
Abstract
Interleukin-36 (IL-36) signaling plays an important role in promoting CD8+ T cell-mediated antitumor immune responses. The role of IL-36 signaling in CD8+ T cells that are involved in host immune responses during human immunodeficiency virus-1 (HIV-1) infection has not been characterized. Sixty-one patients living with chronic HIV-1 infection and 23 controls were enrolled in this study. The levels of IL-36 cytokine family members were measured by enzyme-linked immunosorbent assay. Purified CD8+ T cells were stimulated with recombinant IL-36gamma (1 or 10 ng/mL). The expression of inhibitory receptors, the secretion of cytotoxic molecules and interferon-gamma, and the mRNA levels of apoptosis-related ligands were assessed to evaluate the effect of IL-36gamma on CD8+ T cell function in vitro. There were no significant differences in IL-36alpha, IL-36beta, or IL-36 receptor antagonist levels between patients living with chronic HIV-1 infection and controls. Plasma IL-36gamma levels were reduced in patients living with chronic HIV-1 infection. Perforin, granzyme B, and granulysin secretion, as well as tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and Fas ligand (FasL) mRNA expression, but not programmed death-1 (PD-1) or cytotoxic T lymphocyte-associated protein-4 (CTLA-4) expression was downregulated in CD8+ T cells from patients living with chronic HIV-1 infection. The addition of both 1 and 10 ng/mL IL-36gamma enhanced perforin, granzyme B, granulysin, and interferon-gamma secretion by CD8+ T cells without affecting PD-1/CTLA-4 or TRAIL/FasL mRNA expression in CD8+ T cells from patients living with chronic HIV-1 infection. The addition of 1 ng/mL IL-36gamma also promoted perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. The reduced IL-36gamma levels in patients living with chronic HIV-1 infection might be insufficient for the activation of CD8+ T cells, leading to CD8+ T cell exhaustion.
Introduction
CD8
Thus, CD8+ T cell reprogramming or restoration, combined with the administration of other available immunomodulators, may promote the effect of adoptive cell therapy to potentially cure HIV-1 infection (Perdomo-Celis et al., 2022).
Interleukin-36 (IL-36) cytokines belong to the IL-1 superfamily and include four members, including three agonists (IL-36alpha, IL-36beta, and IL-36gamma) and one receptor antagonist (IL-36 receptor antagonist, IL-36Ra) (Elias et al., 2021). All IL-36 cytokines bind to the IL-36 receptor complex (IL-36R), which is composed of IL-1 receptor 6 (IL-1R6) and IL-1 receptor accessory protein (IL-1RacP) (Neurath, 2020).
IL-36alpha, IL-36beta, and IL-36gamma functionally act as IL-36R agonists. Signaling initiated by the binding of IL-36alpha, IL-36beta, and IL-36gamma to IL-36R leads to the recruitment of IL-1RacP and induces inflammatory responses through the activation and regulation of several inflammatory pathways (Elias et al., 2021; Ngo et al., 2021). Conversely, IL-36Ra binding fails to recruit IL-1RacP and cannot initiate signaling responses (Elias et al., 2021; Ngo et al., 2021). Thus, IL-36 cytokines play dual roles, potentially mediating inflammatory responses in cancers and inflammation and protecting epithelial barrier tissues from infection (Han et al., 2020). Furthermore, IL-36 cytokines can also regulate immune cell functions. IL-36 cytokines promote the infiltration and activation of myeloid cells in the skin (Foster et al., 2014).
IL-36beta strongly promotes CD8+ T cell activation, contributing to antitumor immune responses by activating mammalian target of rapamycin complex 1 (mTOC1) (Zhao et al., 2019). IL-36gamma also functions synergistically with T cell receptor signaling and/or IL-12 to stimulate CD8+ T cells, natural killer cells, and γδT cells, resulting in the transformation of the tumor microenvironment in favor of tumor eradication (Wang et al., 2015).
Chronic hepatitis B virus (HBV) infection induces increased production of IL-36alpha, which correlates with HBV viral load (Gong et al., 2017). However, few studies have focused on the expression profile of IL-36 cytokines in patients living with chronic HIV-1 infection. Due to the ability of IL-36 to modulate CD8+ T cells, we hypothesized that IL-36 cytokines also contribute to the regulation of CD8+ T cell function during chronic HIV-1 infection. To test this hypothesis, we first analyzed circulating IL-36 levels in patients living with HIV-1 infection. The regulatory effects of recombinant human IL-36 cytokines on purified CD8+ T cells from patients living with HIV-1 infection were then assessed in vitro.
Materials and Methods
Study subjects
Sixty-one patients living with chronic HIV-1 infection (47 males and 14 females; median age, 41 years; range, 21–57 years) and 23 healthy controls (19 males and 4 females; median age, 39 years; range, 26–59 years) were enrolled in the study at Lanzhou Pulmonary Hospital. All the patients who were living with chronic HIV-1 infection were treatment naive. The laboratory values for these patients were as follows: CD4+ T cell count, 116 (275–901)/μL; and HIV-1 RNA load, 391,104 (58,346–3,099,000) copies/mL. Patients who had acute or chronic disease (e.g., viral hepatitis, autoimmune disorders, or cancers) or were using any drugs/alcohol were excluded from the study. Informed consent was obtained from each participant according to the guidelines for conducting of clinical research. The study protocol was approved by Lanzhou Pulmonary Hospital Institutional Review Boards (no. 2020-198-07[K]).
Cell purification
Peripheral blood specimens were collected from all the study individuals and were centrifuged to separate the plasma. Peripheral blood mononuclear cells (PBMCs) were separated from each whole blood sample by Ficoll-Hypaque (Solarbio, Beijing, China) density gradient centrifugation. CD8+ T cells were negatively selected with a MagCellect Human CD8+ T Cell Isolation Kit (R&D Systems, Minneapolis, MN). The purity of CD8+ T cells was analyzed by staining with fluorochrome-conjugated antibodies against CD3, CD14, and CD19 and subsequent flow cytometry analysis. The CD8+ T cell purity was >95%, the cells that used for all experiments were free of detectable CD14+ monocytes or CD19+ B cells. PBMCs or CD8+ T cells were cultured in complete Roswell Park Memorial Institute (RPMI) 1640 supplemented with 10% fetal calf serum (FCS) and antibodies against CD3/CD28 (100 ng/mL) in the presence or absence of recombinant human IL-36gamma (aa18-169) protein (final concentration, 1 or 10 ng/mL; R&D Systems) for 48 h.
In certain experiments for enzyme-linked immunospot (ELISPOT) assay, PBMCs from 12 HLA-A*02-restricted patients living with chronic HIV-1 infection were stimulated with HLA-A*02-restricted gag p17 epitope (SLYNTVATL) (Vali et al., 2011) (final concentration, 10 μg/mL) and HLA-A*02-restricted gag p24 epitope (TLNAWVKVV) (Schaubert et al., 2007) (final concentration, 10 μg/mL) in the presence or absence of recombinant human IL-36gamma (aa18-169) protein (final concentration, 1 ng/mL) for 24 h.
Flow cytometry
PBMCs were stained with anti-CD3-peridinin-chlorophyll-protein complex, anti-CD8-allophycocyanin, anti-programmed death-1 (PD-1)-fluorescein isothiocyanate (FITC), and anti-cytotoxic T lymphocyte-associated protein-4 (CTLA-4)-P-phycoerythrin (PE) (BD Pharmagen, San Jose, CA) antibodies. Isotype controls for the FITC- and PE-conjugated antibodies were also used for the separation of positive and negative cells. For analysis, cell data were acquired on a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA), and fluorescence activating cell sorter data were analyzed using FlowJo software.
Enzyme-linked immunosorbent assay
The levels of IL-36alpha, IL-36beta, IL-36gamma, and IL-36Ra in plasma samples as well as the levels of perforin, granzyme B, granulysin, and interferon-gamma in supernatants were measured using the Enzyme-Linked Immunosorbent Assay (ELISA) Kits. The IL-36alpha, IL-36beta, IL-36gamma, and IL-36Ra ELISA Kits were purchased from R&D Systems. The Perforin, granzyme B, granulysin, and interferon-gamma ELISA Kits were purchased from Neobioscience (Shenzhen, Guangdong Province, China).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from CD8+ T cells using TRIzol reagent (Invitrogen, Carlsbad, CA) and reverse transcribed into cDNA using the PrimeScript RT Reagent Kit (TaKaRa, Beijing, China). Real-time polymerase chain reaction (PCR) was then performed using TB Green Premix Taq (TaKaRa). The primers for IL-1R6 and IL-1RacP were purchased from Applied Biosystems (Foster, CA). The primer sequences for tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) were as follows: forward: 5′-CTG CTG GCA AGT CAA GTG GCA ACT C-3′, and reverse: 5′-GTC GCA TCC TGA AAA CTG AAT AGT-3′; those for Fas ligand (FasL) were as follows: forward: 5′-ATG TTT CAG CTC TTC CAC CTA CAG AAG GA-3′, and reverse: 5′-CAG AGA GAG CTC AGA TAC GTT GAC-3′. All the samples were analyzed in triplicate, and the relative expression of target genes was determined by normalizing to the expression of glyceraldehyde 3-phosphate dehydrogenase to calculate a fold change in expression. Gene expression was calculated using the comparative delta-delta-Ct method.
ELISPOT assay
The secretion of perforin and granzyme B by HIV-1-specific CD8+ T cells were measured using the Human Perforin ELISPOT Kit (Abcam, Cambridge, MA) and the Human Granzyme B ELISPOT Kit (Abcam), respectively. The results were shown as numbers of spot-forming cells.
Statistical analysis
Data were analyzed using SPSS 23.0 software. Student's t tests, one-way analysis of variance (ANOVA), or least significant difference (LSD)-t tests were employed to assess the statistical significance of differences in data that followed a normal distribution, and these data are presented as the mean ± standard deviation. Mann–Whitney tests or Kruskal–Wallis tests were employed to assess the statistical significance of differences in data that followed a skewed distribution, and these data are presented as the median (Q1–Q3). Relations between variables were tested using Spearman rank correlation test. All the tests were two-tailed, and p-values of <0.05 were considered significant.
Results
Plasma IL-36gamma levels were decreased in patients living with chronic HIV-1 infection
The plasma levels of IL-36 cytokine family members in all study participants were measured. There were no significant differences in IL-36alpha (73.60 [47.84–129.7] pg/mL vs. 88.32 [53.36–193.2] pg/mL; Mann–Whitney test, p = 0.236, Fig. 1A), IL-36beta (87.00 [53.94–121.8] pg/mL vs. 71.34 [44.27–107.9] pg/mL; Mann–Whitney test, p = 0.266, Fig. 1B) or IL-36Ra (44.46 [31.98–84.99] pg/mL vs. 57.72 [34.32–88.92] pg/mL; Mann–Whitney test, p = 0.574, Fig. 1D) between patients living with chronic HIV-1 infection and healthy controls. Plasma IL-36gamma levels were significantly decreased in patients living with chronic HIV-1 infection compared with healthy controls (589.3 [365.3–862.0] pg/mL vs. 1,047 [603.9–1,763] pg/mL; Mann–Whitney test, p = 0.0017, Fig. 1C). Plasma IL-36gamma levels were negatively correlated with CD8+ T cell counts in patients living with chronic HIV-1 infection (Spearman rank correlation test, r = −0.293, p = 0.022, Fig. 1F), but there was no remarkable correlation between IL-36gamma levels and CD4+ T cell counts (Spearman rank correlation test, r = 0.100, p = 0.443, Fig. 1E).

Expression of IL-36 cytokine family members and the correlation with clinical indices in patients living with chronic HIV-1 infection. Plasma levels of IL-36 cytokine family members in patients living with chronic HIV-1 infection (n = 61) and controls (n = 23) were measured by ELISA.
Plasma IL-36gamma levels did not correlate with HIV RNA levels in patients living with chronic HIV-1 infection (Spearman rank correlation test, r = 0.031, p = 0.811, Fig. 1G).
Cytotoxic molecule secretion and TRAIL and FasL expression, but not PD-1 or CTLA-4 expression, by CD8+ T cells were reduced in patients living with chronic HIV-1 infection
PBMCs from all study participants were stained with antibodies against CD3, CD8, PD-1, and CTLA-4. The representative flow dot plots showing PD-1 and CTLA-4 expression in the CD3+CD8+ T cells from a patient living with chronic HIV-1 infection and those from a healthy control are presented in Figure 2A and B, respectively. There were no significant differences in either the PD-1+CD8+ T cell proportion (6.99 ± 1.58% vs. 7.40 ± 1.71%; Student's t test, p = 0.304, Fig. 2A) or CTLA-4+CD8+ T cell proportion (16.80 ± 6.29% vs. 16.31 ± 3.63%; Student's t test, p = 0.726, Fig. 2B) among CD8+ T cells from patients living with chronic HIV-1 infection and healthy controls.

PD-1 and CTLA-4 expression in CD8+ T cells from patients living with chronic HIV-1 infection. PBMCs were isolated from patients living with chronic HIV-1 infection (n = 61) and controls (n = 23). A total of 5 × 106 PBMCs were stained with anti-CD3-PerCP, anti-CD8-APC, anti-PD-1-FITC, and anti-CTLA-4-PE antibodies. Isotype controls for the FITC- and PE-conjugated antibodies were also used for the separation of positive and negative cells.
A total of 2 × 105 CD8+ T cells, which were purified from 33 patients living with chronic HIV-1 infection and 14 healthy controls, were cultured in RPMI 1640 supplemented with FCS and anti-CD3/anti-CD28 antibodies for 48 h. Supernatants were harvested for ELISA, and cells were harvested for real-time PCR analysis. The levels of perforin (299.6 ± 58.03 pg/mL vs. 351.7 ± 50.30 pg/mL; Student's t test, p = 0.0054, Fig. 3A), granzyme B (157.6 ± 33.60 pg/mL vs. 204.8 ± 57.11 pg/mL; Student's t test, p = 0.0009, Fig. 3B), and granulysin (53.49 ± 11.26 pg/mL vs. 71.66 ± 17.96 pg/mL; Student's t test, p = 0.0001, Fig. 3C) in the culture supernatants of CD8+ T cells from patients living with chronic HIV-1 infection were significantly reduced compared with those of CD8+ T cells from controls. However, there was no marked difference in interferon-gamma secretion by CD8+ T cells from patients living with chronic HIV-1 infection or those from controls (14.82 ± 2.06 pg/mL vs. 14.87 ± 1.60 pg/mL; Student's t test, p = 0.933, Fig. 3D).
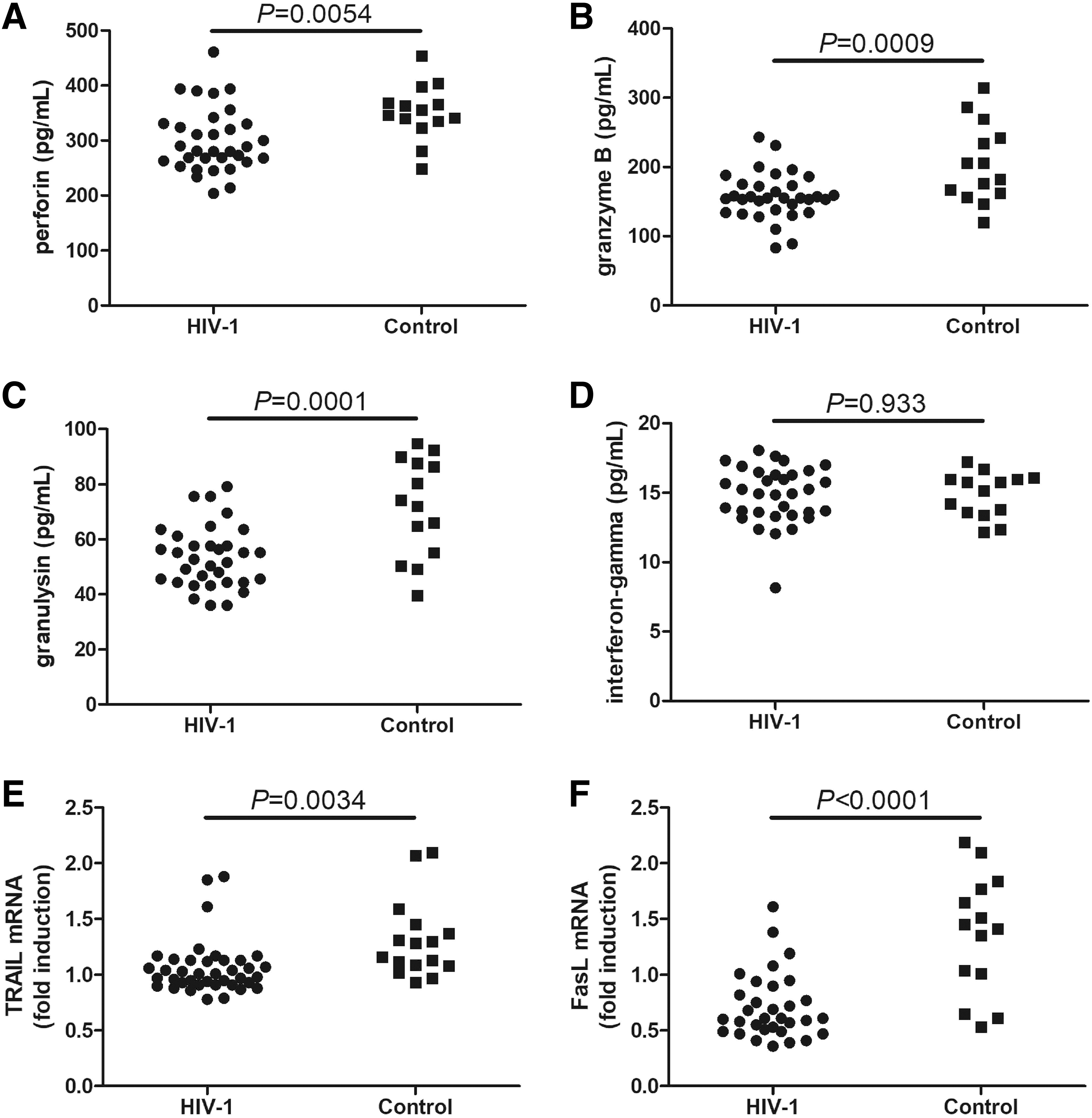
Cytotoxic molecule and cytokine secretion and TRAIL and FasL mRNA expression by CD8+ T cells from patients living with chronic HIV-1 infection. CD8+ T cells were purified from patients living with chronic HIV-1 infection (n = 33) and controls (n = 14). A total of 2 × 105 CD8+ T cells were cultured in RPMI 1640 supplemented with FCS and anti-CD3/anti-CD28 antibodies for 48 h. The expression levels of perforin, granzyme B, granulysin, and interferon-gamma in the supernatants were measured by ELISA, while the mRNA expression levels of TRAIL and FasL in CD8+ T cells were semiquantified by real-time PCR.
The mRNA expression levels of TRAIL (1.06 ± 0.24 vs. 1.31 ± 0.35; Student's t test, p = 0.0034, Fig. 3E) and FasL (0.71 ± 0.29 vs. 1.37 ± 0.54; Student's t test, p < 0.0001, Fig. 3F) were also strongly downregulated in CD8+ T cells from patients living with chronic HIV-1 infection compared with those from controls.
Exogenous IL-36gamma enhanced cytotoxic molecule production without affecting PD-1/CTLA-4 or TRAIL/FasL mRNA expression in CD8+ T cells from patients living with chronic HIV-1 infection
To investigate the influence of IL-36gamma on CD8+ T cells, we first investigated IL-36 receptor mRNA expression in CD8+ T cells. There were no significant differences in either the IL-1R6 mRNA (0.93 [0.65–1.66] vs. 0.92 [0.71–1.49]; Mann–Whitney test, p = 0.863, Fig. 4A) or IL-1RacP mRNA (0.78 [0.52–1.39] vs. 1.01 [0.61–1.74]; Mann–Whitney test, p = 0.316, Fig. 4B) levels in CD8+ T cells from patients living with chronic HIV-1 infection and those from controls.
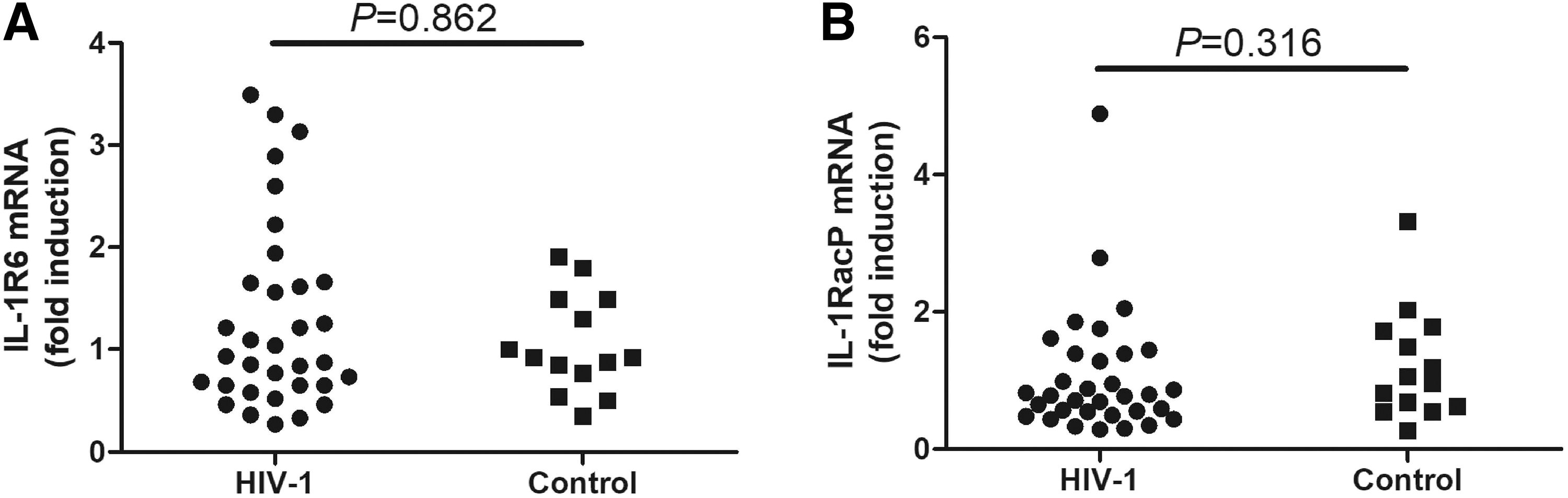
IL-1R6 and IL-1RacP mRNA expression in CD8+ T cells from patients living with chronic HIV-1 infection. CD8+ T cells were purified from patients living with chronic HIV-1 infection (n = 33) and controls (n = 14). The mRNA expression of IL-1R6 and IL-1RacP in CD8+ T cells was semiquantified by real-time PCR.
A total of 5 × 105 PBMCs from 28 patients living with chronic HIV-1 infection and 12 controls were stimulated with 1 or 10 ng/mL recombinant human IL-36gamma for 48 h. PD-1 and CTLA-4 expression in CD3+CD8+ T cells was analyzed by flow cytometry. The representative flow dot plots in Figure 5A and B show PD-1 and CTLA-4 expression in CD3+CD8+ T cells from a patient living with chronic HIV-1 infection and a healthy control, respectively, after being exposed to no stimulation, 1 ng/mL IL-36gamma, or 10 ng/mL IL-36gamma stimulation. Neither 1 nor 10 ng/mL of IL-36gamma stimulation affected the PD-1+CD8+ T cell proportion (one-way ANOVA, p > 0.05, Fig. 5C) or CTLA-4+CD8+ T cell proportion (Kruskal–Wallis tests, p > 0.05, Fig. 5D) among CD8+ T cells from patients living with chronic HIV-1 infection or controls.
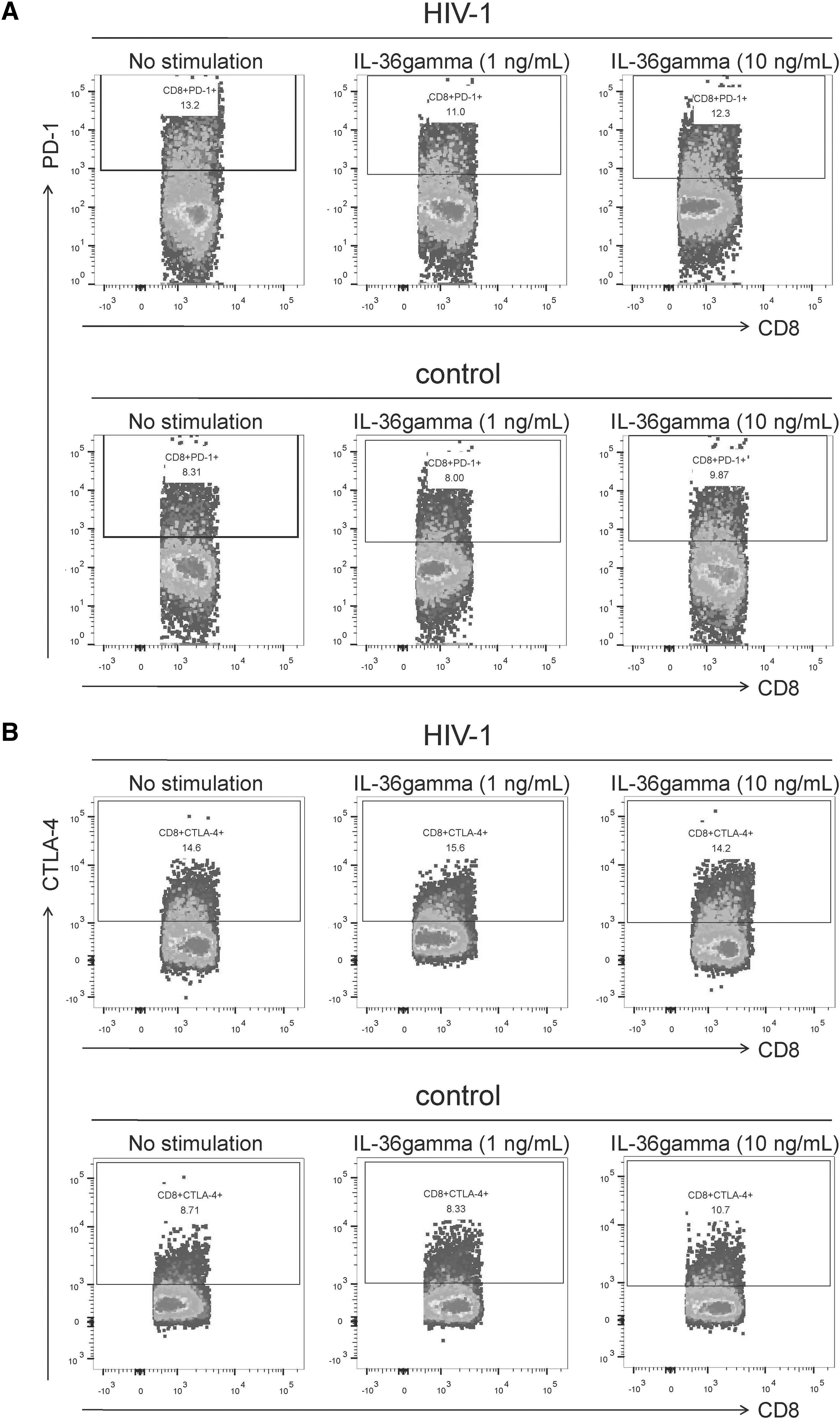
Effect of exogenous IL-36gamma on PD-1 and CTLA-4 expression in CD8+ T cells from patients living with chronic HIV-1 infection and controls. A total of 5 × 105 PBMCs from patients living with chronic HIV-1 infection (n = 28) and controls (n = 12) were stimulated with 1 or 10 ng/mL recombinant human IL-36gamma for 48 h. Cells were harvested and were stained with anti-CD3-PerCP, anti-CD8-APC, anti-PD-1-FITC, and anti-CTLA-4-PE antibodies. The expression of PD-1 and CTLA-4 in CD8+ T cells was assessed by flow cytometry.

A total of 2 × 105 CD8+ T cells, which were purified from 22 patients living with chronic HIV-1 infection and 10 healthy controls, were stimulated with 1 or 10 ng/mL recombinant human IL-36gamma for 48 h. Stimulation with either 1 or 10 ng/mL IL-36gamma enhanced perforin (LSD-t test, p < 0.01, Fig. 6A), granzyme B (LSD-t test, p < 0.05, Fig. 6B), and granulysin (LSD-t test, p < 0.05, Fig. 6C) secretion by CD8+ T cells from patients living with chronic HIV-1 infection. However, only 10 ng/mL IL-36gamma, but not 1 ng/mL IL-36gamma, promoted perforin, granzyme B, and granulysin production by CD8+ T cells from controls (LSD-t tests, p < 0.05, Fig. 6A–C). Stimulation with IL-36gamma at either 1 or 10 ng/mL elevated interferon-gamma secretion by CD8+ T cells from patients living with chronic HIV-1 infection and controls (LSD-t test, p < 0.01, Fig. 6D). However, stimulation with neither 1 nor 10 ng/mL of IL-36gamma affected TRAIL mRNA or FasL mRNA expression in CD8+ T cells from patients living with chronic HIV-1 infection or controls (one-way ANOVA, p > 0.05, Fig. 6E, F).

Effect of exogenous IL-36gamma on cytotoxic molecules and cytokine secretion and TRAIL and FasL mRNA expression in CD8+ T cells from patients living with chronic HIV-1 infection and controls. CD8+ T cells were purified from patients living with chronic HIV-1 infection (n = 22) and controls (n = 10). A total of 2 × 105 of CD8+ T cells were stimulated with 1 or 10 ng/mL recombinant human IL-36gamma for 48 h. The expression levels of perforin, granzyme B, granulysin, and interferon-gamma in the supernatants were measured by ELISA, while the mRNA expression levels of TRAIL and FasL in CD8+ T cells were semiquantified by real-time PCR.
A total of 106 PBMCs from 12 HLA-A*02-restricted patients living with chronic HIV-1 infection were stimulated with HLA-A*02-restricted gag peptides and 1 ng/mL of recombinant human IL-36gamma for 24 h. One nanogram per microliter of IL-36gamma stimulated promoted both perforin and granzyme B secretion by HIV-1-specific CD8+ T cells (Student's t tests, p < 0.01, Fig. 7A, B).
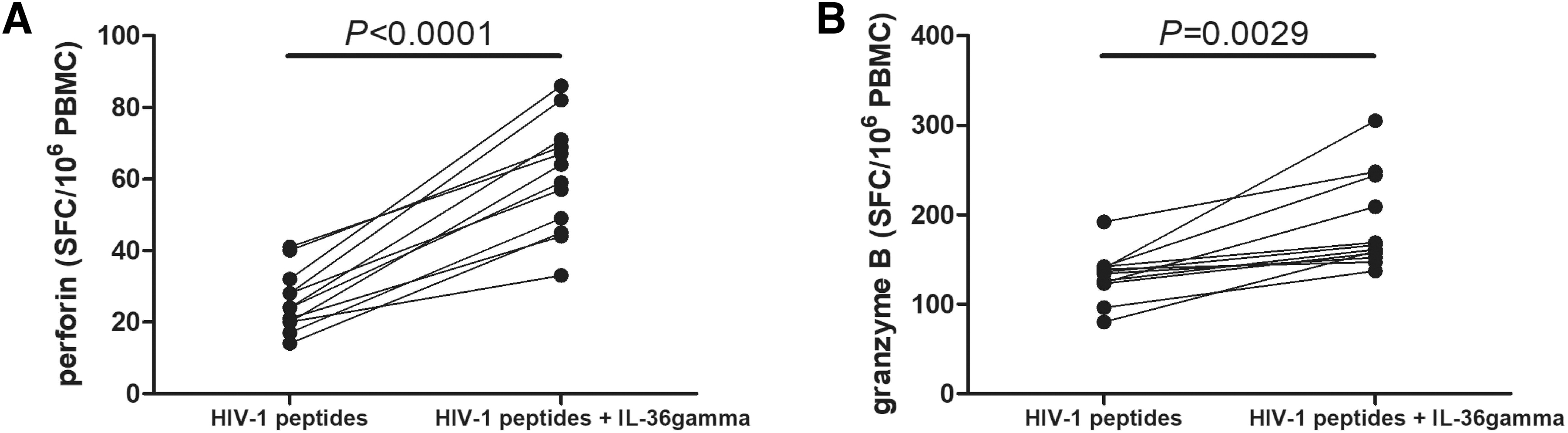
Effect of exogenous IL-36gamma to perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. A total of 106 PBMCs from 12 HLA-A*02-restricted patients living with chronic HIV-1 infection were stimulated with HLA-A*02-restricted gag p17 epitope (SLYNTVATL) (final concentration, 10 μg/mL) and HLA-A*02-restricted gag p24 epitope (TLNAWVKVV) (final concentration, 10 μg/mL) in the presence or absence of 1 ng/mL of recombinant human IL-36gamma for 24 h. Perforin and granzyme B secretion was measured by ELISPOT.
Discussion
In this study, we first screened IL-36 cytokine family members in the circulation of in patients living with chronic HIV-1 infection. We found that there were no significant differences in peripheral levels of IL-36alpha, IL-36beta, or IL-36Ra between patients living with chronic HIV-1 infection and controls. Importantly, IL-36gamma levels were strongly decreased and negatively correlated with CD8+ T cell counts in patients living with chronic HIV-1 infection.
Furthermore, CD8+ T cells exhibited a dysfunctional phenotype in patients living with HIV-1 infection, exhibiting reduced secretion of cytotoxic molecules (including perforin, granzyme B, and granulysin) and downregulation of the mRNA levels of apoptosis-related ligands (TRAIL and FasL). Stimulation of either a lower concentration (1 ng/mL) or a higher concentration (10 ng/mL) of recombinant IL-36gamma in vitro strongly enhanced cytotoxic molecule and interferon-gamma secretion by CD8+ T cells purified from patients living with chronic HIV-1 infection, but only a higher concentration of recombinant IL-36gamma promoted cytotoxic molecule expression by CD8+ T cells purified from controls.
More importantly, 1 ng/mL IL-36gamma also promoted perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. This indicated that CD8+ T cells from patients living with chronic HIV-1 infection might be more sensitive to IL-36gamma stimulation. The immunomodulatory properties of IL-36gamma might contribute to restoring the dysfunctional or exhausted CD8+ T cell phenotype during the treatment of chronic HIV-1 infection.
Similar to the potential dual functions of IL-36 cytokines in inflammation (Han et al., 2020), controversy remains regarding IL-36 cytokine expression and activity during viral infection. Influenza virus infection induced high expression of IL-36gamma in both PBMCs and human lung epithelial cells. The elevation of IL-36gamma enhanced type I and III interferons, promoted apoptosis, and suppressed autophagy in the early stages of influenza infection in vitro (Liu et al., 2020)). Moreover, IL-36R-deficient mice were protected from influenza virus-induced lung injury and mortality (Aoyagi et al., 2017; Koss et al., 2021). Decreased mortality was closely related to reduced early accumulation of neutrophils and monocytes/marcophages, activation of lymphocytes, secretion of proinflammatory cytokines/chemokines, and permeability of the alveolar–epithelial barrier (Aoyagi et al., 2017). IL-36 agonists might exacerbate lung injury during influenza virus infection. Herpes simplex virus-1 (HSV-1) caused increases in IL-36alpha and IL-36beta mRNA expression in infected skin.
IL-36beta-deficient mice succumbed more frequently to HSV-1 infection and developed larger zosteriform skin lesions along infected neurons (Milora et al., 2017). Similarly, HSV-2 infection induced increases in IL-36gamma. In vivo treatment of mice with IL-36gamma not only suppressed HSV-2 replication but also drove the transient production of immune mediators and promoted polymorphonuclear leukocyte infiltration into the vaginal microenvironment, leading to resistance to HSV-2 infection and genital disease (Gardner and Herbst-Kralovetz, 2018). These results indicated that IL-36 cytokines provided protection against HSV-1 and HSV-2 infection. A designed truncated IL-36gamma-encoding plasmid acted as a potent molecular adjuvant that enhanced several DNA-encoded vaccine targets, including HIV-1, influenza virus, and Zika virus in immunization models (Louis et al., 2019).
To the best of our knowledge, this was the first report on the IL-36 cytokine profile during HIV-1 infection. Our current data revealed that chronic HIV-1 infection suppressed only the plasma IL-36gamma levels but did not affect the IL-36alpha, IL-36beta, or IL-36Ra levels.
Reduced circulating IL-36gamma levels were negatively correlated with CD8+ T cell counts in patients living with HIV-1 infection but not with either CD4+ T cell counts or HIV-1 viral loads. This indicated that IL-36gamma might be associated with the regulation of CD8+ T cells in patients with chronic HIV-1 infection. The predominant role of IL-36gamma during HIV-1 infection needs further elucidation.
A hallmark of chronic viral infections is the presence of dysfunctional or exhausted CD8+ T cells, which are characterized by impaired effector function, distinct transcriptional programs, and elevated expression of inhibitory receptors and apoptosis-related ligands (Sandu et al., 2020). Chronic hepatitis E virus (HEV) infection was associated with HEV-specific CD8+ T cell exhaustion, which presented as diminished HEV-specific CD8+ T cell counts and exhausted phenotypic features (Kemming et al., 2022). CD8+ T cells were present in fewer numbers in the full-term placenta of HBV-infected pregnant women and exhibited decreased expression of the activation molecules CD69 and HLA-DR. Thus, CD8+ T cells in term pregnancy deciduas with chronic HBV infection demonstrated impaired functional signatures through the reduced production of interferon-gamma and compromised degranulation and proliferation abilities (Ma et al., 2022). Circulating CD8+ T cell subsets in patients with chronic HBV infection exhibited an exhausted phenotype.
CD8+ T cells from HBV carriers expressed higher levels of inhibitory receptors and produced lower levels of proinflammatory cytokines (interferon-gamma and TNF-alpha) and granzyme B (Jiang et al., 2022). Our current data showed a similar dysfunctional phenotype of CD8+ T cells from patients living with chronic HIV-1 infection. CD8+ T cells from patients living with HIV-1 infection exhibited reduced secretion of cytotoxic molecules and decreased expression of apoptosis-related ligands. These results were consistent with previous findings of the exhausted phenotype of CD8+ T cells during chronic HIV-1 infection (Dyrhol-Riise et al., 2001; Haridas et al., 2003; Kuerten et al., 2008). However, interferon-gamma secretion and inhibitory receptor expression were comparable between patients living with chronic HIV-1 infection and controls. This could happen for two reasons. On the one hand, increased expression levels of immune checkpoint molecules was usually observed in HIV-1-specific CD8+ T cells, whereas we evaluated the expression in total CD8+ T cells.
On the other hand, immune checkpoint molecule expression levels gradually decrease overtime as demonstrated in some longitudinal studies (de Armas et al., 2019; Rai et al., 2022). Collectively, CD8+ T cells from patients living with chronic HIV-1 infection exhibited dysfunctional phenotype.
The restoration of CD8+ T cell activity in patients living with HIV-1 infection is one popular topic in research aimed at identifying a functional cure. Importantly, cytokine-mediated activation of CD8+ T cells could lead to viral clearance. IL-15 potently promoted the survival and effector function of HIV-specific CD8+ T cells and might be helpful in enhancing the antiviral function of CD8+ T cells (Mueller et al., 2003). Both CD4+ and CD8+ T cells were able to produce IL-21 in response to HIV-1 infection. IL-21-producing CD4+ T cells are the best indicator of functional CD8+ T cells, and IL-21-producing HIV-1-specific CD8+ T cells are closely associated with viral control (Williams et al., 2011). In vitro IL-21 stimulation efficiently expanded HIV-1-specific CD8+ T memory stem cells to suppress HIV-1 replication (Wu et al., 2019). In vivo IL-21 therapy also controlled immune activation and maintained antiviral CD8+ T cell responses during acute simian immunodeficiency virus infection (Mendez-Lagares et al., 2017). IL-36 cytokine family members could promote CD8+ T cell activation and antitumor immune responses (Zhao et al., 2019).
We found that reduced circulating levels of IL-36gamma level were inversely correlated with CD8+ T cell counts in patients living with HIV-1 infection, indicating that IL-36gamma might contribute to the regulation of CD8+ T cell function during HIV-1 infection. Thus, we used stimulated CD8+ T cells from both controls and patients living with chronic HIV-1 infection with different concentrations of recombinant human IL-36gamma (1 and 10 ng/mL) for in vitro. Although IL-36R expression was comparable between the two groups, CD8+ T cells from controls and patients living with chronic HIV-1 infection exhibited differential responses to IL-36gamma stimulation. Neither 1 nor 10 ng/mL recombinant IL-36gamma stimulation affected the expression of inhibitory receptors (PD-1 and CTLA-4) or apoptosis-related ligands (TRAIL and FasL) in CD8+ T cells from controls or patients living with HIV-1 infection.
Interferon-gamma secretion was strongly increased in response to both concentrations in the two groups. Importantly, stimulation with 10 ng/mL IL-36gamma enhanced cytotoxic molecule (perforin, granzyme B, and granulysin) production in CD8+ T cells from both controls and patients living with chronic HIV-1 infection, but stimulation with 1 ng/mL IL-36gamma only promoted cytotoxic molecule expression in CD8+ T cells from patients living with chronic HIV-1 infection. One nanogram per microliter IL-36gamma also promoted perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. This indicated that CD8+ T cells from patients living with chronic HIV-1 infection exhibited increased responsiveness to IL-36gamma, and a lower concentration of IL-36gamma was sufficient for the activation of CD8+ T cells, probably through the enhancement of cytotoxic molecule expression during chronic HIV-1 infection. Further in vivo experiments are needed to elucidate of the regulatory effect of IL-36gamma on CD8+ T cells during HIV-1 infection.
Conclusion
In summary, reduced IL-36gamma level in patients living with chronic HIV-1 infection might be insufficient for the activation of CD8+ T cells, leading to CD8+ T cell exhaustion. IL-36gamma might be a potential therapeutic target for the functional cure of chronic HIV-1 infection.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.