
Abstract
Despite the great success of vaccines in various infectious diseases, most current vaccines are not effective enough, and on the contrary, clinically approved alum adjuvants cannot induce sufficient immune responses, including a potent cellular immune response to confer protection. In this study, we used Nanochelating Technology to develop novel nanoadjuvants to boost the potency of the alum-adjuvanted inactivated severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) vaccine. BALB/c mice were immunized twice over 2 weeks with different doses of adjuvanted-vaccine formulations and immune responses were assessed. The analysis results of IFN-γ and IL-17 cytokines demonstrated the effectiveness of the nanoadjuvants produced by the Nanochelating Technology in shifting the alum-based vaccine toward a stronger Th1 pattern. In addition, these nanoadjuvants improved IL-2 cytokine response, which shows the efficacy of these novel formulations in inducing specific T lymphocyte proliferation. Using these nanoadjuvants increased IL-10 cytokine secretion that may be representative of a better immunoregulatory impact and may also potentially prevent immunopathology responses. Moreover, specific IgG titer analysis revealed the potency of these nanoadjuvants in improving humoral immune responses. The enzyme-linked immunosorbent assay of receptor-binding domain (RBD)-specific IgG response showed that the developed novel formulations induced strong IgG responses against this protein. This study shows that the nanostructures produced by the Advanced Nanochelating Technology have potent adjuvant effects on alum-based SARS-CoV-2 vaccines to not only compensate for alum weakness in inducing the cellular immune responses by smart regulation of the immune system but also significantly improve the humoral and cellular immune responses simultaneously.
Introduction
Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) infects the respiratory tract, resulting in pneumonia and acute respiratory distress syndrome in the majority of cases (De Greef et al., 2021; Vajari et al., 2021). SARS-CoV-2 is one of the three coronaviruses in the last two decades, which has caused serious infectious respiratory disease and mortality in the world (Qiu et al., 2020; Ruiz-Hitzky et al., 2020). Scientific endeavors to develop vaccines have resulted in the development of several vaccines that have been approved for human use (Kim et al., 2021; Tregoning et al., 2020). These vaccines are developed by inactivated-, subunit protein-, viral vector- and mRNA-based approaches (Burki, 2020; Dagan et al., 2021; Kim et al., 2021); however, the most serious concern in vaccine development is the efficacy as well as the duration of protection regarding some technologies such as inactivated and recombinant vaccines (Paltiel et al., 2021; Voysey et al., 2021).
Inactivated and recombinant subunit vaccines are not immunogenic enough, so they need adjuvants to boost the immunogenicity of vaccine antigens and induce protective immunity (Coffman et al., 2010). In fact, adjuvants are one of the components of recombinant and inactivated vaccine formulations and the potency of a vaccine in the induction of immune response is somehow dependent on the nature of the adjuvants that are used in the vaccine formulation (Mahdavi et al., 2011). Although clinically approved alum adjuvants improve humoral immune responses and antigen stability (Oleszycka and Lavelle, 2014), they are not potent inducers of cellular immune responses and Th1-type cytokines in human vaccines, so they are not capable of inducing effective protection against some pathogens (Petrovsky and Aguilar, 2004). Under such circumstances, it seems that using efficient adjuvants with higher immunomodulatory impact and efficient Th1 immune stimulation could be useful for increasing vaccine potency.
Nanochelating Technology is a new approach that has demonstrated positive effects when used in drug and vaccine formulations in the past decade. The structures synthesized based on this technology have a range of compositions, shapes, and surface properties with higher efficiency, lower toxicity, and significant modulatory impact on the immune system and are therefore promising for designing novel vaccine formulations (Fakharzadeh et al., 2014; Fakharzadeh et al., 2013; Hafizi et al., 2019; Kalanaky et al., 2016). The previous study on the effect of the first synthesized nanoadjuvant Hep-C on the hepatitis B vaccine showed its immunomodulatory effect that could improve the cellular immune responses (Fakharzadeh et al., 2013).
In light of the lower efficiency of inactivated SARS-CoV-2 vaccines, especially in the induction of Th1 response, the novel nanoadjuvants of S1, Hep-C2, and F13 were synthesized based on the Nanochelating Technology. These nanoadjuvants were then analyzed to investigate their impact on increasing the potency of the inactivated SARS-CoV-2 vaccine. Therefore, experimental mice were immunized with these novel vaccine formulations first, and then their immunologic parameters were assessed.
Materials and Methods
SARS-CoV-2 virus
SARS-CoV-2 (SARS-CoV-2/human/IRN/Alborz-IR/2020) was cultured on a Vero cell line in Dulbecco's modified Eagle's medium supplemented with 3% heat-inactivated fetal bovine serum (FBS) in a cell culture room at biosafety level-III facilities. After observing the cytopathic effect on the Vero cell line starting 1 day after infection, the culture supernatant was harvested for identification or purification. The virus was inactivated with formalin (1/3,000 vol/vol at room temperature). Following the clarification of the cell debris and ultrafiltration, purification was performed using PEG-1500 and column chromatography. The purified, inactivated viruses were dialyzed versus phosphate-buffered saline (PBS), passed through a 0.22 μm filter, quantified using the Bradford method, and finally stored at −70°C until use (SARS-CoV-2/human/IRN/Alborz-IR/2020 surface glycoprotein (S) gene, complete cds 3,822 bp linear RNA MW709393.1 GI: 1998262910) (Kaffashi et al., 2021).
Nanoadjuvants synthesis and characterization
Three different adjuvants of Hep-C2, S1, and F13 nanoparticles with three different chemical structures were designed and synthesized based on the Advanced Nanochelating Technology patented in the United States Patent and Trademark Office (2012) (Patent No. US8288587B2) (Department of Research and Development of Sodour Ahrar Shargh Company, Tehran, Iran). Based on the well-known impact of selenium and iron on the immune system function (Arthur et al., 2003; Mahdavi et al., 2017a; Seligman et al., 1992; Thorson et al., 1991), three nanoadjuvants were designed and synthesized for this study; Hep-C2 nanoadjuvant with iron chelating property, selenium-containing S1 nanoadjuvant, and iron-containing F13 nanoadjuvant.
Hep-C2 characterization
Hep-C2 is a molecule with a molecular weight of 380.22 Da and 38 atoms, where we used a polymerized form as a nanoadjuvant. According to dynamic light scattering (DLS) particle size distribution analysis at the KEFA laboratory, the zeta potential of this molecule is 6,931 mV indicating its high hydrodynamic stability (Fig. 1a). The power distance index (PDI) of 0.676 proves the accuracy of the analysis (Nano-ZS, Red Badge, ZEN 3600 Device; Malvern Instrument).

Dynamic light scattering analysis of the nanoadjuvants produced by the nanochelating technology. Hep-C2
S1 characterization
S1 nanoadjuvant is a chelated selenium-containing nanoparticle. As can be seen in the transmission electron microscopy (TEM) image, the molecular size of this nanoparticle is 30–40 nm (Fig. 2a). According to the DLS analysis, 100% of the molecules are within 22 nm range showing the hydrodynamic property of this nanoadjuvant (Fig. 1b). The PDI of 0.44 also proves the accuracy of the test.

TEM images of nanoadjuvants. The S1
F13 characterization
F13 nanoadjuvant is a chelated iron-containing nanoparticle. As the TEM image shows, the molecular size of this nanoparticle is ∼25–30 nm (Fig. 2b), and the DLS analysis also confirms that ∼30% of the nanoparticles are within 30 nm range. In addition, the reported PDI of 0.37 certifies the accuracy of the test (Fig. 1c).
Nanoadjuvants median lethal dose (LD50)
The LD50s of Hep-C2, S1, and F13 nanoadjuvants are 1776.59, 54.59, and 1688.5 mg/kg, respectively (assessed according to the standard Organization for Economic Co-operation and Development guidelines).
Vaccine formulations
Inactivated SARS-CoV-2 virus was formulated in alum hydroxide adjuvant at molecular grade (Brenntag Biosector, Denmark) according to the isoelectric point. To formulate vaccines, 2 μg of the inactivated SARS-CoV-2 virus was admixed with 800 μg of alum hydroxide adjuvant in PBS buffer and was shaken at 100 rpm for 60 min, and then the mixture was left to allow the viral particles to be adsorbed on to the surface of alum gel. After loading the virus on the surface of the alum gel, Hep-C2 (0.02, 2 and 800 μg/dose), F13 (0.4, 2 and 800 μg/dose), and S1 (0.02 and 2 μg/dose) nanoadjuvants were admixed with the vaccine in different tubes. When the vaccines were completely formulated, 200 μL of each vaccine, containing 2 μg of the inactivated SARS-CoV-2 virus, was used for immunization.
Experimental groups and immunization
Six- to eight-week-old male BALB/c mice (N = 79) were purchased from the Pasteur Institute of Iran (Tehran, Iran). The mice were maintained in 12-h light and 12-h dark cycles for 1 week before the experiments while they had free access to food and water. The experimental mice were randomly divided into 10 groups and were injected subcutaneously. Each group containing mice was defined as follows: Group 1: 2 μg of inactivated SARS-CoV-2 + Alum + 0.02 μg of Hep-C2 (n = 7 mice) Group 2: 2 μg of inactivated SARS-CoV-2 + Alum + 2 μg of Hep-C2 (n = 7 mice) Group 3: 2 μg of inactivated SARS-CoV-2 + Alum +800 μg of Hep-C2 (n = 8 mice) Group 4: 2 μg of inactivated SARS-CoV-2 + Alum + 0.4 μg of F13 (n = 7 mice) Group 5: 2 μg of inactivated SARS-CoV-2 + Alum + 2 μg of F13 (n = 7 mice) Group 6: 2 μg of inactivated SARS-CoV-2 + Alum +800 μg of F13 (n = 8 mice) Group 7: 2 μg of inactivated SARS-CoV-2 + Alum + 0.02 μg of S1 (n = 7 mice) Group 8: 2 μg of inactivated SARS-CoV-2 + Alum + 2 μg of S1 (n = 7 mice) Group 9: 2 μg of inactivated SARS-CoV-2 + Alum as control (n = 7 mice) Group 10: PBS as control group (n = 6 mice)
In addition to the experimental mice groups, an untreated group (n = 8) was considered to obtain both anti-receptor-binding domain (RBD) IgG and anti-inactivated SARS-CoV-2 IgG cutoff points through enzyme-linked immunosorbent assay (ELISA) in this group to calculate specific IgG titer in the experimental groups. The experimental mice were immunized subcutaneously twice at a 14-day interval and their immunologic parameters were assessed 2 weeks after the second immunization. The experimental mice were all cared for according to Iran Ministry of Health and Medical Education guidelines (
In vitro antigen recall responses of spleen cells
Two weeks after the second immunization, the spleens of the immunized mice were removed under sterile conditions and were then mechanically dissected and suspended in sterilized cold penicillin- and streptomycin-containing PBS. The cell suspension was centrifuged at 300 g/5 min, RBCs were lysed using lysis buffer, and single-cell suspension was washed three times before being adjusted to 3 × 106 cells/mL in RPMI-1640 (Gibco, Germany) supplemented with 10% FBS, 4 mM
The spleen cells were then stimulated in vitro by adding 1 μg/mL of the inactivated SARS-CoV-2 virus, and the plates were incubated at 37°C in 5% CO2. After 60 h of postantigen restimulation, the culture supernatants were centrifuged at 2,000 rpm/10 min, and the supernatants were harvested and stored at −70°C for cytokines assay.
IFN-γ, IL-4, IL-2, TNF-α, IL-17, IL-10, and TGF-β cytokines ELISA
IFN-γ, IL-4, IL-2, TNF-α, IL-17, IL-10, and TGF-β cytokines were measured by mouse cytokine ELISA kits (Mabtech, Stockholm, Sweden) according to the manufacturer's instruction. The mouse cytokines were evaluated using ELISA along with the cytokines standard and using the standard curve of each cytokine, the pg/mL of each sample was reported. To calculate IFN-γ/IL-4 and IFN-γ/TNF-α ratios, the quantity of IFN-γ of each mouse was divided by its IL-4 and/or TNF-α cytokine, respectively.
ELISA of specific IgG antibodies titer and their isotypes
Specific antibodies were determined by an optimized indirect ELISA. In brief, 100 μL of 5 μg/mL of the inactivated SARS-CoV-2 in PBS was added to 96-well ELISA Maxisorp plates (Greiner, Germany) and incubated at 4°C overnight. The wells were washed with 0.1% Tween 20–containing PBS (washing buffer) and blocked at 37°C for 60 min with 2% skimmed milk in PBS + 0.1% Tween 20 (blocking buffer). The plates were washed with the washing buffer and 100 μL of 1/25 diluted sera up to 16 serial dilutions to be then added into each well and incubated at 37°C for 90 min. The wells were washed five times with the washing buffer and incubated for 2 h with 100 μL of 1/8,000 dilution of anti-mouse conjugated to horseradish peroxidase (HRP; Razirad, Iran). Following that, the wells were washed five times and incubated for 30 min with 100 μL of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate in the dark.
The reaction was then stopped with 100 μL of 2 N H2SO4 and color density was measured at A450nm with an ELISA plate reader. The ELISA cutoff point for each experimental dilution of the sera samples in the untreated group was calculated by the mean + 3 standard deviation (SD) formula, so the titer of each mouse in the experimental groups was obtained based on the cutoff. The final results of the specific IgG antibody responses for every individual mouse were presented as specific IgG titers. In addition, the specific IgG1 and IgG2a subclasses were detected using goat anti-mouse IgG1 and IgG2a secondary antibodies (Sigma) according to the manufacturer's instruction.
Assessment of specific IgG anti-RBD protein
Specific IgG antibodies against RBD protein were determined by an optimized indirect ELISA.
In brief, 100 μL of 2 μg/mL of RBD protein (The Native Antigen Company, United Kingdom) in carbonate-bicarbonate buffer with pH 9.6 was added to 96-well ELISA Maxisorp plates (Greiner) and incubated at 4°C overnight. The wells were washed with the washing buffer and blocked at 37°C for 60 min with 1.5% bovine serum albumin in PBS + 0.05% Tween 20 (blocking buffer). The plates were washed five times with the washing buffer, and 100 μL of 1/50 diluted sera of the experimental vaccinated mice and/or the mice in the untreated group were added to each well and incubated at 37°C for 90 min.
Afterward, the wells were washed five times with the washing buffer and incubated for 90 min with 100 μL of 1/8,000 dilution of anti-mouse conjugated to HRP (Razirad). The wells were washed five times again and incubated for 10 min with 100 μL of TMB substrate in the dark. The reaction was then stopped with 100 μL of 2 N HCl, and color density was measured at A450nm with an ELISA plate reader. The RBD-ELISA cutoff point of the sera samples in the untreated group was calculated by the mean + 3SD formula and then the IgG response to RBD for every individual mouse was reported as a ratio by the formula “OD of RBD ELISA of individual mouse/cutoff.”
Statistical analysis
The data were presented as mean ± standard error of the mean of each experiment. The statistical significance of differences was analyzed using Mann–Whitney U test (GraphPad Prism 6.01 software; GraphPad Software, La Jolla, CA). In all cases, p < 0.05 were considered significantly different.
Results
IFN-γ cytokine
IFN-γ cytokine response analysis showed that immunization with all the experimental vaccines resulted in a significant increase in the level of this cytokine (Fig. 3a) as compared with the PBS group (p < 0.0043). Immunization with the inactivated SARS-CoV-2 + Alum + 2 μg Hep-C2 formulation resulted in a 124.60% increase in IFN-γ cytokine secretion as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.3660). Immunization with the inactivated SARS-CoV-2 + Alum + 0.4 μg F13 also showed a significant increase in IFN-γ compared with the inactivated SARS-CoV-2 + Alum group (p = 0.0140). In addition, immunization with the inactivated SARS-CoV-2 + Alum + 2 μg S1 resulted in a substantial increase in IFN-γ response as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.0029).

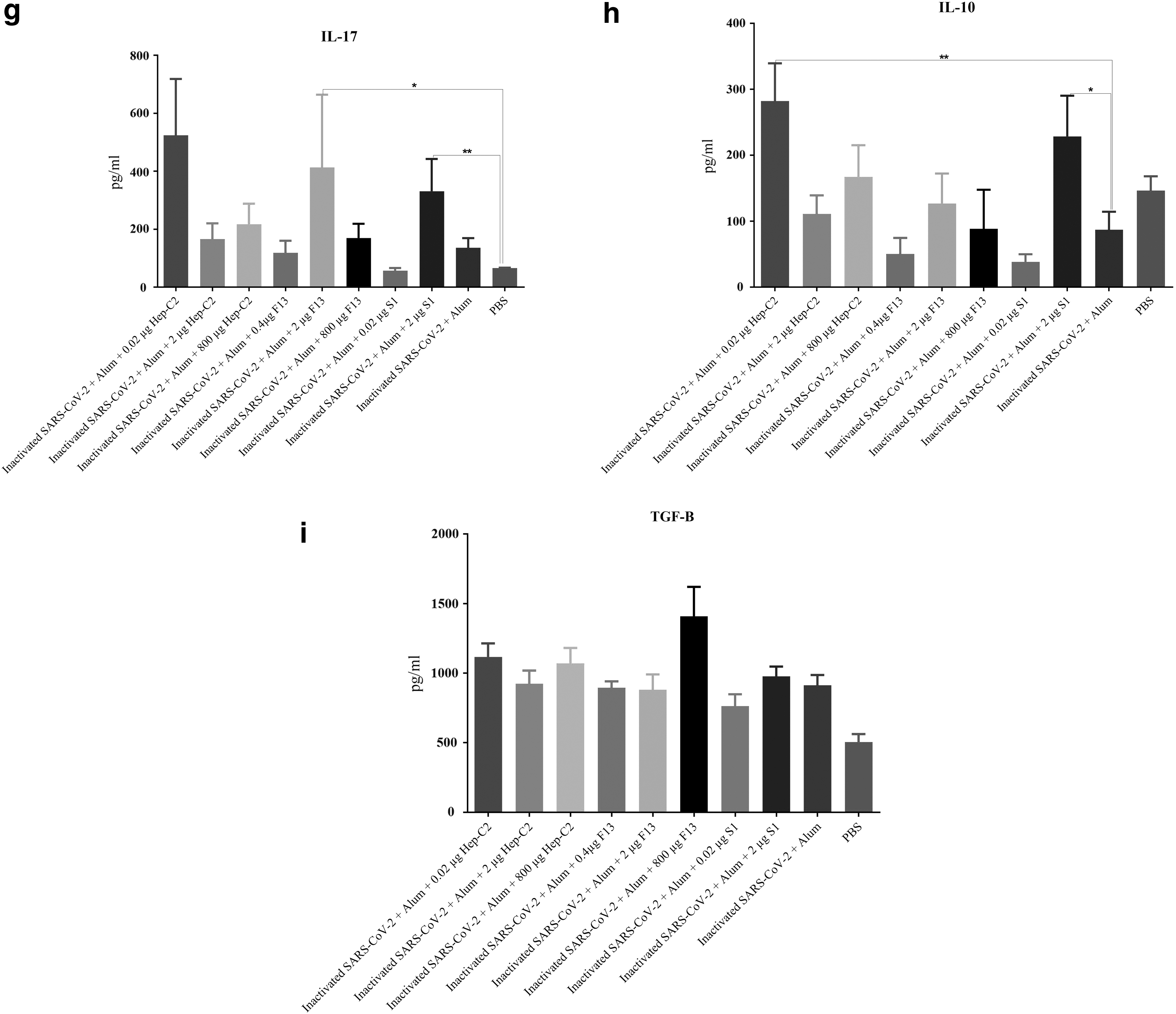
Cytokine responses of experimental group after immunization by different vaccine formulations. Mice spleen cell suspension was prepared and recalled in vitro with the inactivated SARS-CoV-2, and then the culture supernatant was used for cytokines assessment by quantitative ELISA.
IL-4 cytokine
The experimental mice injected with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2, inactivated SARS-CoV-2 + Alum + 2 μg Hep-C2, inactivated SARS-CoV-2 + Alum + 800 μg Hep-C2, inactivated SARS-CoV-2 + Alum + 0.02 μg S1, and inactivated SARS-CoV-2 + Alum + 2 μg S1 indicated a significant increase in IL-4 cytokine secretion compared with the PBS group (p < 0.0303).
In addition, immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2, inactivated SARS-CoV-2 + Alum + 800 μg Hep-C2, and inactivated SARS-CoV-2 + Alum + 2 μg S1 led to a significant increase in IL-4 cytokine response as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.0105, p = 0.0421, and p = 0.0017, respectively). However, the other vaccine formulations did not show any significant difference when compared with the inactivated SARS-CoV-2 + Alum group (p > 0.4043; Fig. 3b).
IFN-γ/IL-4 ratio
The analysis of the IFN-γ/IL-4 ratio demonstrated that immunization with different vaccine formulations resulted in a significant increase in the IFN-γ/IL-4 ratio in comparison with the PBS group (p < 0.0022). Immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 increased the IFN-γ/IL-4 ratio by 43.32% in comparison with the inactivated SARS-CoV-2 + Alum group (p = 0.2343). In addition, the inactivated SARS-CoV-2 + Alum +800 μg F13 and inactivated SARS-CoV-2 + Alum + 2 μg S1 also increased the IFN-γ/IL-4 ratio by 6.91% and 100.51% respectively compared with the inactivated SARS-CoV-2 + Alum group (p = 0.6126 and p = 0.0728, respectively; Fig. 3c).
TNF-α cytokine
The analysis results indicated that the TNF-α cytokine secretion level of the mice immunized with different vaccine formulations increased significantly as compared with the PBS group (p < 0.0043). Immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 and inactivated SARS-CoV-2 + Alum +800 μg Hep-C2 increased TNF-α response by 30.97% and 28.58%, respectively, in comparison with the inactivated SARS-CoV-2 + Alum group (p = 0.3829 and p = 0.8665, respectively). Similarly, immunization with the inactivated SARS-CoV-2 + Alum + 2 μg F13 and inactivated SARS-CoV-2 + Alum + 2 μg S1 resulted in a 14.76% and 45.25% increase in TNF-α cytokine response, respectively, as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.9015 and p = 0.0379, respectively; Fig. 3d).
IFN-γ/TNF-α ratio
The results analysis showed that the IFN-γ/TNF-α ratio of the mice immunized with different vaccine formulations increased significantly in comparison with the PBS group (p < 0.0043). The inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 increased IFN-γ/TNF-α ratio by 18.15% in comparison with the inactivated SARS-CoV-2 + Alum group (p = 0.2677). In addition, the inactivated SARS-CoV-2 + Alum + 2 μg S1 vaccine also resulted in an increase of 56.21% in IFN-γ/TNF-α ratio as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.0262; Fig. 3e).
IL-2 cytokine
IL-2 cytokine analysis of the experimental groups revealed that different vaccine formulations resulted in a significant increase in the level of this cytokine as compared with the PBS group (p < 0.0043). The experimental mice immunized with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 vaccine demonstrated a 21.23% increase in IL-2 cytokine secretion compared with the inactivated SARS-CoV-2 + Alum group (p = 0.6943). In addition, the injection of the inactivated SARS-CoV-2 + Alum + 2 μg S1 vaccine increased IL-2 cytokine level by 85.62% compared with the inactivated SARS-CoV-2 + Alum group (p = 0.1206; Fig. 3f).
IL-17 cytokine
The analysis of IL-17 cytokine response in the experimental groups showed that immunization with the inactivated SARS-CoV-2 + Alum (p = 0.0635), inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 (p = 0.0584), inactivated SARS-CoV-2 + Alum +800 μg F13 (p = 0.0386), and inactivated SARS-CoV-2 + Alum + 2 μg S1 (p = 0.0023) resulted in an increase in the level of this cytokine in comparison with the PBS group. In addition, immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2, inactivated SARS-CoV-2 + Alum + 2 μg F13, and inactivated SARS-CoV-2 + Alum + 2 μg S1 increased IL-17 cytokine response by 287.05%, 204.99%, and 144.55%, respectively, compared with the inactivated SARS-CoV-2 + Alum group (p = 0.1690, p = 0.8735 and p = 0.1783, respectively; Fig. 3g).
IL-10 cytokine
The analysis of IL-10 cytokine response in the experimental mice demonstrated that the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 and inactivated SARS-CoV-2 + Alum + 0.02 μg S1 increased IL-10 cytokine secretion in comparison with the PBS group (p = 0.0649 and p = 0.0043, respectively). In addition, immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2 and inactivated SARS-CoV-2 + Alum + 2 μg S1 resulted in a significant increase in IL-10 cytokine response in comparison with the inactivated SARS-CoV-2 + Alum group (p = 0.0082 and p = 0.0245, respectively; Fig. 3h).
TGF-β cytokine
The results analysis showed that, except for the inactivated SARS-CoV-2 + Alum + 0.02 μg S1, there was a significant increase in TGF-β cytokine secretion in the experimental mice immunized with different vaccine formulations as compared with the PBS group (p < 0.0163). However, there was no significant difference between the experimental vaccinated groups and the inactivated SARS-CoV-2 + Alum group (p > 0.0721; Fig. 3i).
Specific IgG titer against the inactivated virus
The results analysis showed that there was a significant increase in the specific IgG titer level in the mice immunized with different vaccine formulations compared with the PBS group (p < 0.0043). There was an 89.58% increase in the specific IgG titer in the group vaccinated with the inactivated SARS-CoV-2 + Alum +800 μg Hep-C2 in comparison with the inactivated SARS-CoV-2 + Alum group (p = 0.0970). Besides, there was a 75.00% increase in the specific IgG titer level of the mice immunized with the inactivated SARS-CoV-2 + Alum + 2 μg F13 as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.1166). Vaccination with the inactivated SARS-CoV-2 + Alum + 2 μg S1 also increased the specific IgG titer level by 16.66% compared with the inactivated SARS-CoV-2 + Alum group (p = 0.3677; Fig. 4a, b).
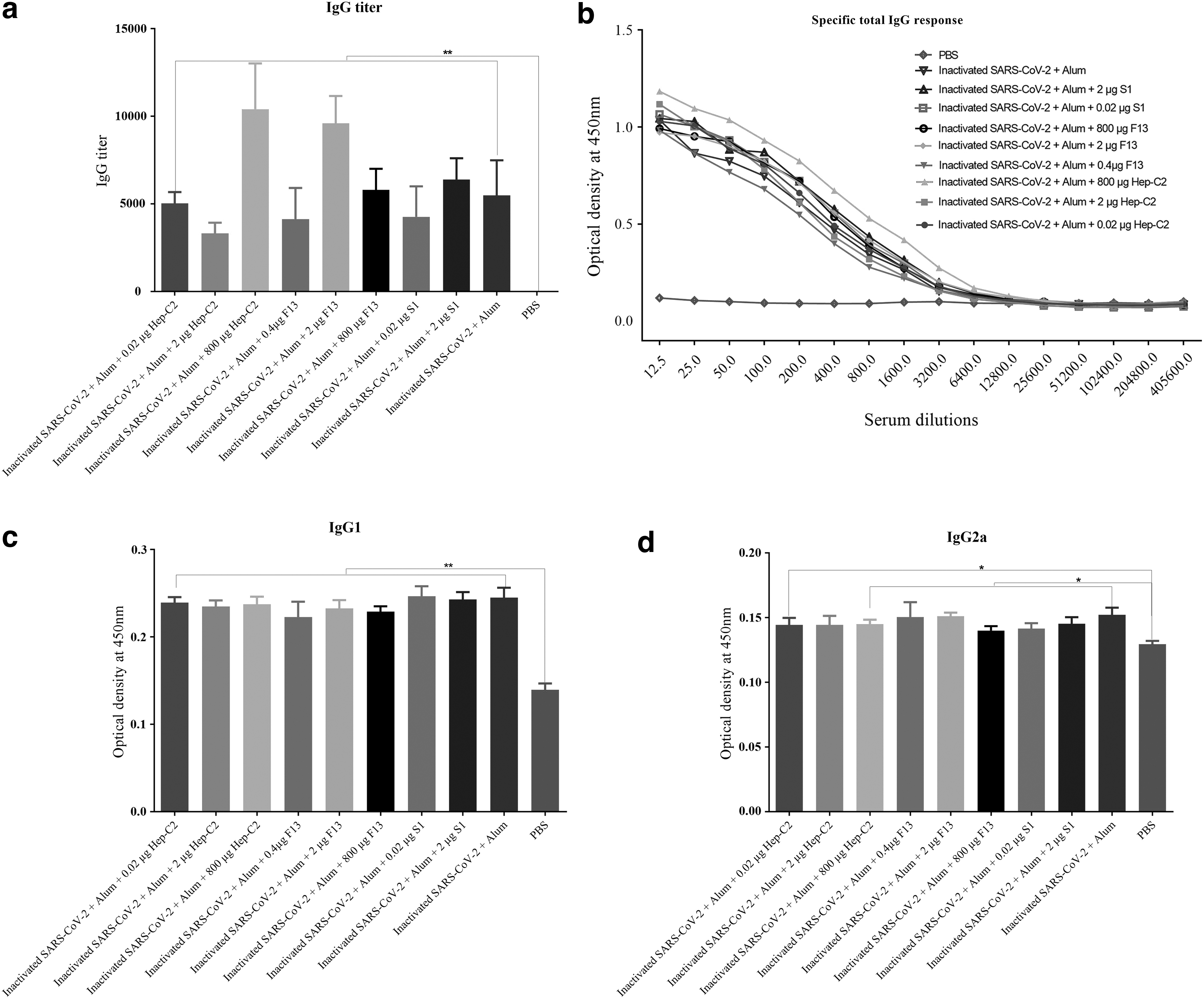
Specific total IgG responses measurement of the experimental mice after two times immunization at a 2-week interval. Inactivated SARS-CoV-2 virus was coated on ELISA plates and the specific total IgG was measured by an optimized ELISA.
Specific IgG1 antibody
The results indicated a significant increase in the specific IgG1 antibody isotype of the mice immunized with different vaccine formulations as compared with the PBS group (p < 0.0022). Immunization with the inactivated SARS-CoV-2 + Alum + 0.4 and 800 μg F13 resulted in a decrease in the IgG1 response as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.0140 and p = 0.0107, respectively). However, there was no significant difference between the other vaccine formulations and the inactivated SARS-CoV-2 + Alum group in terms of specific IgG1 antibody isotype (p > 0.0903; Fig. 4c).
Specific IgG2a antibody
The analysis of the specific IgG2a antibody isotype demonstrated that, except for the inactivated SARS-CoV-2 + Alum + 2 μg Hep-C2 (p = 0.0787 vs. PBS), there was a significant increase in the level of this antibody in the mice immunized with different vaccine formulations compared with the PBS group (p < 0.0473). However, there was no significant difference between the different vaccine formulations and the inactivated SARS-CoV-2 + Alum group (p > 0.0775; Fig. 4d).
Specific IgG antibodies against RBD protein
The results of the specific IgG antibodies analysis against RBD protein demonstrated that immunization with the inactivated SARS-CoV-2 + Alum resulted in a response lower than the cutoff point, whereas the inactivated SARS-CoV-2 + Alum formulated with 0.02, 2 and 800 μg of Hep-C2 nanoadjuvant resulted in an increase of 49.14%, 35.21%, and 38.72% in the specific IgG antibodies, respectively, compared with the inactivated SARS-CoV-2 + Alum group (p = 0.3176, p = 0.0262, and p = 0.1206). In addition, immunization with the inactivated SARS-CoV-2 + Alum + 0.4 μg F13 showed a 56.38% increase in the specific IgG antibodies compared with the inactivated SARS-CoV-2 + Alum group (p = 0.2949). The inactivated SARS-CoV-2 + Alum + 0.02 μg S1 increased anti-RBD response by 27.97% as compared with the inactivated SARS-CoV-2 + Alum group (p = 0.4685; Fig. 5).
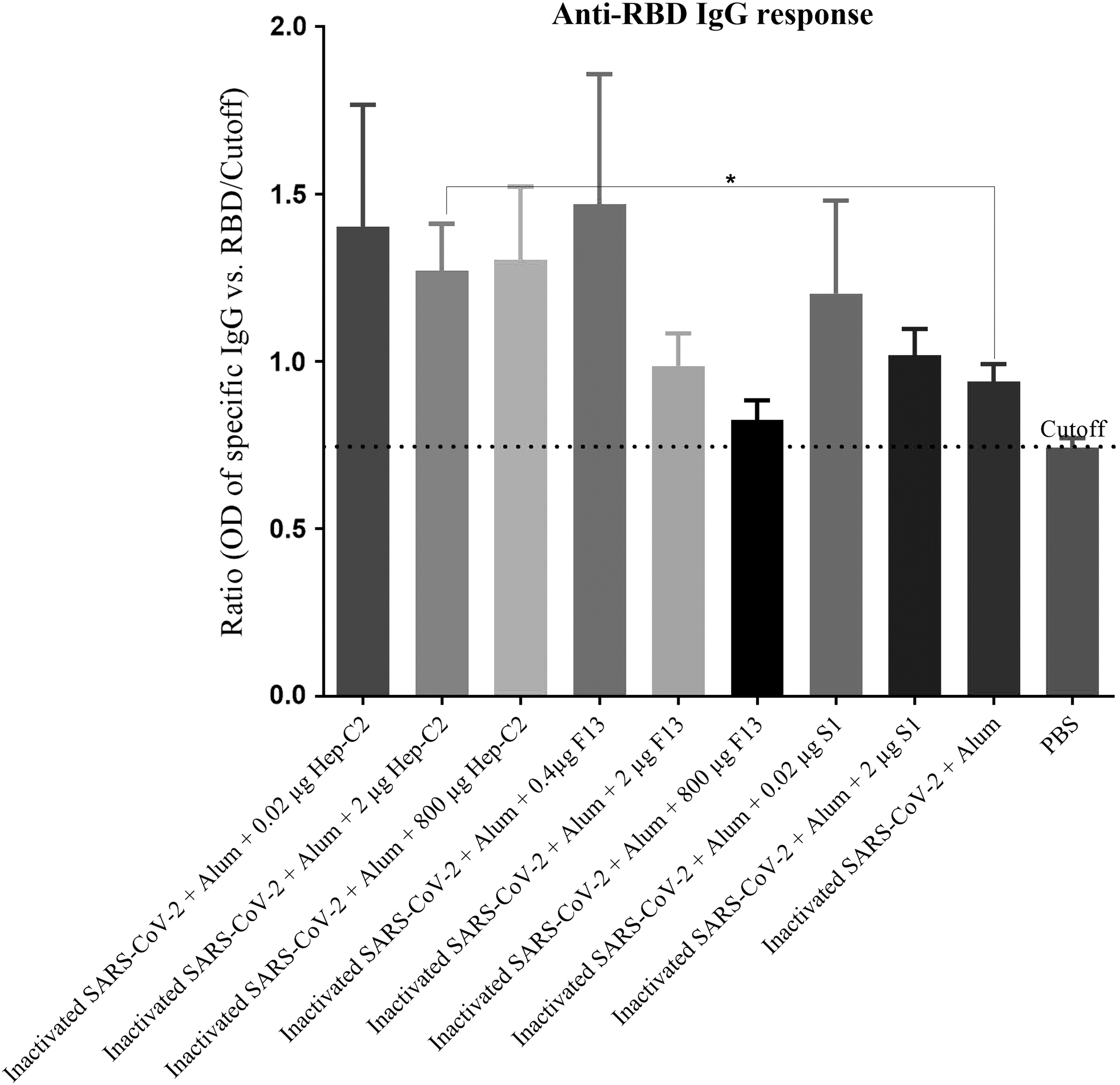
Specific IgG antibodies against RBD protein measurement of the experimental mice after two times immunization at a 2-week interval. Anti-RBD IgG response was determined by an optimized indirect ELISA. The IgG response to RBD for each individual mouse was reported as ratio by the formula “OD of RBD ELISA of individual mouse/cutoff.” Immunization with the inactivated SARS-CoV-2 + Alum resulted in a response lower than the cutoff point, whereas the inactivated SARS-CoV-2 + Alum formulated with 0.02, 2 (*p < 0.05) and 800 lg of Hep-C2 nanoadjuvant resulted in 49.14%, 35.21%, and 38.72% increase in the specific IgG antibodies, respectively, compared with the inactivated SARS-CoV-2 + Alum group. In addition, immunization with the inactivated SARS-CoV-2 + Alum +0.4 μg F13 resulted in a 56.38% increase in the specific IgG antibody compared with the inactivated SARS-CoV-2 + Alum group (p = 0.2949). The inactivated SARS-CoV-2 + Alum +0.02 μg S1 also increased anti-RBD response by 27.97% as compared with the inactivated SARS-CoV-2 + Alum group. The data are presented as mean ± SEM. p-Values were analyzed by Mann–Whitney U test (*p < 0.05). RBD, receptor-binding domain.
Discussion
As coronaviruses cause serious infections, both in humans and animals, and are able to spread quickly (García-Azorín et al., 2021; Kwok, 2021), developing efficient vaccines to inhibit COVID-19 infection is deemed to be the most rational approach (Sanyal et al., 2021).
Several vaccines have already been approved, but their side effects (Schultz et al., 2021), protective efficacy, long-term immune responses, and the new variants of the virus are major concerns (Yan et al., 2021). Because inactivated viral organisms are not generally highly immunogen, they need to be adjuvanted to boost their effectiveness and immune responses (Lima et al., 2004; Wang and Xu, 2020). In this study, three different nanoadjuvants of Hep-C2, F13, and S1 were developed based on the Advanced Nanochelating Technology to assess their impact on increasing the potency of the inactivated SARS-CoV-2 vaccine formulations.
IFN-γ is an immunoregulatory cytokine with broad-spectrum antiviral and antimicrobial functions influencing multiple cells and cellular activities (Billiau and Matthys, 2009; Kang et al., 2018). Recent studies on the treatment of COVID-19 patients with IFN-γ have represented the pivotal role of this cytokine in restricting virus replication and distribution (Hooks et al., 2003). IFN-γ is, therefore, recently suggested to be used for COVID-19 treatment, particularly for endangered patients when standard therapies are not effective (Noh, 2020). On the contrary, Th1 cell responses contribute to protection against SARS-CoV-2, whereas the key component of this response is the production of IFN-γ by T cells (Lange et al., 2021).
Studies have revealed that protection against COVID-19 infection is highly dependent on the Th1 and IFN-γ cytokine responses (Kalimuddin et al., 2021; Tan et al., 2021; Wang et al., 2021). Despite the widely approved importance of Th1 and IFN-γ responses in protection against viruses, alum adjuvants are weak inducers of this response and can even be Th1 suppressors (Oleszycka et al., 2018). So, finding a proper strategy to overcome such a limitation is essential.
The results analyses of this study revealed that adding the nanochelating-based adjuvants to the inactivated SARS-CoV-2 vaccine increased IFN-γ cytokine response substantially. This finding demonstrates the potency of these nanoadjuvants in the polarization of alum-based vaccines toward a more robust Th1 pattern. The results of our previous work on the HBsAg vaccine model were also in line with the findings of this study where Hep-C another nanochelating-based adjuvant demonstrated high potency in the induction of robust IFN-γ cytokine response.
Because it is, at present, believed that a mixture of the Th1 and Th2 immune responses are essential for the optimum immune responses against viral infections, IL-4 cytokine response, as the hallmark of Th2, was also assessed in this study (Hasan et al., 2021; Hellerstein, 2020; Mahdavi et al., 2010; Prompetchara et al., 2021). The results indicated that immunization with the inactivated SARS-CoV-2 + Alum + 0.02 μg Hep-C2, inactivated SARS-CoV-2 + Alum +800 μg Hep-C2, and inactivated SARS-CoV-2 + Alum + 2 μg S1 led to a significant increase in IL-4 cytokine response compared with the inactivated SARS-CoV-2 + Alum group. This finding shows that Hep-C2 and S1 nanoadjuvants were able to improve the Th2 as well as Th1 immune responses. It means, a mixture of the Th1/Th2 immune responses with these novel formulations was achievable. These results are similar to the outcome of the previous study on the HBsAg vaccine, where Hep-C nanochelating-based adjuvant could improve IL-4 cytokine response in the HBsAg vaccine (Fakharzadeh et al., 2013).
Immunization with the inactivated SARS-CoV-2 + Alum vaccine adjuvanted by Hep-C2 and S1 resulted in an increase in IFN-γ/IL-4 ratio compared with the inactivated SARS-CoV-2 + Alum group, which stresses the Th1 polarization (Mahdavi et al., 2017a). Similarly, Hep-C2 and S1 nanoadjuvant increased IFN-γ/TNF-α ratio compared with the inactivated SARS-CoV-2 + Alum group, indicating the potency of these two nanoadjuvants in the induction of higher levels of the IFN-γ and Th1 immune responses with lower inflammatory response level. This means that potentially lower immunopathology responses may occur when vaccines are adjuvanted by these nanoadjuvants (Prabhavathi et al., 2015; Xu et al., 2019) as one other advantage of these nanoadjuvants that can trigger higher cellular immune responses with a lower inflammatory response (Prabhavathi et al., 2015).
Evidence shows that IL-2 cytokine is considered a key factor in combating viral infections through the clonal expansion of T cells and activation of natural killer cells (Coppola et al., 2020; Wiedemann et al., 2020; Xie et al., 2019; Zhou et al., 2020). The results of this study showed that adding Hep-C2 and S1 to the inactivated vaccine resulted in a 21.23% and 85.62% increase in the secretion of this cytokine. Considering the role of IL-2 as a T cell proliferation factor (Fathi et al., 2021), this finding shows the potency of these nanoadjuvants in the induction of specific T lymphocyte proliferation.
Studies have revealed that IL-17 is necessary for the clearance of some bacterial and viral infections through the recruitment and activation of macrophages in infection sites (Gurczynski et al., 2019; Li and Sun, 2018; Mahdavi et al., 2017b; Shibue et al., 2019). In addition, the Th17 lineage is critical for vaccine-induced memory immune responses against infectious diseases (Lin et al., 2010). Immunization with the inactivated SARS-CoV-2 + Alum adjuvanted by Hep-C2, F13 and S1 nanoadjuvants increased IL-17 response. Therefore, these nanoadjuvants may be able to generate effective vaccine-induced long-lived immunity against viruses, which is going to be assessed in the future studies.
One major challenge to the application of novel adjuvants is keeping a balance between the proinflammatory and adaptive immune responses (Audibert and Lise, 1993; Gnjatic et al., 2010; Tom et al., 2019). As a result of this, limiting the side effects of such inflammation is the major obstacle to the use of adjuvants in human vaccines. IL-10 cytokine is a potent anti-inflammatory and pleiotropic mediator that protects lungs from early immune-mediated damage and interference with viral clearance in COVID-19 infection (Lindner et al., 2021).
In this study, IL-10 response analysis showed a significant increase in the level of this cytokine in the Hep-C2 and S1 groups in comparison with the inactivated SARS-CoV-2 + Alum group.
It should be noted that as IL-10 cytokine is well-known for hemostasis induction in the immune system and considering the increase in the immunogenic responses (i.e., increase in the other immunologic parameters) of these two experimental groups, it can be claimed that an increase in IL-10 cytokine response may reflect a better immune system activation owing to the pleiotropic effects on immune cells and/or a higher immunoregulatory effect that can potentially prevent the immunopathology responses (Fathi et al., 2021; Mollazadeh et al., 2019). In parallel to this, the results of the TGF-β cytokine response in different vaccine formulations showed no significant change at the level of this cytokine, revealing the fact that the nanoadjuvants did not have any suppressive effects on the immune responses.
The specific IgG response to the inactivated SARS-CoV-2 vaccine was examined in the current investigation to determine the effect of this new vaccine formulation on humoral immune responses. The results showed that Hep-C2, F13, and S1 formulated vaccines increased the specific IgG titer by 89.58%, 75.00%, and 16.66%, respectively, compared with the inactivated SARS-CoV-2 + Alum vaccine. This finding shows the impact of these nanoadjuvants on the improvement of humoral immune responses. In light of the fact that each isotype of IgG antibody has a distinct function in the immune responses, it seems that all the vaccine formulations induced both the IgG1 and IgG2a responses, which may show the positive effect of these novel nanoadjuvants on the quality of the humoral immune responses (Velashjerdi Farahani et al., 2016).
It is well proven that the specific IgG response plays a key role in the prevention and protection against COVID-19 infection through binding to RBD of SARS-CoV-2 virus (Khoury et al., 2021; Starr et al., 2021), so the anti-RBD IgG responses of the experimental groups were assessed as well. The anti-RBD response analysis of the novel vaccine formulations showed that the nanoadjuvants increased this response ranging from 27.97% up to 56.38% as compared with the inactivated SARS-CoV-2 + Alum group.
This finding demonstrates the potency of these novel nanoadjuvants in the augmentation of the specific IgG responses against RBD sequences that are potentially critical in the SARS-CoV-2 virus. This is a critical property in adjuvants as evidence shows that antibodies against RBD sequences are correlated with protection against COVID-19 infection (Papenburg et al., 2021). However, a challenging study in an animal model will reveal this property in an in vivo model.
Conclusions
In conclusion, the results of this study show that the nanostructures produced by the Nanochelating Technology have a potent adjuvant activity on the inactivated alum-based SARS-CoV-2 vaccine and are able to induce both the cellular and humoral immune responses. However, the dose of nanoadjuvants in the vaccine formulation is an important parameter and affecting on the nature and level of immune responses. As a result, they can be promising candidates to be introduced as novel adjuvants for human alum-based vaccines to overcome their limitations and improve their immunogenicity. In our next study, we aim at finding the efficacy of the inactivated SARS-CoV-2 vaccine when formulated with these nanoadjuvants.
Ethical Approval
Iran Ministry of Health and Medical Education Ethics Committee (Approval Code: IR.SBMU.NRITLD.REC.1400.104).
Footnotes
Acknowledgments
The authors thank the Department of Research and Development at Sodour Ahrar Shargh Company, Borna Zist Pajoohan Knowledge-based Company, Dr. Rohullah Keshavarz and Dr. Morteza Taghizadeh from Razi Vaccine and Serum Research Institute of Iran for their kind help in this project.
Authors' Contributions
S.K., S.F., M.H., and M.H.N. are from Sodour Ahrar Shargh Company (Tehran, Iran); P.K. is from Islamic Azad University (Tehran, Iran); H.J. is from Shahid Beheshti University of Medical Sciences (Tehran, Iran); S.M.H. and M.M. are from Pasteur Institute of Iran (Karaj, Iran); and A.K. is from Razi Vaccine and Serum Research Institute (Karaj, Iran), all where research and education are the primary functions.
Author Disclosure Statement
Hep-C2, F13 and S1 nanoadjuvants were designed and synthesized using the Nanochelating Technology, invented by M.H.N. and patented in the United States Patent and Trademark Office (US8288587B2). The authors of this research have no conflict of interest.
Funding Information
No funding was received for this article.