Tim-3 and gal-9 were positively correlated with the ratio of CD4+/CD8+ and negatively correlated with CD3+, CD3+CD8+, WBC, L, ALT, AST, GGT, and LDH (p < 0.05). There was no correlation between Tim-3 and IL-22, CD3+CD4+ cells, natural killer (NK) cells, PLT, and PA (p > 0.05). There was no correlation between gal-9 and IL-17A, IL-22, CD3+CD4+, NK cells, PLT, and PA (p > 0.05). In the case group, IL-17A was positively correlated with L, GGT, and LDH and negatively correlated with the NK count.
In addition, IL-17A and IL-22 were positively correlated with CD3+ and CD3+CD8+ counts, ALT, and AST and negatively correlated with the ratio of CD4+/CD8+ (p < 0.05). IL-17A did not correlate with gal-9, IL-22, CD3+CD4+, WBC, PLT or PA (p > 0.05). IL-22 did not correlate with Tim-3, gal-9, IL-17A, CD3+CD4+, NK cells, WBC, L, PLT, GGT, LDH or PA (p > 0.05, Table 4).
Correlation Between T Cell Immunoglobulin Molecule-3, Galectin-9, Interleukin (IL)-17A and IL-22, and Other Indicators in the Case Group
Comparison of blood routine and biochemical tests between liver and non-liver damage groups
In the liver damage group, L, ALT, AST, GGT, LDH, and α-HBDH were significantly higher than those in the non-liver damage group (p < 0.05). There were no significant statistical differences in WBC, N, PLT, or PA between the two groups (p > 0.05, Table 5).
Comparison of Blood Routine and Biochemical Tests Between Liver and Non-Liver Damage Groups
Comparison of immune-related indicators in the liver and non-liver damage groups
In the liver damage group, IL-17A, IL-22, CD3+, CD3+CD8+, IgA, IgG, and IgM were higher than those in the non-liver damage group, while Tim-3, gal-9, the ratio of CD4+/CD8+, and NK count were significantly lower than those in the non-liver damage group (p < 0.05). There were no significant differences in CD3+CD4+, CD3−CD19+, and CD19+CD23+ cells between the two groups (p > 0.05, Table 6).
Comparison of Immune-Related Indicators in Liver and Non-Liver Damage Groups
Discussion
After EBV infection, B lymphocytes proliferate in large numbers, express specific antigen molecules, induce innate and acquired immune responses, and promote immune clearance of the body. The innate immune cells—dendritic cells (DC)—present EBV antigen and stimulate initial T cells. The cells differentiate into cytotoxic T cells (Tcs), helper T cells, and Tregs (Gujer et al., 2019). Therefore, the counts of CD3+, CD3+CD4+, and CD3+CD8+ in the case group were higher than those in the control group. In addition, DC can also secrete IL-12, IFN-α, and IFN-β, increasing the killing activity of NK cells (Png et al., 2021).
This study found that the NK count in the case group was higher than that in the control group, which was consistent with previous research results (Ye et al., 2021). In the acute phase of IM, CD8+ T cells significantly proliferate and specifically recognize EBV lysate and latent antigen (Wang et al., 2021), and so, CD4+/CD8+ is lower than that of the control group, suggesting that CD8+ T cells play an important role in the process of virus clearance in the acute phase of IM.
The expression of IL-17A was higher in the case group. Combined with correlation analysis, IL-17A was positively correlated with CD3+ and CD3+CD8+ counts. In different cytokine environments, CD8+ T cells proliferate significantly and produce subpopulations, including Tc2, Tc9, Tc17, and CD8+Tregs. The cytokine IL-17A produced by the Tc17 subtype can aggravate autoimmunity and participate in viral infection immunity (Wu et al., 2013). The Tc17 subset can clear influenza virus (Hamada et al., 2013) and hepatitis B virus (Jiang et al., 2022) through the Fas/FasL pathway. It is speculated that significantly proliferating CD8+ T cells participate in the pathological immune response of EBV through the production of IL-17A. In the peripheral blood of patients with chronic active Epstein–Barr virus infection caused by repeated EBV infection, IL-17A mostly comes from the differentiated CD4+Th17 cell (Ohta et al., 2013).
The percentage of Th17 cells in the peripheral blood of children with IM is significantly higher than that of the control group (Yang et al., 2021). Primary EBV infection can trigger CD4+ T cells to activate and recognize viral antigens. IL-17A can also come from the CD4+ T cell subset Th17 cells. Next, we need to isolate and enrich Tc17 cells and Th17 cells, and further evaluate the source of IL-17A through intracellular staining.
Th17 cells can also secrete IL-22, but the role of IL-22 and IL-17A is different. In Th17 cells, TGF-β, IL-17, and costimulation pathways all inhibit IL-22 secretion (Meng et al., 2012). Therefore, IL-22 in the case group was lower than that in the control group. IL-22 can bind to the IL-22 and IL-10 receptors and promote the repair of damaged cells through the Janus kinase-signal transducers and activator of transcription signal pathway. IL-22 can also enhance the secretion of antimicrobial peptides and enhance the ability of epithelial cells to clear pathogenic bacteria (Zenewicz, 2018), suggesting that IL-22 is related to the body's defense ability.
As a negative regulatory molecule, Tim-3 is activated by gal-9, inducing cellular calcium influx and apoptosis, thereby inhibiting the inflammatory response. Tim-3 is expressed by several immune cell populations, such as terminally differentiated Th1, Th17, and Tc17 lymphocytes, while Th22 do not express Tim-3 (Su et al., 2022). The expression of Tim-3 and gal-9 was decreased in the case group. Th17 function was not inhibited and the secretion of IL-17A increased. In the case group, correlation analysis between Tim-3, gal-9, IL-17A, and IL-22 and immune indicators showed that Tim-3 and gal-9 were negatively correlated with CD3+CD8+ cell count, but not significantly correlated with CD4+ T cells. In addition, they were positively correlated with CD4+/CD8+, suggesting that the binding of Tim-3 to gal-9 has the potential to contribute to the functional inactivation of CD8+ T cells in persistent viral infections.
Latent membrane protein 2 (LMP2A) mediates upregulation of the F3 gene through the phosphatidylinositol-3-kinase (PI3K)/AKT signaling pathway, causing PLT aggregation. Besides, PLTs are reduced by secondary hypersplenism (Cen et al., 2018) and anti-PLT antibodies (Hsieh et al., 2007). Comparing blood routine tests, it was found that the WBC and L counts in the case group were higher than those in the control group, which were related to clean virus. The lower PLT count was associated with the hypersplenism and antiplatelet antibodies.
Approximately 51.9% of children with EBV infection have elevated liver enzymes (Zhang et al., 2021), which are closely related to CD8+ cells (Zhang et al., 2020). EBV infects CD8+ T cells, combines with specific adhesion molecules expressed by hepatic Kupffer cells, and directly kills target cells or induces apoptosis of target cells through the Fas/FasL and perforin/granzyme pathways (Ge et al., 2020), thereby causing liver damage. Therefore, ALT and AST in the case group were higher than those in the control group. Cytokines associated with EBV infection damage bile canaliculus cell and interfere with the transportation function of the hepatic sinus and canaliculus.
Therefore, GGT in the case group was higher than that in the control group. LDH is an important enzyme system of anaerobic glycolysis and gluconeogenesis. Upon tissue damage, LDH increases in serum, which is associated with tumor progression (Li et al., 2019a). The increased LDH in the case group is related to tissue damage after virus infection. PA is synthesized by hepatocytes with a half-life of only ∼1.9 days, and its concentration in plasma has high sensitivity for estimating malnutrition (Ingenbleek, 2022) and hepatic insufficiency (Li et al., 2019b). PA in the case group was lower than that in the control group, indicating recent liver function impairment caused by EBV infection.
The Tim-3/gal-9 pathway is involved in the immunopathological processes of IM, which may be related to liver damage. We further divided the case group into liver and non-liver damage groups. In the liver damage group, IL-17A and IL-22 expression and CD3+ and CD3+CD8+ cell counts were higher than those in the non-liver damage group, indicating that the hepatic damage caused by EBV was related to lymphocyte infiltration and inflammatory factors. The increase of α-HBDH, LDH, ALT, AST, and GGT can indicate liver damage.
In the liver damage group, the expression of Tim-3 and gal-9 that promoted apoptosis of inflammatory cells was less than that in the non-liver damage group, while the CD8+ T cell count was higher. IM is a proliferative disease of B lymphocytes, with elevated levels of IgA, IgG, and IgM. The NK cell count in the liver damage group was lower, which was consistent with the changes of NK cells during chronic EBV infection (Maakaroun et al., 2010). It suggests that the immune function in the liver damage group is significantly disordered.
Summary
We confirmed that IL-17A, IL-22, Tim-3, and gal-9 are involved in the immunopathogenesis of IM caused by EBV in children, which may be related to immune liver injury. We can further explore its mechanism by the major molecules that enhance or inhibit this pathway at the cellular level.
Footnotes
Acknowledgment
We thank Mitchell Arico from Liwen Bianji (Edanz) (
Authors' Contributions
M.X. carried out the molecular genetic studies, participated in sequence alignment, and drafted the article; Y.L. and M.C. performed data analysis and selected the topic; image processing was done by Z.J. W.Z. reviewed and revised the article. All the authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by the Scientific and Technological Development Project (Livelihood Technology) of Suzhou (no. SYS2019085).