
Abstract
This study aimed to investigate the changes of toll-like receptor 4 (TLR4), proinflammatory cytokine expression, hepatitis B virus surface antigen (HBsAg), and hepatitis B virus envelope antigen (HBeAg) expression as well as innate immune cell percentages in a mouse model of persistent hepatitis B virus (HBV) infection to better understand the innate immune response. Mouse models of persistent HBV infection, HBsAg expression, and HBeAg expression were developed using high-pressure tail-vein injection of recombinant adeno-associated viruses. Enzyme-linked immunosorbent assays (ELISAs) were used to determine the serum proinflammatory cytokine levels. Immunohistochemistry and western blot assays were used to detect TLR4 expression. Flow cytometric analysis was used to assess the percentage of innate immune cells in the whole blood. Persistent HBV infection, HBsAg expression, and HBeAg expression each significantly decreased the expression of TLR4. Persistent HBV infection significantly increased the percentages of T cells and monocytes, whereas it decreased the percentage of natural killer (NK) cells. Persistent HBeAg expression also decreased the percentage of NK cells, whereas persistent HBsAg expression increased the percentage of NK cells. Both persistent HBsAg and HBeAg expression increased the percentage of monocytes. However, both persistent HBsAg and HBeAg expression decreased the percentage of T cells. HBV as well as HBsAg and HBeAg showed similar effects on the expression of TLR4 and proinflammatory cytokines as well as the percentage of monocytes. Persistent HBV infection increased the percentage of T cells and decreased the percentage of NK cells, whereas only persistent HBeAg expression contributed to a decreased percentage of NK cells.
Introduction
Hepatitis B virus (HBV) is a partially double-stranded circular DNA virus of ∼3.2 kb in length (Yan et al., 2022). The HBV genome contains four overlapping open reading frames: PreC/C, P, PreS/S, and X, encoding HBV core antigen and hepatitis B virus envelope antigen (HBeAg), HBV DNA polymerase, hepatitis B virus surface antigen (HBsAg), and HBV X protein, respectively (Li et al., 2020; Yan et al., 2022). HBV infection can cause acute and chronic hepatitis, which can progress into cirrhosis and even hepatocellular carcinoma. The World Health Organization (2022) has estimated that there are 296 million people living with chronic HBV infection, with the highest rates of infection in Africa and Asia, resulting in ∼820,000 deaths in 2019. Most chronic HBV carriers acquire the virus through vertical transmission, that is, from their carrier mothers during birth and delivery. On the contrary, horizontal transmission (exposure to infected blood) often leads to self-limited acute HBV infection (Tsai et al., 2018).
The natural history of chronic HBV infection comprises the following phases: immune-tolerant phase, immune-reactive phase, low replicative phase, and reactivation phase (Sarin et al., 2016). HBV clearance mainly depends on the antiviral ability of the host's immune system. For HBV infection in adults, >90% of cases will resolve, manifesting as self-limited acute infection, during which the innate immune system is the primary contributor (Zhong et al., 2021).
Toll-like receptors (TLRs) are well-defined pattern recognition receptors that can recognize specific pathogen-associated molecular patterns to trigger the innate immune response to activate antiviral mechanisms, including intracellular antiviral pathways and the production of interferons (IFNs) and proinflammatory cytokines (e.g., interleukin [IL]-1β, IL-6, tumor necrosis factor [TNF]-α, and transforming growth factor [TGF]-β) (Tsai et al., 2018; Zhang and Lu, 2015). The activation of many TLRs can induce IFNs. Although HBV infection always results in no or weak TLR-mediated innate immunity, studies have demonstrated that activation of the TLR system in hepatic cells can suppress HBV replication (Ma et al., 2015).
Liver injury in chronic hepatitis B may increase TLR2 and TLR4 levels (Akbal et al., 2017). Among all TLRs, TLR4 is an important trigger of the immune response against HBV infection and liver injury, and it plays a pivotal role in viral clearance during HBV infection (Ma et al., 2015; Wu et al., 2014). In addition, a TLR4-dependent immune response, but not HBV reactivation, is important in radiation-induced liver disease resulting from liver cancer radiotherapy (Wu et al., 2014). Natural killer (NK) cells, which respond rapidly to viral infections, are important effectors of innate immunity. The activities of NK cells are reported to be impaired in patients with chronic HBV infection (Ghosh et al., 2016; Tjwa et al., 2011). Moreover, ineffective HBV-specific T cell responses are responsible for persistent HBV replication (Gehring and Protzer, 2019).
Although chronic HBV infection is related to the aforementioned innate immune system or effectors such as TLR4 and NK cells, the identification of which HBV protein might be involved in these processes has not been determined. This study aimed to investigate the changes of TLR4 expression, proinflammatory cytokine (IL-1β, IL-6, TNF-α, and TGF-β, and IFN-γ) levels, and innate immune cell (T cells, NK cells, and monocytes) percentages in the development of chronic HBV infection in a mouse model. In addition, mouse models with persistent HBsAg or HBeAg expression were evaluated.
Materials and Methods
Cells, plasmid construction, and transfection
Hepatocellular carcinoma HuH7 cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in Dulbecco's modified Eagle medium (Gibco) supplemented with 10% fetal bovine serum (Gibco), 1% penicillin, and 1% streptomycin at 37°C in 5% CO2. The pAAV-1.3HBV plasmid containing the 1.3-mer HBV genome (genotype B, serotype ayw) was purchased from Beijing FivePlus Molecular Medicine Institute Co. Ltd. The surface (s) gene fragment was amplified from pAAV-1.3HBV (Forward: 5′-TCGATAAGCTTAGAAGATCTCAATCTCGGGAATCTCAATGTTAGTATTC-3′, reverse: 5′-GCCAGTGAATTCGAGCTCTCGAATAGAAG-3′) and inserted into the HindIII-SacI restriction sites of pAAV-1.3HBV to construct the recombinant pAAV-HBs plasmid.
The HBV pAAV-HBe plasmid was cloned by inserting the HBV envelope (e) gene fragment amplified from pAAV-1.3HBV (Forward: 5′-AAAGGTACCATGCAACTTTTTCACCTCTGC-3′, reverse: 5′-AAACTCGAGCTAACATTGAGATTCCCGAGATTGAGATCT-3′) into the KpnI-XhoI restriction sites of pAAV-MCS. The recombinant plasmids were confirmed by sequencing. For transfection, HuH7 cells were seeded in a 24-well plate; transfected with the pAAV-1.3HBV, pAAV-HBe, or pAAV-HBs plasmid using Lipofectamine 3000 (Invitrogen); and cultured for another 48 h. The supernatants were collected for enzyme-linked immunosorbent assays (ELISAs).
Animals and development of chronic HBV infection in mice
Six-week-old specific pathogen-free (SPF) C57 mice were purchased from China Three Gorges University. The mice were kept at an SPF facility under a room temperature of 22–26°C, a humidity of 50–60%, and a 12-h light/dark cycle, and the animals had free access to food and water ad libitum.
It has been reported previously that a chronic HBV infectious mouse model can be developed through adeno-associated virus (AAV)-mediated delivery of 1.2 copies of the HBV genome (Ye et al., 2015). Many studies have confirmed the sustained and efficient transgene expression after a one-time administration of recombinant AAVs (Bass-Stringer et al., 2018; Ohmori, 2020; Wang et al., 2000). In this study, 48 male mice were randomly divided into 4 groups: pAAV-MCS plasmid (control, n = 12), pAAV-1.3HBV (n = 12), pAAV-HBs (n = 12), and pAAV-HBe (n = 12). The mice in all four groups received a high-pressure tail-vein injection of 10 μg of pAAV-MCS plasmid, pAAV-1.3HBV, pAAV-HBs, or pAAV-HBe, respectively, within 4–6 sec.
Serum samples were collected at 14 days, 2 months, 4 months, and 6 months after the injection. Two weeks after the last blood collection, the mice were anesthetized with 1% pentobarbital sodium, and serum and liver tissues were collected. All animal protocols were approved by the Institutional Animal Care and Use Committee of the local institution. Our animal experiments complied with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978).
Enzyme-linked immunosorbent assay
The levels of secreted HBsAg and HBeAg in the culture supernatants and mouse sera were determined using commercial HBe/HBs ELISA kits (Kehua, Shanghai, China), according to the manufacturer's instructions. The serum levels of TNF-α (No. MU30030, sensitivity: ≤1.6 pg/mL), IFN-γ (No. MU30038, sensitivity: ≤2 pg/mL), IL-6 (No. MU30034, sensitivity: ≤1.2 pg/mL), TGF-β (No. MU30574, sensitivity: ≤3 pg/mL), and IL-1β (No. MU30369, sensitivity: ≤2.4 pg/mL) were assessed using commercial ELISA kits (Bioswamp, Wuhan, China), according to the manufacturer's instructions.
Quantitative polymerase chain reaction of HBV DNA
The total genomic DNA was extracted from the liver tissues using a DNA extraction kit (Aidlab Biotechnologies, Beijing, China), according to the manufacturer's instructions. The quantitative polymerase chain reaction (qPCR) was conducted using a CFX-Connect 96 Realtime System (Bio-Rad). The reaction system was as follows: SYBR FAST qPCR master mix, 10 μL; forward primer, 0.5 μL; reverse primer, 0.5 μL; DNA template, 1 μL; ddH2O, 8 μL. The following procedure was used for the qPCR: initial denaturation at 95°C for 3 min, followed by 40 cycles of 15 sec at 95°C and 30 sec at 60°C, and concluding with a melting-curve stage of 15 sec at 95°C, 1 min at 60°C, and 15 sec at 95°C.
The HBV DNA copies, HBs genes, or HBe genes were quantified using standard curves of the pAAV plasmids. The primers used were as follows: HBV-F, 5′-TGGATTCGCACTCCTCCAGCTT-3′, HBV-R, 5′-GGGACCTGCCTCGTCGTCTA-3′; HBs-F, 5′-AACATGGAGAACATCACATC-3′, HBs-R, 5′-AGCGATAACCAGGACAAGTT-3′; HBe-F, 5′-TCTGTGTTGGGGTGAGT-3′, HBe-R: 5′-TAAGATAGGGGCATTTG-3′. The qPCR results were expressed as relative HBV DNA levels after converting the viral copy numbers to fold changes.
Histological examination
The collected liver tissues were fixed in 10% formaldehyde, dehydrated, and embedded in paraffin. The sections (3 μm) were deparaffinized and stained with hematoxylin and eosin (HE), according to standard protocols. The stained sections were examined under a DM1000 light microscope (Leica, Wetzlar, Germany).
Immunohistochemistry
Paraffin-embedded tissue sections (3 μm) were deparaffinized and rehydrated. After antigen retrieval, the sections were incubated with H2O2 to quench endogenous peroxidase activity and normal goat serum for blocking. The sections were sequentially incubated with anti-TLR4 primary antibody (#PAB33926; 1:100 dilution; Bioswamp) for 12 h at 4°C, and MaxVision™ horseradish peroxidase (HRP)-polymer anti-rabbit secondary antibody (No. KIT-5020; MXB, Fujian, China) for 1 h at room temperature. The sections were developed with 3,3′-diamino benzidine and stained with hematoxylin, according to the manufacturer's instructions. The stained sections were examined under a DM1000 light microscope (Leica).
Flow cytometry
Peripheral blood mononuclear cells (PBMCs) were isolated from mouse sera using a commercial mouse PBMC isolation kit (TBD, Tianjing, China). Subsequently, isolated PBMCs were stained with anti-CD3-FITC (No. 11-0032-82; eBioscience), anti-CD16-APC (No. 17-0161-81; eBioscience), anti-CD14-FITC (No. 11-0141-82; eBioscience), anti-CD11b-PE (No. 12-0112-82; eBioscience), or anti-CD56-PE (No. orb485686; Biorbyt). The frequencies of T cells (CD3+), NK cells (CD3−CD16+CD56+), and mononuclear cells (CD14+CD11b+) were assessed by flow cytometric analysis on an ACEA NovoCyte instrument (Agilent).
Western blotting
Liver tissues were lysed in cold radioimmunoprecipitation assay buffer (No. R0010; Solarbio, Wuhan, China), and the protein concentrations were measured using a bicinchoninic acid assay kit (No. PC0020; Solarbio). Protein samples (20 μg each) were separated on 12% sodium dodecyl sulfate–polyacrylamide gels by electrophoresis and transferred onto nitrocellulose membranes (No. IPVH00010; Millipore, Billerica, MA). The membranes were incubated with 5% nonfat milk in Tris-buffered saline containing Tween 20 for 1 h at room temperature to block nonspecific binding.
The membranes were then sequentially incubated with anti-TLR4 or anti-GAPDH primary antibody (1:1,000 dilution; Bioswamp) for 12 h at 4°C and with HRP-conjugated goat anti-rabbit antibody (No. SAB43714; 1:20,000 dilution; Bioswamp) for 1 h at room temperature. The protein band signals were visualized using an Enhanced Chemiluminescence kit (No. WBKLS0500; Millipore) and a Tanon5200 Imaging System (Tanon, Shanghai, China). The protein level was normalized to that of the internal control GAPDH.
Statistical analysis
Data are expressed as the mean ± standard deviation. Comparisons between groups were performed using analysis of variance with Tukey's post hoc test by GraphPad Prism software (Version 6.0; San Diego, CA). A p-value <0.05 was considered statistically significant.
Results
Expression validation of HBsAg and HBeAg
First, the secreted expression ability of the recombinant plasmids was assessed. As shown in Figure 1, cells transfected with pAAV-1.3HBV secreted high levels of HBsAg and HBeAg, and cells transfected with pAAV-HBs or pAAV-HBe secreted a high level of HBsAg or HBeAg, respectively. These results indicated that recombinant plasmids can express secreted viral proteins.
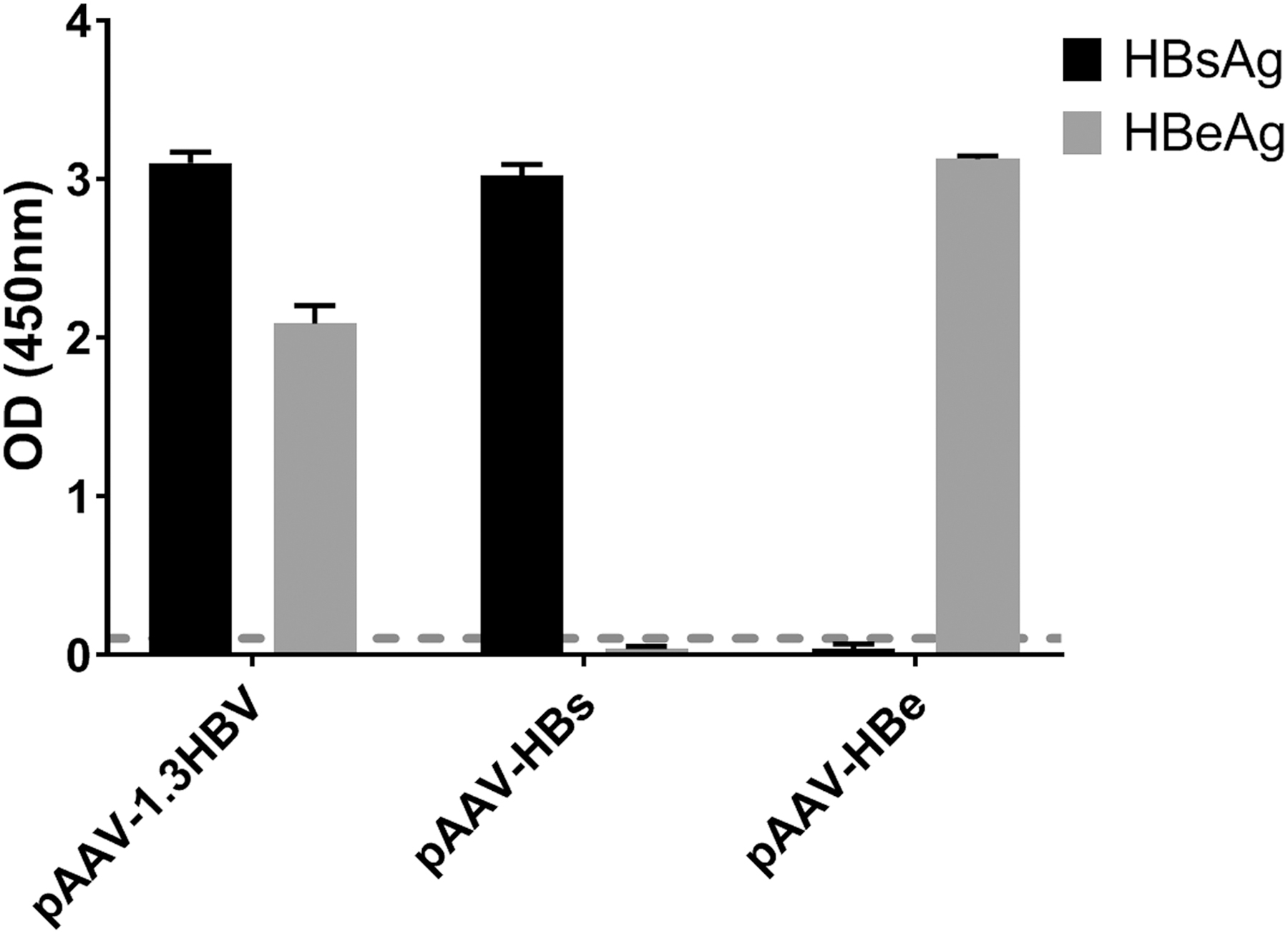
Expression of HBsAg and HBeAg in the supernatant (per mL) of HuH7 cells transfected with the recombinant plasmids pAAV-1.3HBV, pAAV-HBs, and pAAV-HBe, respectively, for 48 h. HBeAg, hepatitis B virus envelope antigen; HBsAg, hepatitis B virus surface antigen.
Establishment of mouse models of chronic HBV infection, persistent HBsAg expression, and persistent HBeAg expression
The serum HBsAg and HBeAg levels are shown in Figure 2A and B. For pAAV-1.3HBV, both HBsAg and HBeAg showed the highest levels at 2 months after the injection, and they gradually decreased. At 6 months after the injection, HBsAg and HBeAg could only be detected in seven mice. For pAAV-HBs, the HBsAg level was the highest at 14 days after the injection and decreased thereafter. At 6 months after the injection, HBsAg could only be detected in three mice. For pAAV-HBe, no HBeAg could be detected after the injection. However, the HBV DNA copy number, HBs gene, and HBe gene could be detected in the liver at 6 months after the injection (Fig. 2C).
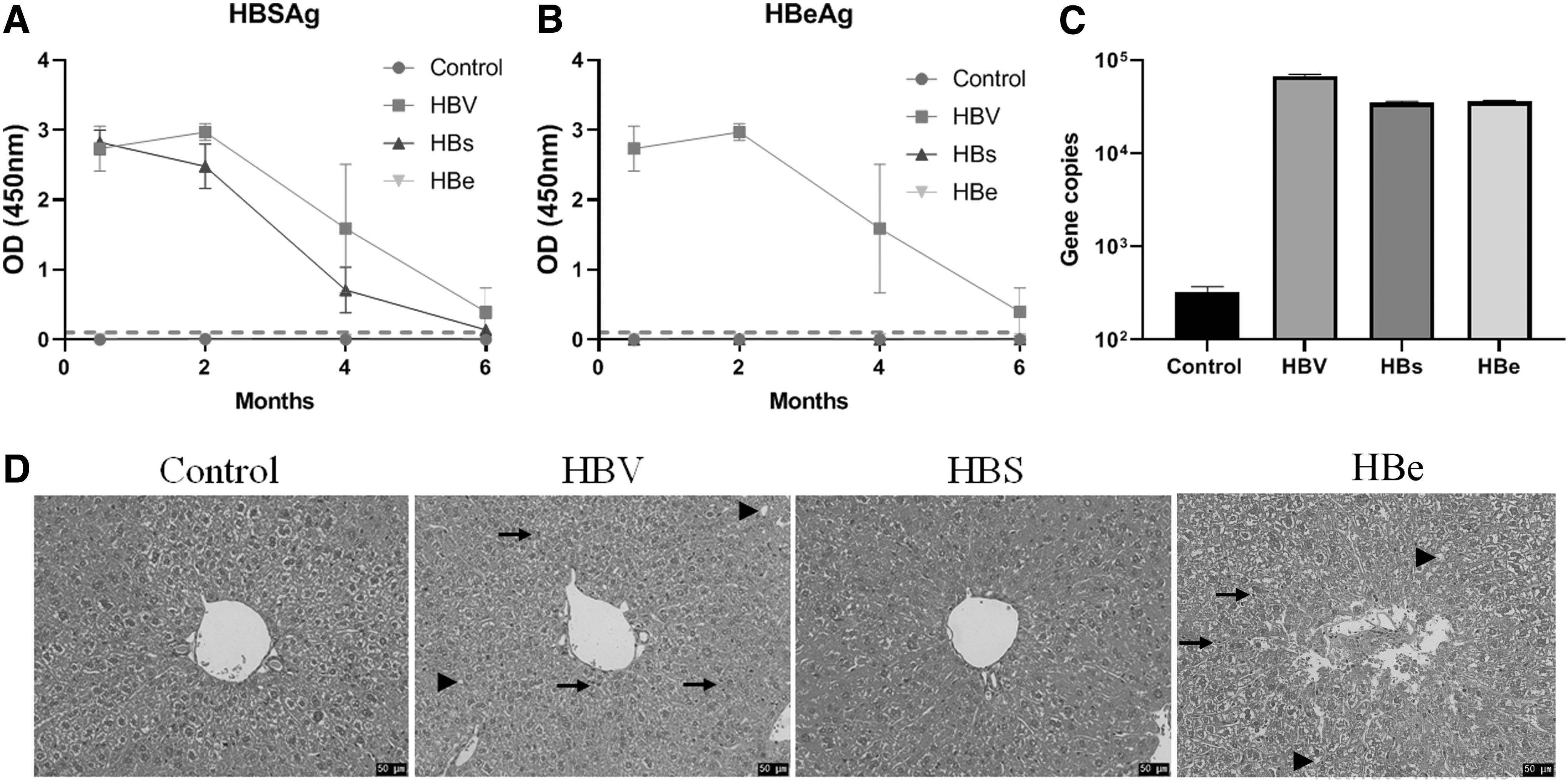
Establishment of persistent HBV infection, persistent HBsAg expression, or persistent HBeAg expression.
HE staining showed that, compared with the mice injected with pAAV-CMV, the mice injected with pAAV-1.3HBV or pAAV-HBe showed obvious inflammatory cell infiltration and fat accumulation, whereas the mice injected with pAAV-HBs showed no changes (Fig. 2D).
Persistent HBsAg or HBeAg expression decreases TLR4 expression
We detected the expression of TLR4 in the liver using both immunohistochemistry and western blot assays. As depicted in Figure 3, both assays showed that HBV infection significantly decreased the expression of TLR4. Moreover, the expression of HBsAg or HBeAg significantly decreased the expression of TLR4, although the change was less than that with HBV infection (p < 0.05).
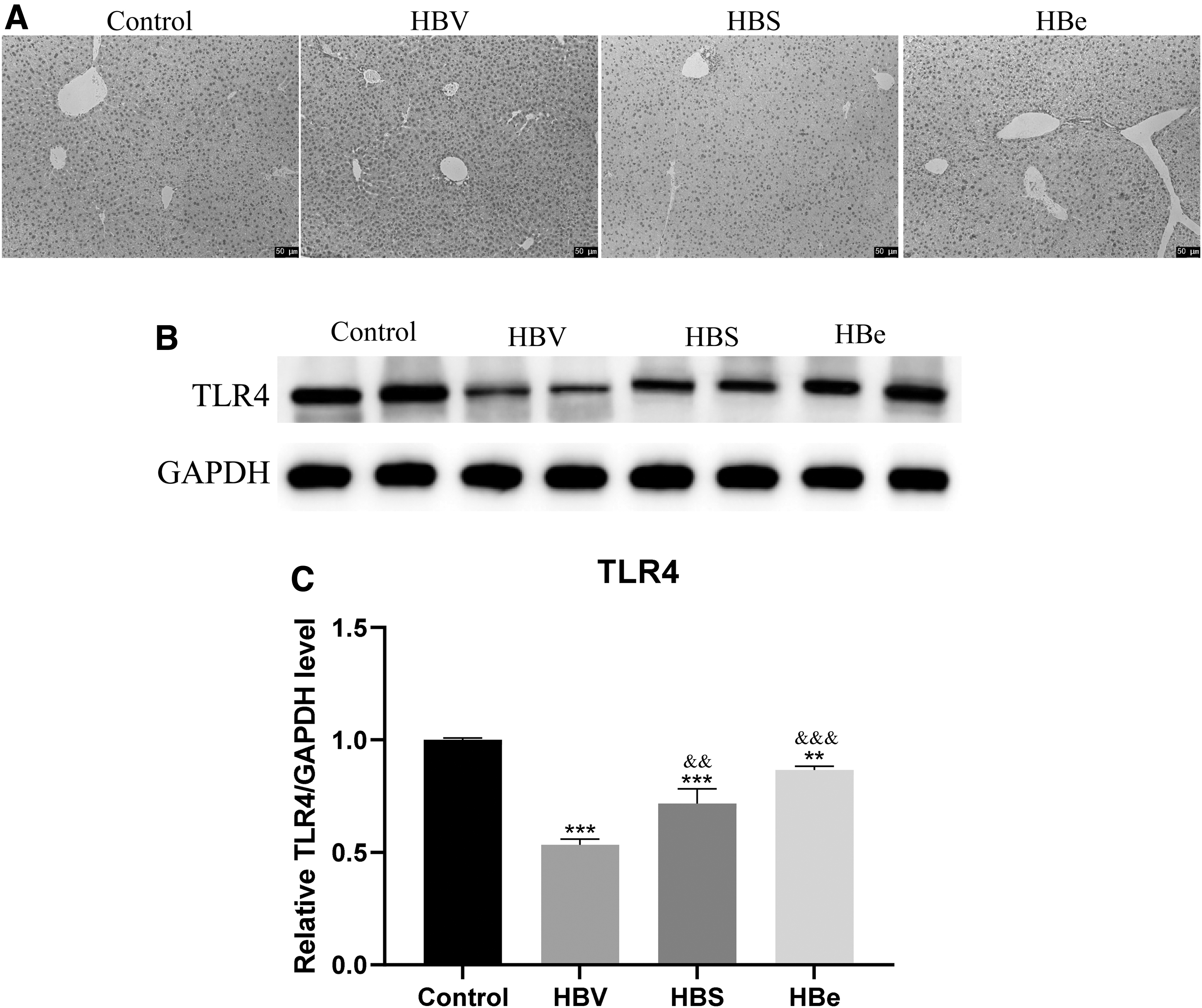
Changes of TLR4 in the three mouse models.
HBsAg and HBeAg contribute to the effect of HBV on proinflammatory cytokines
Compared with the control mice, HBV infection significantly increased the levels of the proinflammatory cytokines IL-1β, IL-6, TNF-α, and TGF-β, whereas it decreased the level of IFN-γ (Fig. 4; all p < 0.001). Moreover, compared with the control mice, proinflammatory cytokines and IFN showed similar changes in the persistent HBsAg expression and persistent HBeAg expression groups (all p < 0.001), showing no significant difference from the HBV infection group, except for IL-6 and IFN-γ in the HBsAg expression group.

Changes of serum proinflammatory cytokines in the three mouse models.
Persistent HBsAg expression and persistent HBeAg expression decrease the percentage of T cells
As shown in Figure 5, chronic HBV infection significantly increased the percentage of T cells. However, both persistent HBsAg expression and persistent HBeAg expression decreased the percentage of T cells, showing an opposite effect than that after HBV infection.
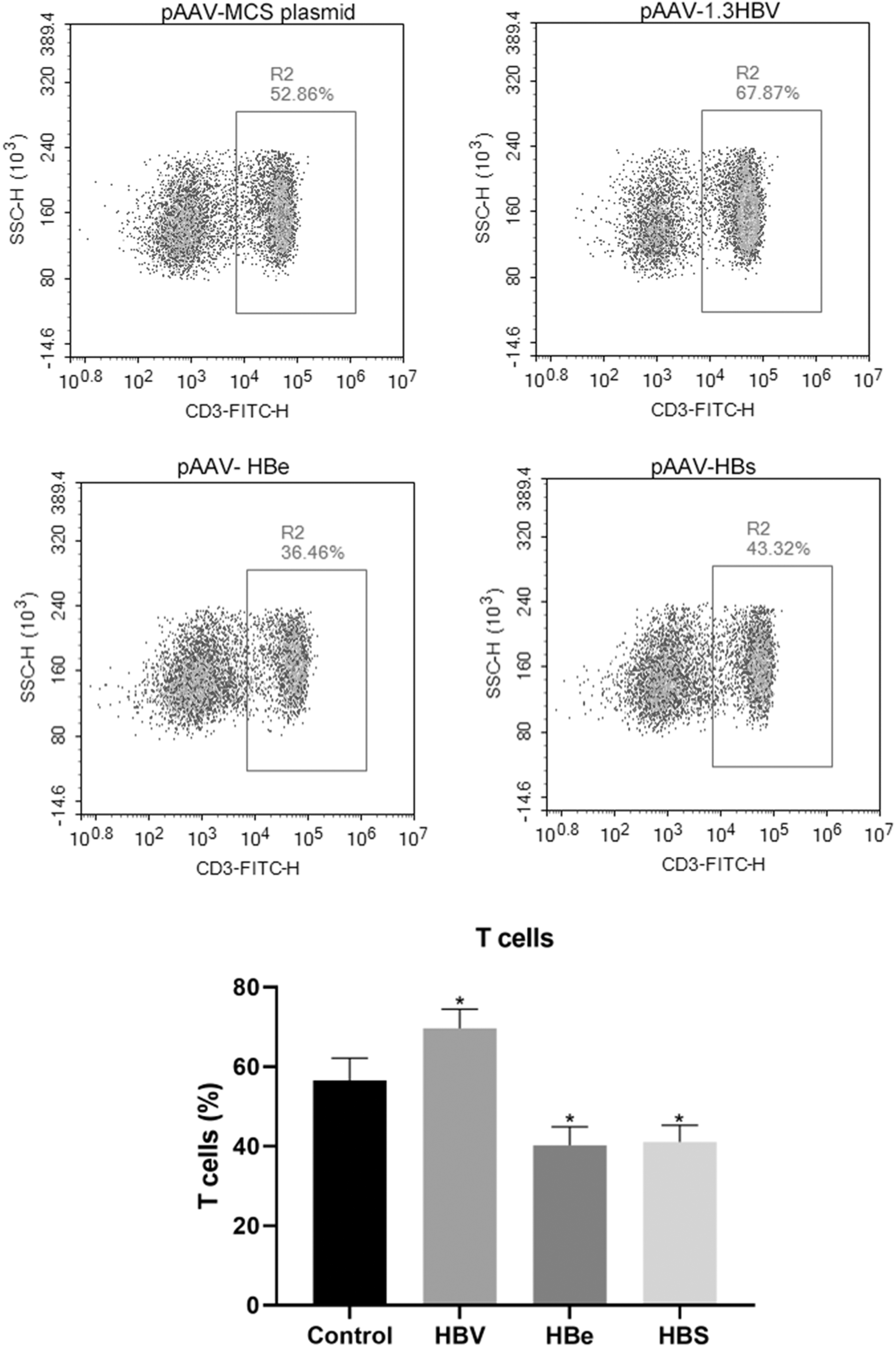
Changes of T cells (CD3+) in the three mouse models. *p < 0.01 compared with the control.
Persistent HBsAg expression increases, whereas persistent HBeAg expression decreases the percentage of NK cells
As shown in Figure 6, chronic HBV infection significantly decreased the percentage of NK cells, and persistent HBeAg expression also decreased the percentage of NK cells. On the contrary, persistent HBsAg expression increased the percentage of NK cells.

Changes of natural killer cells (CD3−CD16+CD56+) in the three mouse models. **p < 0.01 compared with the control, and ***p < 0.001 compared with the control.
Persistent HBsAg expression and persistent HBeAg expression increase the percentage of monocytes
As shown in Figure 7, chronic HBV infection significantly increased the percentage of monocytes. Moreover, both persistent HBsAg expression and persistent HBeAg expression increased the percentage of monocytes, indicating that both HBsAg and HBeAg may be involved in the effect of HBV infection on monocytes.
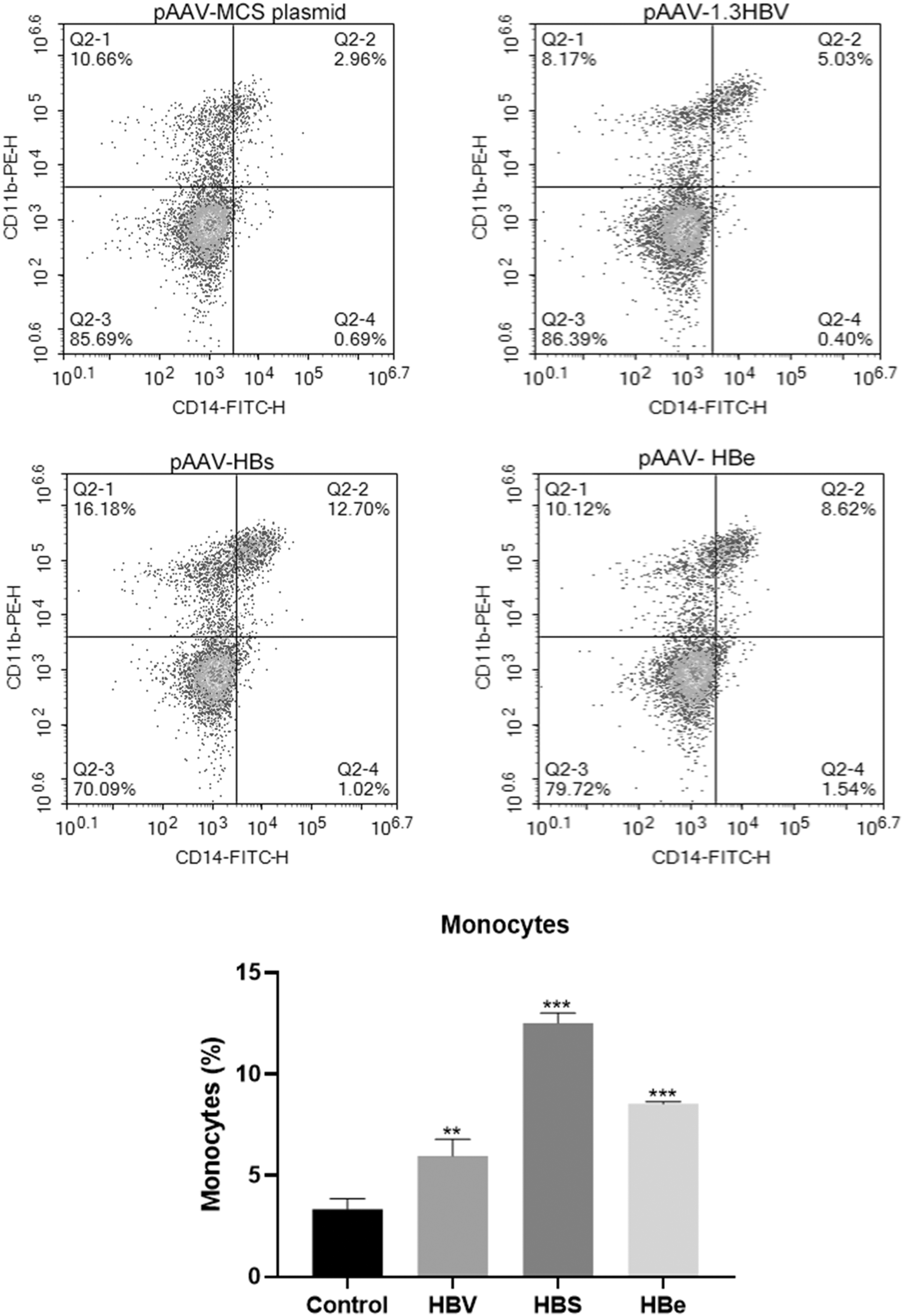
Changes of monocytes (CD14+CD11b+) in the three mouse models. **p < 0.01 compared with the control, and ***p < 0.001 compared with the control.
Discussion
In this study, we established a chronic HBV infection mouse model using high-pressure tail-vein injection of pAAV-1.3HBV. We also developed mouse models of persistent HBsAg or HBeAg expression using the same technique with recombinant pAAV-HBs or pAAV-HBe, respectively. Although secreted HBeAg was not detected, DNA detection and changes of TLR4, cytokines, and immune cells confirmed its expression. We found that chronic HBV infection significantly increased the levels of proinflammatory cytokines, whereas it decreased the level of IFN-γ. Persistent HBsAg expression and persistent HBeAg expression showed similar effects. HBV infection increased the percentages of T cells and monocytes and decreased the percentage of NK cells. However, persistent HBsAg or HBeAg expression showed similar effects only on monocytes but opposite effects on T cells. Persistent HBeAg expression decreased, whereas persistent HBsAg expression increased the percentage of NK cells.
The proinflammatory cytokine IL-1β plays important roles in the control and clearance of HBV. Previous studies have reported that chronic HBV infection adopts a variety of strategies to inhibit both the production and effect of IL-1β. In patients with chronic HBV infection who did not respond to IFN-α treatment, IL-1β production has been shown to be suppressed due to inhibition of the nuclear factor (NF)-κB signaling and inflammasome pathways by HBV (Lei et al., 2019). Moreover, it has been reported that HBeAg significantly inhibits lipopolysaccharide-induced NLRP3 inflammasome activation and IL-1β production through suppressing the NF-κB pathway and reactive oxygen species production (Wang et al., 2019; Yu et al., 2017). In this study, we found that both HBV and its components, HBsAg and HBeAg, increased IL-1β levels. The contrasting results between studies may be due to the different models used, that is, cell models in their studies and a chronic HBV infection mouse model in this study.
IL-6, a cytokine mainly produced by activated monocytes in response to inflammatory stimuli, inhibits HBV entry and transcription. Experiments performed in vitro have showed that the HBV middle S protein enhances IL-6 production through the p38, ERK, and NF-κB pathways (Li et al., 2016). In addition, both IFN-α therapy and nucleos(t)ide analog treatment have been demonstrated to significantly decrease IL-6 levels (Yu et al., 2021; Zhou et al., 2020). Our data on IL-6 were consistent with these previous studies.
During HBV infection, TNF-α acts as a mediator of anti-HBV immunity and induces liver inflammation, but sustained liver inflammation leads to disease progression (Zhong et al., 2021). In addition, TNF-α is associated with ongoing liver damage but not viral clearance in patients with chronic HBV infection (Barathan et al., 2021; Wang et al., 2020). Furthermore, elevated levels of TNF-α have been observed in the serum and hepatocytes of patients with acute or chronic hepatitis B (Wang et al., 2021). We found that the TNF-α level was increased in all treatment groups, indicating increased liver damage. Both HBsAg and HBeAg may contribute to this change. Previous studies performed in vitro also have found that HBeAg can suppress the expression of TNF-α in PBMCs induced by either TLR2 or TLR4 ligands (Zhang and Lu, 2015), whereas TNF-α is reported as a negative regulator during IFN-α production determined by HBsAg (Shi et al., 2012).
TGF-β exerts dual regulatory functions in the immune system in response to HBV infection. TGF-β stimulates the differentiation of Th17 cells, contributing to all stages of liver diseases (Zhong et al., 2021). In this study, both HBV and its components, HBsAg and HBeAg, increased the TGF-β levels, which may cause the differentiation of T cells.
IFNs are the first line of defense that are activated upon viral infection. HBV-specific IFN-γ-producing CD4+ T cells are positively correlated with the decrease of HBsAg and are associated with viral clearance (Wang et al., 2020). In this study, we found that IFN-γ was decreased in all three groups, indicating that HBV as well as HBsAg and HBeAg decreased the antiviral activities of the host.
NK cells are important effectors of innate immunity and respond rapidly to viral infections. In patients with chronic HBV infection, NK cells may be more pathogenic than protective, with preserved cytolytic activity but with a poor capacity to produce antiviral cytokines (Fisicaro et al., 2019). This study revealed that HBV infection suppressed NK cells, whereas it activated T cells and monocytes. A previous study has reported that HBV generates suppressive monocytes, thus initiating the differentiation of regulatory NK cells and resulting in T cell inhibition (Li et al., 2018).
Moreover, HBV infection has been shown to increase the levels of the inhibitory receptor NKG2A on NK cells in mice and humans as well as to reduce their ability to clear HBV, thus facilitating persistent HBV infection (Li et al., 2013). NK cells contribute to hepatic injury and help in viral persistence during chronic hepatitis B progression (Ghosh et al., 2016). Experiments performed in vitro have shown that both HBsAg and HBeAg can directly inhibit NK cell function, thus impairing NK cell cytotoxicity and cytokine production (Yang et al., 2016).
However, our data demonstrated that HBeAg inhibited NK cells, whereas HBsAg increased NK cells. One review has summarized that HBsAg can suppress monocytes, dendritic cells, NK cells, and NK T cells by direct interaction (Kondo et al., 2013). In addition, hepatic NK cells from murine chronic HBsAg carriers show impaired function (Chen et al., 2005). Furthermore, recombinant HBsAg suppresses the activation of monocytes through interaction with a serum protein and a monocyte-specific receptor and also has demonstrated anti-inflammatory properties by inhibiting IL-18 and IL-12 production in a human monocytic cell line. Therefore, HBsAg may regulate NK cells through downregulating the expression of signal transducer and activator of transcription 3 (Zheng et al., 2019).
This study has several limitations that must be addressed. First, the expression of HBsAg and HBeAg in cells was not validated due to the limitation of available antibodies. Second, although HBsAg and HBeAg were found to contribute to the innate immune response during persistent HBV infection, further experiments are needed to uncover their functions due to the complex innate immune system.
Conclusions
In conclusion, we found that persistent HBV infection as well as persistent HBsAg expression and persistent HBeAg expression significantly increased the expression of TLR4 and proinflammatory cytokines as well as the percentage of monocytes, whereas they decreased the level of IFN-γ. HBV infection increased the percentage of T cells and decreased the percentage of NK cells, whereas only persistent HBeAg expression contributed to a decreased percentage of NK cells, indicating that although both HBsAg and HBeAg contribute to persistent HBV infection, they may function differently.
Footnotes
Authors' Contributions
Conceptualization, project administration, supervision, and resources by D.-W.Z. Writing original draft and formal analysis by J.-M.Z. Resources, formal analysis, and writing review and editing by N.-L.K. Investigation and formal analysis by L.-Y.W.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Fujian Clinical Research Center for Hepatopathy and Intestinal Diseases (No. 2021Y2006) and the Science and Technology Department of Fujian (No. 2018J01164).