
Abstract
Kyasanur Forest Disease Virus (KFDV) is a tick-borne flavivirus that causes life-threatening hemorrhagic fever in humans with case fatality rates of 3–5%. Relatively little is known about the mechanism of its pathogenesis or host immune responses to KFDV infection. Here, we investigated KFDV-specific cellular immune responses in the recovered cases of Kyasanur Forest Disease (KFD). Peripheral blood mononuclear cells of the recovered KFD cases and healthy controls were exposed to γ-inactivated KFDV antigen ex vivo. The proliferation index was determined using an enzyme-linked immunosorbent assay-based lymphoproliferative assay. The frequencies of CD4+ and CD8+ T cells expressing intracellular interferon (IFN)-γ in response to stimulation with γ-inactivated KFDV antigen were determined using flow cytometry. A significant increase in lymphoproliferation and a high frequency of CD4+ and CD8+ T cells secreting IFN-γ against γ-inactivated KFDV antigen were found in the recovered KFD group compared to the healthy control group. In conclusion, the study indicated the generation of cellular immune responses in individuals who recovered from KFD and can be used as indicators of cellular immunity in KFD vaccine studies.
Introduction
Kyasanur Forest Disease (KFD) is an acute viral disease caused by the KFD virus (KFDV) (Trapido et al., 1959). It was discovered in the Kyasanur forest of the Shimoga district in the state of Karnataka, India, in 1957 (Work et al., 1957). KFDV, a risk group 3 pathogen, is transmitted to animals and humans primarily by the hard tick Haemaphysalis spinigera (Trapido et al., 1959). KFDV was originally limited to the state of Karnataka but gradually disseminated to other regions of the southwest part of India, viz., Tamil Nadu, Kerala, Goa, and Maharashtra.
KFDV, a member of the family Flaviviridae, has a nonsegmented, linear, positive-sense RNA genome. It has a spherical size of 40–65 nm with an icosahedral nucleocapsid. Its genome has 10,774 nucleotide bases and encodes a single polypeptide. Post-translationally, the polypeptide is cleaved into three structural proteins (C, E, and M/prM) and seven nonstructural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5) (Dodd et al., 2011).
Despite the public health importance of KFD over the past six decades and the increased geographical expansion, we still lack effective countermeasures. There is no approved drug for KFD and the management of patients is primarily based on symptomatic treatment and supportive care. Although a vaccine based on formalin-inactivated chick embryo fibroblast is available, it is only partially effective against infection (Kasabi et al., 2013). Therefore, it is imperative to prioritize work on the development of effective vaccines against KFD.
An in-depth understanding of the host immune responses to the KFDV is lacking, and therefore protective immune markers or correlates of protection for KFD could not be characterized. Notably, Sathe et al.'s (1991) prior investigation noted higher circulating levels of interferon (IFN) during the acute phase compared to the convalescent phase. Recent findings by Devadiga et al. (2020) suggest robust T and B cell responses, particularly CD8+ T cells, in the majority of cases during the acute phase of KFD infection. Insights from other flaviviruses indicate that viral components within infected cells can induce signaling cascades, resulting in the production of type 1 interferons (IFN-α and IFN-β). This, in turn, activates the JAK/STAT signaling pathway, establishing an antiviral state and initiating adaptive immune responses (de la Fuente et al., 2017).
The epidemiological findings that the majority (>80%) of the KFDV-infected individuals recover without any consequences with long-lasting immunity indicate the possibility of developing an effective prophylactic vaccine against KFD (Iyer et al., 1959; Work et al., 1957). The underlying mechanism of protective immunity responsible for the resistance to reinfection as observed in the recovered cases of KFD would provide cues for the development of effective vaccine candidates. With this background, the present study was undertaken to elucidate KFDV-specific cellular immune responses against γ-inactivated KFDV antigen using peripheral blood mononuclear cells (PBMCs) from recovered cases of KFD and healthy individuals.
Materials and Methods
Study subjects
Recovered KFD (KFD-R) cases (n = 20) with a confirmed medical record were recruited from three primary health centers, namely Talkat, Sateli Bhedasi, and Morgav in the state of Maharashtra, India. Four milliliters of blood samples were collected into Li-Heparin vacutainer tubes (BD Biosciences, USA), gently mixed, and placed in the plastic secondary container in the upright position. The secondary container was placed into a suitable outer container with gel packs and transported to ICMR-National Institute of Virology (Pune, Maharashtra, India). Blood donors from the KFD-nonendemic region were included as healthy controls (HC) (n = 19).
Ethics statement
The study (No. NIV/IEC/Jan/2020/D-17) was approved by the Institutional Human Ethics Committee of ICMR-National Institute of Virology and carried out following the Indian Council of Medical Research Guidelines, 2017. Written informed consent was obtained from all the study participants before the collection of the blood sample.
Phenotyping of lymphoid cells in the peripheral blood
To understand the generalized cellular immune responses, we analyzed the phenotype and proportion of immune cells such as T cytotoxic, T helper, natural killer (NK), and B cells in the peripheral blood of KFD-R (n = 20) and HC (n = 19) individuals as previously reported (Kaushal et al., 2016; Kaushal et al., 2014). In brief, 0.1 mL of anticoagulated blood was stained with antibodies conjugated with fluorochrome along with their respective isotype controls. Two groups of sample tubes were prepared, one for B and NK cells (CD3-FITC, CD45-PerCP, CD19-APC, and CD16 + 56-PE) and one for T cells (CD3-PerCP, CD4-FITC, and CD8-PE). Following incubation at 4°C for 30 min, 2 mL of lysing buffer solution was added to each tube and kept at room temperature for 12 min.
Thereafter, the cells were washed by centrifugation, and the supernatant was gently removed. The cell pellets were resuspended in 500 μL wash buffer and analyzed on an Attune NxT Flow Cytometer on a minimum of 50,000 lymphocytes (R1). For analysis, lymphocytes were gated using side scatter (SSC) and forward (FSC) attributes and the proportions of CD4+ and CD8+ T cells were gated on CD3+ T cells. Similarly, the frequencies of CD19+ and CD16++56+ cells were acquired on CD45+ cells.
Preparation of KFDV antigen
The Maximum Containment Facility of ICMR-National Institute of Virology, provided the inactivated KFDV antigen. In brief, the KFDV stock (NIV12839) was prepared by propagating the virus in the Baby Hamster Kidney (BHK)-21 cells. After harvesting, the culture supernatant was irradiated with a cobalt-60 source in a GC-5000 gamma chamber. It was exposed to 24 kGy for 1 h to inactivate the virus. The irradiated KFDV stock was subjected to two passages in the BHK-21 cell line and observed for 7 days after infection to confirm the absence of virus growth, thereby certifying that the virus is inactivated. The γ-inactivated KFDV antigen stock was then filtered through a 30 kDa filter. Following protein estimation, the inactivated whole virus was used as an antigen for stimulating PBMCs.
Lymphoproliferative assay
Lymphoproliferation was determined as previously described (Kaushal et al., 2016; Kaushal et al., 2014). PBMCs from KFD-R (n = 20) and HC (n = 19) were isolated from the heparinized blood by the density sedimentation method using Ficoll-Paque. The buffy coat was gently collected, washed, and suspended in complete RPMI-1640 media with L-glutamine, 10% fetal bovine serum, streptomycin (100 μg/mL), and penicillin (100 U/mL). PBMCs at 1 × 106 cells/mL concentration were cultured in triplicate in 96-well culture plates and challenged with either KFDV antigen (10 μg/mL) or phytohemagglutinin M (PHA-M) (10 μg/mL) for 96 hours in a humidified 37°C/5% carbon dioxide incubator.
At 80 hours of incubation, 20 μL of 5-bromo-2′-deoxyuridine labeling solution was mixed and the samples were reincubated for another 16–18 hours. A commercial enzyme-linked immunosorbent assay (ELISA) kit (Biotrak™ Cell Proliferation ELISA System; GE Healthcare, USA) was used to evaluate lymphoproliferation. The proliferation index (PI) was determined as the ratio of the optical density of stimulated and unstimulated PBMCs from each study participant.
Intracellular staining and flow cytometry
Freshly isolated PBMCs (106 cells/mL) from KFD-R (n = 20) and HC (n = 19) individuals were incubated with KFDV antigen (10 μg/mL) in a culture plate for 96 hours at 37°C. GolgiStop (1 μg/mL; BD Biosciences) was added to PBMCs culture to inhibit IFN-γ secretion and incubated for another 6 hours. The cells were carefully collected and washed twice with washing solution (1 × phosphate buffered saline [PBS]) at 200 × g for 5 min. The cells were then surface-stained with antibodies conjugated with fluorochromes such as CD3-APC-H7, CD4-PerCP, and CD8-V500 (BD Biosciences) along with appropriate Fluorescence Minus One at 4°C for 30 min. For intracellular IFN-γ analysis, cells were fixed and permeabilized by the Cytoperm/Cytofix Plus Kit (BD Biosciences) as per the manufacturer's instructions.
After 20 min of incubation and washing, cells were labeled with anti-human IFN-γ-PE. Samples were acquired and analyzed using Attune NxT Flow Cytometry on a minimum of 50,000 lymphocytes (R1). For analysis, lymphocytes (R1) were gated using FSC and SSC attributes, and the proportions of CD4+ and CD8+ T cells were analyzed on CD3+ T cell populations. The frequencies of CD4+ IFN-γ+ and CD8+ IFN-γ+ cells were analyzed on CD4+ and CD8+ T cell populations, respectively.
Determination of humoral response
KFDV-specific IgG levels were assessed in plasma samples from KFD-R (n = 20) and HC individuals (n = 19) using the ELISA method, as previously outlined with modifications (Chamakh-Ayari et al., 2014). In brief, 96-well flat-bottom ELISA plates were coated with 100 μL of KFDV antigen (10 μg/mL) in bicarbonate buffer (pH 9.6) at 4°C overnight. After incubation, the plates were gently washed and blocked with 200 μL of 1% bovine serum albumin at 37°C for 1 hour, followed by three washes with PBS containing 0.1% Tween 20 (PBST). The plates were then incubated for 1 h with diluted sera (1:100) from HC and KFD-R. Subsequently, the wells were washed three times with PBST and incubated with horseradish peroxidase-conjugated anti-human IgG (1:5,000) for 30 min at 37°C.
After another round of washing, the O-phenylenediamine dihydrochloride substrate was added to develop color. The reaction was stopped by adding 1 N H2SO4, and the optical density (OD) was measured at 492 nm using an ELISA reader. Each reaction was performed in triplicate. The cutoff for determining KFDV-specific IgG positivity was based on the mean plus three times the standard deviation (SD) of OD values observed in the HC group.
Statistical analysis
Statistical significance between the two study groups was determined by the nonparametric, Mann–Whitney U test using GraphPad Prism 9.5 software (GraphPad Software, USA). A p < 0.05 was considered significant.
Results
Study subjects
The study included KFD-R cases (n = 20; male/female [M/F]: 10/10), (age range, years, 30–72; age, mean ± SD, 49 ± 11.57). These are healthy individuals with a recorded history of KFD infection who recovered at least 6 months back from the time of blood collection. The range of post-treatment duration for the KFD-R group was 0.5 to 5.75 years, with a duration, mean ± SD, 3.06 ± 1.45. The majority (14/20) of the KFD-R group participants were vaccinated with at least two doses of the KFD vaccine. The clinical symptoms during the acute disease phase were fever (70%), headache (65%), myalgia (50%), vomiting (40%), diarrhea (25%), general weakness (15%), abdominal pain (5%), nausea (10%), and black stool (5%).
Further, we did not find differential symptoms associated with either vaccinated or nonvaccinated individuals. All individuals who had recovered from KFD tested positive for anti-KFDV IgG. Healthy control individuals (n = 19; M/F: 10/9) (age range, years, 18–50; age mean ± SD, 25.92 ± 9.07) from KFD nonendemic areas were included.
Lymphocyte profile in KFD-R cases and HC groups
The frequencies of cells expressing CD3, CD8, CD4, CD19, or CD16 + 56 among the total PBMCs were not different between the KFD-R and HC groups (Supplementary Table S1; Supplementary Fig. S1).
Enhanced lymphoproliferative responses against inactivated KFDV antigen in the KFD-R group
Lymphoproliferative response (PI) to KFDV antigen ex vivo was assessed in the PBMCs isolated from KFD-R (n = 20) and HC groups (n = 19). All the study participants showed high lymphoproliferation with PHA-M (HC, PI mean ± SD, 6.428 ± 2.701; KFD-R, 6.222 ± 2.725). In response to stimulation with γ-inactivated KFDV antigen, the mean lymphoproliferative response of KFD-R (PI mean ± SD, 2.034 ± 0.617, p < 0.0001) was significantly higher compared to the HC group (PI mean ± SD, 1.178 ± 0.0.248) (Fig. 1). However, the mean lymphoproliferative response between vaccinated (n = 14, PI mean ± SD, 1.956 ± 0.4052) and nonvaccinated (n = 6, PI mean ± SD, 2.217 ± 0.9814) cases of the KFD-R group was found to be comparable (Supplementary Fig. S2).
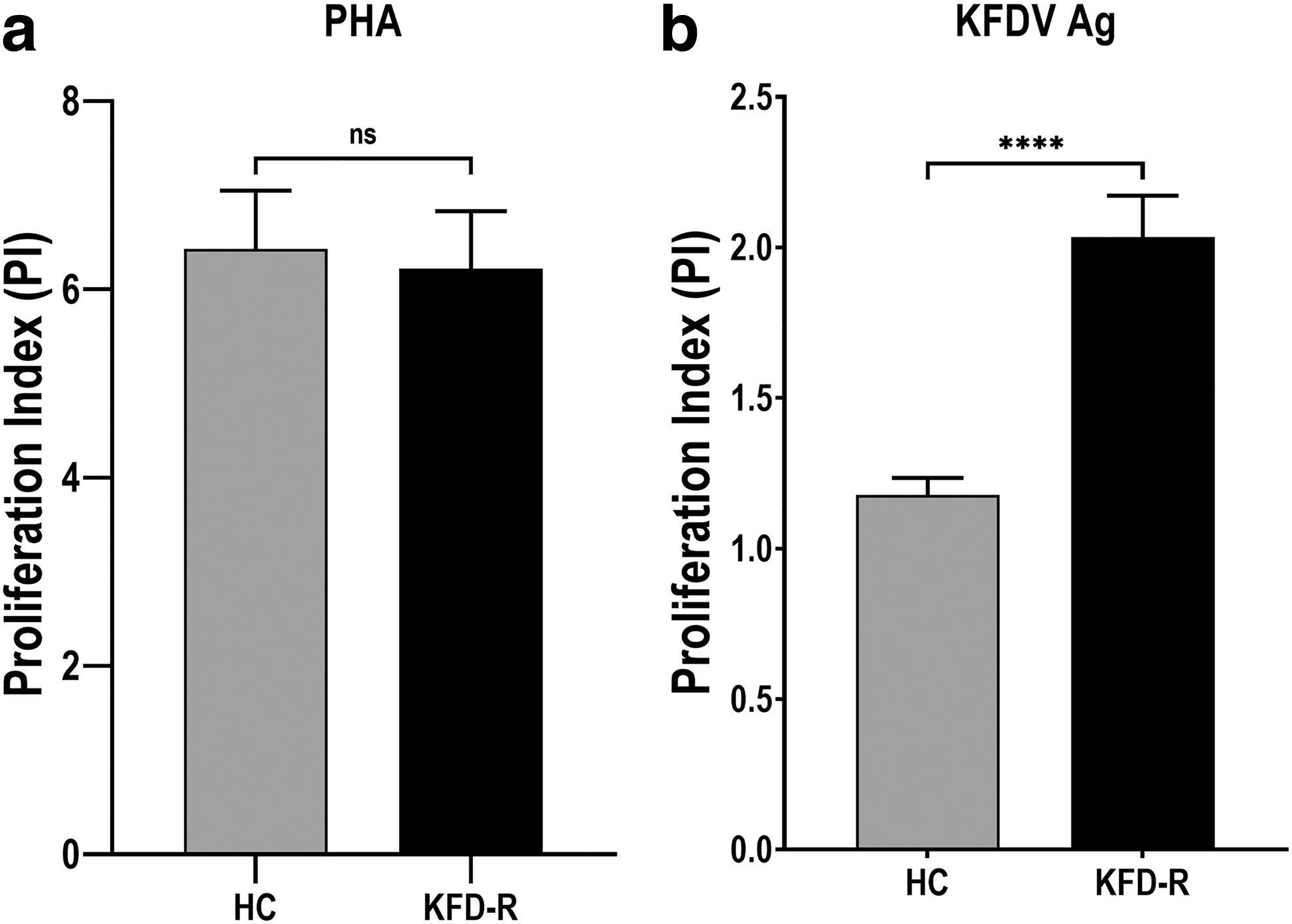
Comparative lymphoproliferative responses in healthy controls and recovered cases of KFD in vitro. PBMCs from KFD-R (n = 20) and HC group (n = 19) were incubated with
T cells secrete IFN-γ in response to inactivated KFDV antigen in the KFD-R group
The phenotypes and frequencies of IFN-γ-producing T cells upon stimulation with inactivated KFDV antigens of PBMCs obtained from the HC and KFD-R groups were investigated. In both the HC and KFD-R groups, the frequency of IFN-γ producing T cells upon stimulation with KFDV antigens was normalized by subtracting the IFN-γ-producing T cells in unstimulated cultures. In the KFD-R group, the percentage of IFN-γ-producing CD4+ T cells was significantly high (mean ± SD, 0.41 ± 0.426, p < 0.01) after stimulation with the inactivated KFDV antigens compared to the HC group (mean ± SD, 0.097 ± 0.223). Similarly, the percentage of IFN-γ-producing CD8+ T cells after stimulation with the inactivated KFDV antigens in the KFD-R group (mean ± SD, 0.744 ± 0.694, p < 0.0001) was significantly higher compared to the HC group (mean ± SD, 0.053 ± 0.108; Fig. 2; Supplementary Fig. S3).
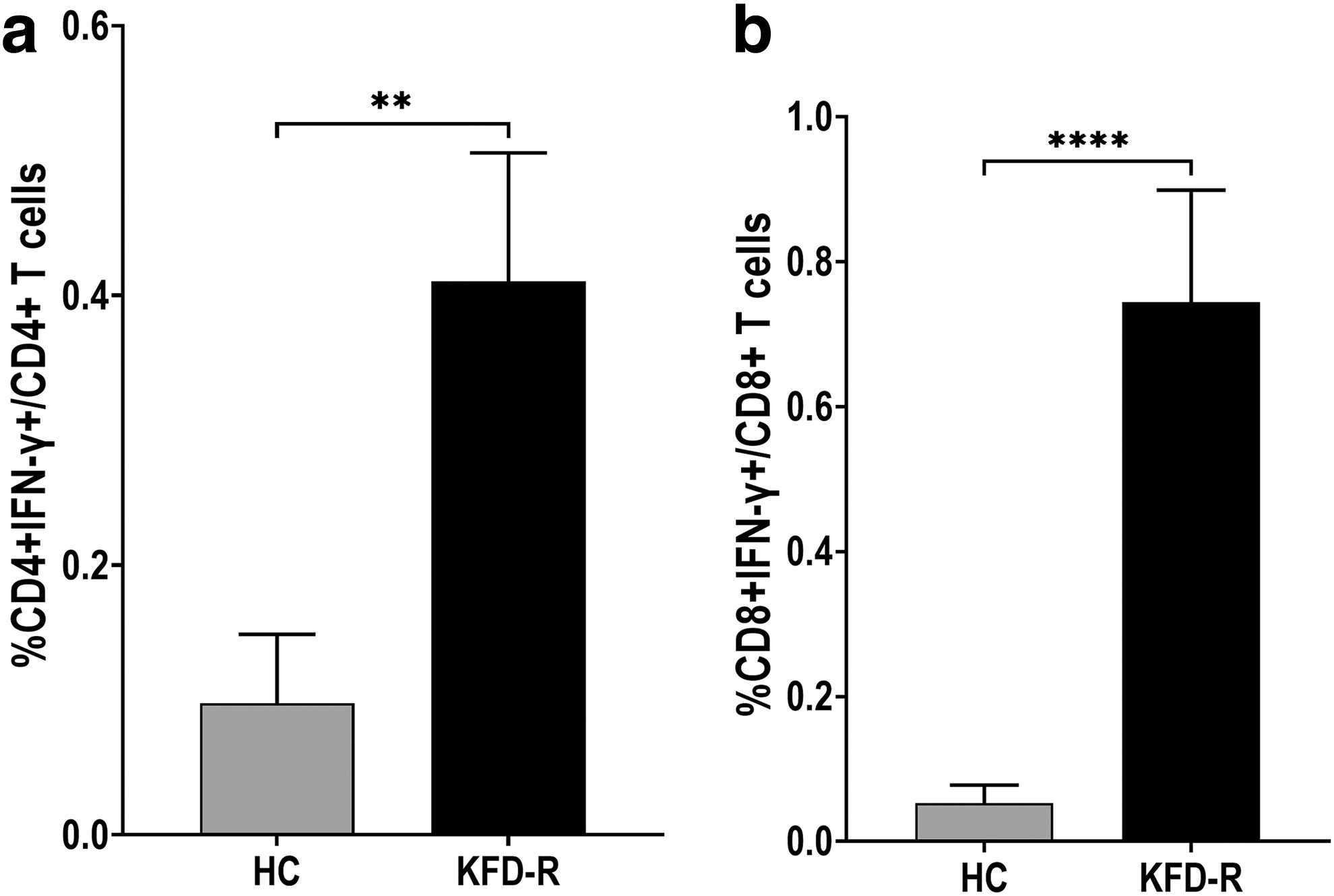
Percentage of IFN-γ-producing T cells after stimulation of PBMCs with inactivated KFDV antigens obtained from KFD-R (n = 20) and HC (n = 19). The results are expressed as a bar diagram of the group mean of two study groups. Intracellular IFN-γ analysis in T cells upon stimulation with inactivated KFDV antigens.
Furthermore, upon stimulation with γ-inactivated KFDV antigen, the % CD4+IFN-γ+ (mean ± SD, 0.415 ± 0.38, p > 0.05) and % CD8+ IFN-γ+ T cells (mean ± SD, 0.844 ± 0.677, p > 0.05) in the vaccinated (n = 14) was found comparable to unvaccinated (n = 6; CD4+IFN-γ+, 0.399 ± 0.559; CD8+IFN-γ+, 0.509 ± 0.738) cases of recovered KFD group (Supplementary Fig. S2).
Humoral responses to KFDV antigen in the recovered KFD group
We attempted to utilize the relative abundances of anti-KFDV IgG to discriminate immunity among the KFD-R and HC groups. Plasma samples from the KFD-R group (n = 20) and HC group (n = 19) were tested for the presence of anti-KFDV IgG. The results are represented as mean absorbance at an OD of 492 nm wavelength (mean ± SD OD492). The anti-KFDV IgG level was significantly elevated in the KFD-R group (0.36 ± 0.096, p < 0.0001) compared to the HC group (0.107 ± 0.045). However, the anti-KFDV IgG level in the vaccinated group (n = 14; 0.362 ± 0.106, p > 0.05) was found to be comparable to that of the non-vaccinated group (n = 6; 0.356 ± 0.078) in the recovered KFD individuals (Supplementary Fig. S4).
Discussion
Despite the frequent occurrence of KFD in the endemic areas, comparatively little is known about cellular immune responses to KFDV infection. In this study, the frequencies of different lymphoid cells, such as T, B, and NK cells, in the peripheral blood of the KFD-R and HC groups were evaluated and were comparable between the HC and KFD-R groups. Moreover, comparable lymphoproliferation in response to phytohemagglutinin observed in both the KFD-R and the HC groups demonstrated that the generalized cellular immunity was not impaired. Thereafter, KFDV-specific cellular immune responses were evaluated ex vivo by determining lymphoproliferation upon stimulation with γ-inactivated KFDV antigen. The study demonstrated high lymphoproliferation in the KFD-R group compared to the HC group. The higher lymphoproliferation in the KFD-R group could be due to circulating KFDV-specific memory T cells that responded positively to the KFDV antigen challenge and is indicative of pronounced cellular immunity.
This finding reiterates the fact that pre-exposed individuals can elicit an enhanced cell-mediated immune response against the KFDV. An in-depth understanding of the protective cellular immunity generated upon natural infection provides important cues for the design and formulation of newer vaccine candidates (Macallan et al., 2017). In the present study, the majority of the participants in the recovered KFD group were vaccinated before contracting the KFDV infection. Moreover, the cell-mediated immune responses to inactivated KFDV antigen were comparable between vaccinated and nonvaccinated individuals within the recovered KFD group. This indicates that there is a limited advantage of the existing vaccine in eliciting cellular immunity. This further reiterates the fact that there is a need for the formulation of an effective vaccine against KFD.
The results also revealed a higher frequency of both CD8+ and CD4+ T cells secreting IFN-γ in the KFD-R group upon the KFDV antigen challenge. Specifically, the CD8+ T response was higher compared to the CD4+ T cell response. This indicated the generation of cellular immunity in the pre-exposed individuals with the KFDV. Our findings corroborated the study by Devadiga et al. (2020), who reported strong T and B cell responses, mainly CD8+ T cells, during the active phase of KFD infection. Further, they also noted that the clearance of the KFDV from the blood concurred with the peak activation of CD8+ T cells and the appearance of KFDV-specific IgG (Devadiga et al., 2020).
However, their work was based on the phenotyping of immune cells in the peripheral blood in acute and convalescent blood samples of KFD, whereas the present study involved the challenge of PBMCs from recovered KFD cases with KFDV antigen ex vivo. T cells with Th1 type cytokines such as IFN-γ provide protection against viral infection (Kannanganat et al., 2007; Lee and Ashkar, 2018; Weaver et al., 2013). IFN-γ enhances the expression of major histocompatibility complex (MHC) I and MHC II in the nearby viral-infected cells and APCs, respectively (Schroder et al., 2004; Steimle et al., 1994). In addition, it also stimulates the secretion of IL-12 and the expression of CD80 costimulatory molecules in APCs, an important component of Th1 polarization (Bauvois et al., 2009; Ma et al., 1996).
Moreover, elevated humoral responses following infection were evident in the recovered KFD group, as indicated by increased plasma IgG levels. Nevertheless, no discernible disparities in humoral responses were observed between vaccinated and nonvaccinated individuals among the recovered KFD cases. The comparable cellular and humoral immune responses in vaccinated and nonvaccinated groups among the recovered KFD group are supported by a recent study that demonstrated no significant difference in the infection rates, clinical manifestations, or outcomes between the vaccinated and nonvaccinated groups among KFD-confirmed cases (Munivenkatappa et al., 2023).
Overall, the results of lymphoproliferation and intracellular IFN-γ demonstrated the generation of cellular immune responses in the KFD-R group, which supports the epidemiological findings that the majority of recovered KFD cases remain protected against reinfection (Achar et al., 1981; Holbrook, 2013). This suggests the feasibility of the development of an effective vaccine for KFD (Kasabi et al., 2013). Since the present study was conducted on individuals with a history of natural infection, the findings may provide a roadmap for the development and evaluation of any potential vaccine candidates against KFD.
To the best of our knowledge, this is the first study that indicated the presence of cellular immunity in individuals who recovered from KFD, with a postrecovery interval exceeding 6 months. A higher lymphoproliferation index and enhanced frequency of CD8+ and CD4+ T cells secreting IFN-γ in response to challenge with the viral antigen suggests their utility as a marker of cellular immune response in KFD vaccine studies.
Footnotes
Acknowledgments
We acknowledge the support received from the field team, which includes Anuja Desai, Chandrashekhar Naik, Supriya Palav of PHC, Talkat; Gayatri Paste, Arun Gawas of PHC, Morgav; Shruti Shirodkar, Jayawant Tulaskar, and Reshma Kshirsagar of PHC, Sateli Bhedasi. We are thankful to Dr. Badri Narayan Natarajan of Thermo Fisher Scientific for helpful discussions on Attune NxT flow cytometry.
Authors' Contributions
Conceptualization and methodology: H.K. Resource: R.S.K., T.C., M.M.K., P.D.Y., P.A., V.A.P., C.S., A.M.S., and R.R.S. Investigation: H.K., S.D., S.A.C., P.D.Y., A.M.S., and R.R.S. Formal analysis: H.K. and K.A. Data curation: H.K., K.A., S.D., and S.A.C. Writing, review and editing: H.K., K.A., and P.D.Y. Supervision: H.K. and K.A. Project administration: H.K. Funding acquisition: H.K. All authors have read and agreed to the published version of the article.
Institutional Review Board Statement
The study was approved by the Institutional Ethics Committee of ICMR-National Institute of Virology (NIV/IEC/Jan/2020/D-17 dated January 15, 2020).
Informed Consent Statement
Written informed consent was obtained from all the study participants for the blood sample collection and subsequent analysis.
Data Availability Statement
All the data have been provided in the article.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This research was funded by the Indian Council of Medical Research (Grant No. VIR/25/2020/ECD-1).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.