
Abstract
Zika virus (ZIKV) is an emerging flavivirus associated with several neurological diseases such as Guillain-Barré syndrome in adults and microcephaly in newborn children. Its distribution and mode of transmission (via Aedes aegypti and Aedes albopictus mosquitoes) collectively cause ZIKV to be a serious concern for global health. High genetic homology of flaviviruses and shared ecology is a hurdle for accurate detection. Distinguishing infections caused by different viruses based on serological recognition can be misleading as many anti-flavivirus monoclonal antibodies (mAbs) discovered to date are highly cross-reactive, especially those against the envelope (E) protein. To provide more specific research tools, we produced ZIKV E directed hybridoma cell lines and characterized two highly ZIKV-specific mAb clones (mAbs A11 and A42) against several members of the Flavivirus genus. Epitope mapping of mAb A11 revealed glycan loop specificity in Domain I of the ZIKV E protein. The development of two highly specific mAbs targeting the surface fusion protein of ZIKV presents a significant advancement in research capabilities as these can be employed as essential tools to enhance our understanding of ZIKV identification on infected cells ex vivo or in culture.
Introduction
Zika virus (ZIKV) was first identified in the Zika forest of Uganda in 1947 followed by subsequent identification as a human pathogen in 1952 (Dick et al., 1952; MacNamara, 1954). Infections remained cryptic and sporadic until the 2007 Yap Island outbreak (Duffy et al., 2009; Olson et al., 1981; Simpson, 1964). The 2015 Brazil outbreak (Campos et al., 2015; Cardoso et al., 2015) marked the first documented instance ZIKV was associated with microcephaly, indicating the possibility of transplacental transmission and teratogenicity (Mlakar et al., 2016; Schuler-Faccini et al., 2016).
While most infections in healthy adults are asymptomatic, ZIKV, like other flavivirus infections, causes a flu-like illness with fever, rash, joint pain, and headaches (Armstrong et al., 2016).
ZIKV is mainly transmitted by Aedes mosquitoes and has overlapping distribution with other flaviviruses such as Dengue virus (DENV) and West Nile Virus (WNV) (Alaniz et al., 2017; Bhatt et al., 2013; Chancey et al., 2015; Dash et al., 2013). The envelope (E) protein, situated on the surface of mature viral particles, is responsible for receptor-mediated viral fusion and is a major target for the humoral anti-viral response (Prasad et al., 2017; Rey and Lok, 2018).
E proteins of flaviviruses share a common protomer architecture (Keasey et al., 2017; Reisen et al., 2014); thus, antibody-based differentiation solely based on surface protein recognition is complicated due to conserved epitopes (Musso et al., 2015).
Recently, a new class of neutralizing monoclonal antibodies (mAbs) reactive to Domain I has been characterized, recognizing linear epitopes near the glycan loop that were specific for contemporary strains of ZIKV but not the MR766 strain (Qu et al., 2020). Here, we generate and characterize two ZIKV E-specific mAbs (A11 and A42) that recognize the same epitope but are non-neutralizing and can discriminate ZIKV against other flaviviruses.
We have additionally generated a chimeric monoclonal antibody (cmAb) A42 and demonstrate retention of its binding properties, making these mAb useful tools for molecular techniques to detect ZIKV.
Methods
Ethical statement
Mouse experiments were approved by the University of Hawai`i's Institutional Animal Care and Use Committee (IACUC). The John A. Burns School of Medicine (JABSOM) Laboratory Animal Facility is accredited by the American Association for Accreditation of Laboratory Animal Care (AAALAC). Mice were bred in colonies at JABSOM from original stocks obtained from Taconic Bioscience (Hudson, NY).
Mouse immunization
Soluble recombinant ZIKV E protein was produced as previously described (To et al., 2018). Eight BALB/c mice were immunized intramuscularly with two doses of 10 μg of ZIKV E protein formulated with 2% Alhydrogel adjuvant (InvivoGen, San Diego, CA) 2 weeks apart. Sera were collected from mice after 2 weeks via a tail incision, and ZIKV E specific antibodies were measured using the enzyme-linked immunosorbent assay (ELISA) described next. Mice with high anti-ZIKV E titers were given a booster dose 5 days before splenectomy.
Enzyme-linked immunosorbent assay
Dynex Immulon 96-well flat bottom plates (VWR, Radnor, PA) coated overnight with 200 ng of soluble ZIKV E protein per well were blocked with phosphate buffered saline (PBS) +1% bovine serum albumin (BSA) and washed with PBS +1% BSA +0.02% Tween 20 (PBS-BT) before addition of mAb A11, A42 or 4G2, serially diluted fourfold starting at 10 μg/mL in triplicates. After a PBS-BT wash, anti-mouse-IgG AP (Southern Biotech, Birmingham, AL) was added at a 1:3,000 dilution. The plate was developed with Alkaline Phosphatase Yellow (pNPP) liquid substrate system (Sigma, St. Louis, MO), and the optical densities were measured using a VICTOR Multilabel Plate Reader (PerkinElmer, Waltham, MA). To evaluate binding at mildly acidic pH, an incubation step with PBS at pH of 7.4, 6.0, and 5.0 after the initial blocking step (Tsai et al., 2013) was included before incubation with detection antibody and substrate.
Hybridoma generation and mAb purification
Harvested splenocytes were fused with P3X63Ag8.653 cells as previously described (Bertino et al., 2019; Kohler and Milstein, 1975; Peckham et al., 2010). Hybridoma cell lines underwent three rounds of subcloning in RPMI 1640 media (Invitrogen, Carlsbad, CA) with 10% fetal bovine serum (FBS) and 1% Pen/Strep before being expanded into T25 flasks.
mAbs were purified from the culture supernatant using a 1 mL HiTrap Protein G HP column (GE Healthcare, Piscataway, NJ). The purified mAbs were eluted using a 20 mM glycine buffer, pH 2.5, and buffer exchanged into PBS +0.01% azide. The concentration was determined by ultraviolet (UV)280 absorption and adjusted to 1 mg/mL.
Western Blot analysis
Equal sample loadings were mixed with NuPAGE™ LDS sample buffer (Thermo Fisher Scientific, Waltham, MA) and boiled for 10 min before resolving on NuPAGE 4–12% Bis-Tris Protein Gels (Thermo Fisher Scientific). Protein was transferred onto nitrocellulose membranes using a Bolt Mini Blot Module (Thermo Fisher Scientific) and blocked with 1% nonfat-milk (Nestlé USA, Inc., Solon, OH) in PBST at 4°C before probing with anti-ZIKV E mAbs A11, A42, or pan-flavivirus mAb 4G2 at 1 μg/mL overnight. After washing with PBST, membranes were incubated with goat anti-mouse IgG AP-conjugate, at 1:3,000 (Southern Biotech) and developed with NBT/BCIP substrate solution (Promega, Madison, WI).
Microsphere immunoassay
Magnetic microsphere beads MagPlex® (Luminex Corporation, Austin, TX) were covalently coupled to ZIKV, DENV-2, DENV-3, WNV E protein, and BSA as described previously (To et al., 2018). Mouse sera and cell culture supernatants were diluted 1:200 in PBS-1% BSA and added to a black-sided 96-well plate in triplicates. To assess cross-reactivity to different flavivirus E proteins, ZIKV mAbs were adjusted to 10 μg/mL followed by a series of five fourfold dilutions before addition to beads in a 1:1 ratio.
Incubations were done at room temperature (RT) with agitation on a plate shaker for 1 h. The beads were washed with PBS-BT and R-PE-conjugated anti-mouse IgG, IgG1, IgG2a, IgG2b, or IgG3 [F(ab′)2 fragments; Southern Biotech], diluted to 1:200, were added to the respective wells for 1 h. Beads were washed and resuspended in drive fluid before measuring the median fluorescence intensity using a Magpix instrument (Luminex Corporation).
Virus stock and cell culture
Vero cells were grown in Hanks BBS Medium199, containing 2 mM L-glutamine, 5% FBS +1% Pen/Strep (Invitrogen, Carlsbad, CA), and maintained at 37°C, humidified with 5% CO2. ZIKV (PRVABC59 strain), DENV-1 (Hawaiʻi strain), DENV-2 (NGC1056 strain), DENV-3 (H87 strain), DENV-4 (H241 strain), or WNV (NY99 strain) were obtained from the ATCC (Manassas, VA).
The Spondweni virus (SPONV, SA AR94 strain) was obtained from the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA) at the UTMB. Viruses were amplified in Vero cells, and virus titers were determined by plaque assay as described previously (Kumar et al., 2017). Virus-infected cell lysates were prepared using 1% NP-40 lysis buffer as described previously (Tsai et al., 2015).
Immunofluorescence assay
Vero cells seeded on glass coverslips were infected with DENV-2, DENV-4, or ZIKV at an multiplicity of infection of 2 for 1 h. Forty-eight hours post-infection, cells were fixed with 4% paraformaldehyde and permeabilized with a 0.25% Triton X-100 solution. Each well was blocked with 5% BSA in PBS for 1 h at RT before incubation with mAbs A11, A42, or 4G2, all diluted to 2.5 μg/mL, for 1 h. Cells were washed with PBS-BT before goat anti-mouse IgG-Alexa Fluor 488 (Thermo Fisher Scientific) was added at 4 μg/mL for 1 h. Cells were washed again, and coverslips were mounted facedown onto microscope slides prepared with ProLong™ Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific) and imaged with an Olympus BX41 fluorescence microscope.
Epitope mapping
A 164-peptide array of 15-mers (12 aa overlap) spanning the entire length of the ZIKV E protein, strain PRVABC59 (NR-50553; BEI Resources) was diluted in sodium-bicarbonate buffer (pH 9.5) to 1 mg/mL. Pools of five peptides were plated overnight on Dynex Immulon 96-well flat bottom plates, in duplicates, at a final concentration of 2 μg/mL for each peptide.
After blocking with PBS-1% BSA, mAbs A11 and A42, at 2.5 μg/mL were added to respective wells for 1 h, washed with PBS-BT, and developed using the ELISA protocol shown earlier. Individual peptides from peptide pools that yielded positive results were then tested individually at 1 mg/mL in duplicate using the same protocol.
Site-directed mutagenesis
Overlapping mutagenesis primers encoding for M151A, V153A, R164A, and MV151/3AA were designed using QuikChange software (Agilent Technologies, Inc., Santa Clara, CA). Mutant plasmids were amplified with Phusion® High-Fidelity DNA Polymerase (New England Biolabs, Ipswich, MA) before being treated with DpnI overnight at 37°C and transformed into competent NEB 5-alpha Escherichia coli (New England Biolabs).
Plasmids containing the desired mutations were transfected into Drosophila S2 cells using Lipofectamine LTX with PLUS reagent (Invitrogen, Carlsbad, CA). S2 cells were transiently induced with 200 μM CuSO4. Supernatants were harvested after 48 h and resolved using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and western blots were developed as described earlier.
Antibody variable domain sequencing and analysis
RNA from hybridoma cells was extracted using TRIzol™ Reagent (Invitrogen, Waltham, MA) and converted to complementary DNA (cDNA) using SuperScript™ III Reverse Transcriptase (Invitrogen, Waltham, MA) with 50 μM oligo(dT). Heavy and light chain variable domain genes were amplified with primers pools (Babrak et al., 2017) using GoTaq® Green 2x Master Mix (Promega).
The polymerase chain reaction (PCR) product was cloned using the TOPO™ TA Cloning™ Kit and transformed into One Shot™ TOP10 E. coli cells (Invitrogen, Waltham, MA). Selected colonies were grown in LB broth with carbenicillin, and plasmid vectors were purified using the QIAprep Spin Miniprep Kit (Qiagen, Hilden, Germany). Sanger-confirmed sequences were aligned and annotated using IgBLAST (Ye et al., 2013).
Recombinant, cmAb A42 expression
To generate plasmids for mouse/human cmAb A42 IgG1, gene fragments encoding the variable heavy and kappa light chain domains of mAb A42 were synthesized (Twist Bioscience, San Francisco, CA). Secretion signals L1 and H7 were used for the light and heavy chain, respectively (Haryadi et al., 2015). Both chains were cloned into pTrioz-hIgG1 (InvivoGen) using NEBuilder® HiFi DNA Assembly Master Mix (New England Biolabs) and transfected at 1 μg/mL, final concentration, using Lipofectamine 3000 (Thermo Fisher Scientific) into HEK 293T cells (ATCC).
Cells were grown for 14 days at 37°C and 5% CO2 before the culture supernatant was harvested. Recombinant cmAb A42 was purified using a 5 mL Protein A column (Cytiva, Marlborough, MA) and eluted with 20 mM glycine buffer, pH 2.5.
Statistical analysis
EC50 values were calculated using the nonlinear regression analysis (sigmoidal dose-response, variable slope) on GraphPad Prism 7.0 (GraphPad Software, Boston, MA).
Results
Generation and characterization of ZIKV Envelope specific mAbs
BALB/c mice were immunized intramuscularly with soluble, recombinant ZIKV E antigen (H/PF/2013 strain) formulated with alum adjuvant. Splenocytes from top responders were pooled to generate hybridoma cell lines. After two rounds of subcloning, two hybridoma-produced mAbs, A11.6 and A42.11 (referred to hereon as mAbs A11 and A42, respectively) were highly specific for the ZIKV E but nonreactive to other flavivirus E proteins of WNV, DENV-2, and DENV-3 (Fig. 1A, B). By contrast, pan-flavivirus mAb 4G2, which recognizes a conserved epitope on the flavivirus E (Crill and Chang, 2004), recognized WNV, DENV-3, and DENV-2 E in descending affinity (Fig. 1C).

ZIKV E-specific mAb selection. The binding activity of
The EC50 for mAbs A11 and A42 with ZIKV E were calculated to be 0.047 (95% confidence interval [CI]: 0.4–0.5) and 0.217 (95% CI: 0.044–0.55) μg/mL, respectively. In addition, both mAbs were determined to be IgG1 (data not shown). To evaluate the binding characteristics of the two mAbs in acidic environments, the immune complex was incubated at pH 7.4, 6.0, or 5.0. mAb A11 had the strongest affinity at pH 7.4; however, the EC50 significantly increased to 0.362 (95% CI: 0.29–0.45) μg/mL at pH 5.0 (Fig. 1D, E).
The binding activity of both mAbs A42 and 4G2 did not change in mild acidic environments. Despite the strong affinity and specificity toward ZIKV, neither mAbs A11 nor A42 exhibited any ZIKV neutralization by a standard plaque reduction neutralization assay (data not shown).
mAbs A11 and A42 are ZIKV-specific and do not cross-react with other flaviviruses
We further assessed the binding specificity of mAbs A11 and A42 to common flavivirus antigens by Western blot and immunofluorescence staining. First, we confirmed the specificity of both mAbs A11 and A42 to ZIKV E by testing their reactivity against a panel of recombinant flavivirus E proteins (Fig. 2A). No reactivity was observed with WNV, DENV-2, and DENV-3 E. Conversely, mAb 4G2 bound all tested flavivirus-E proteins.
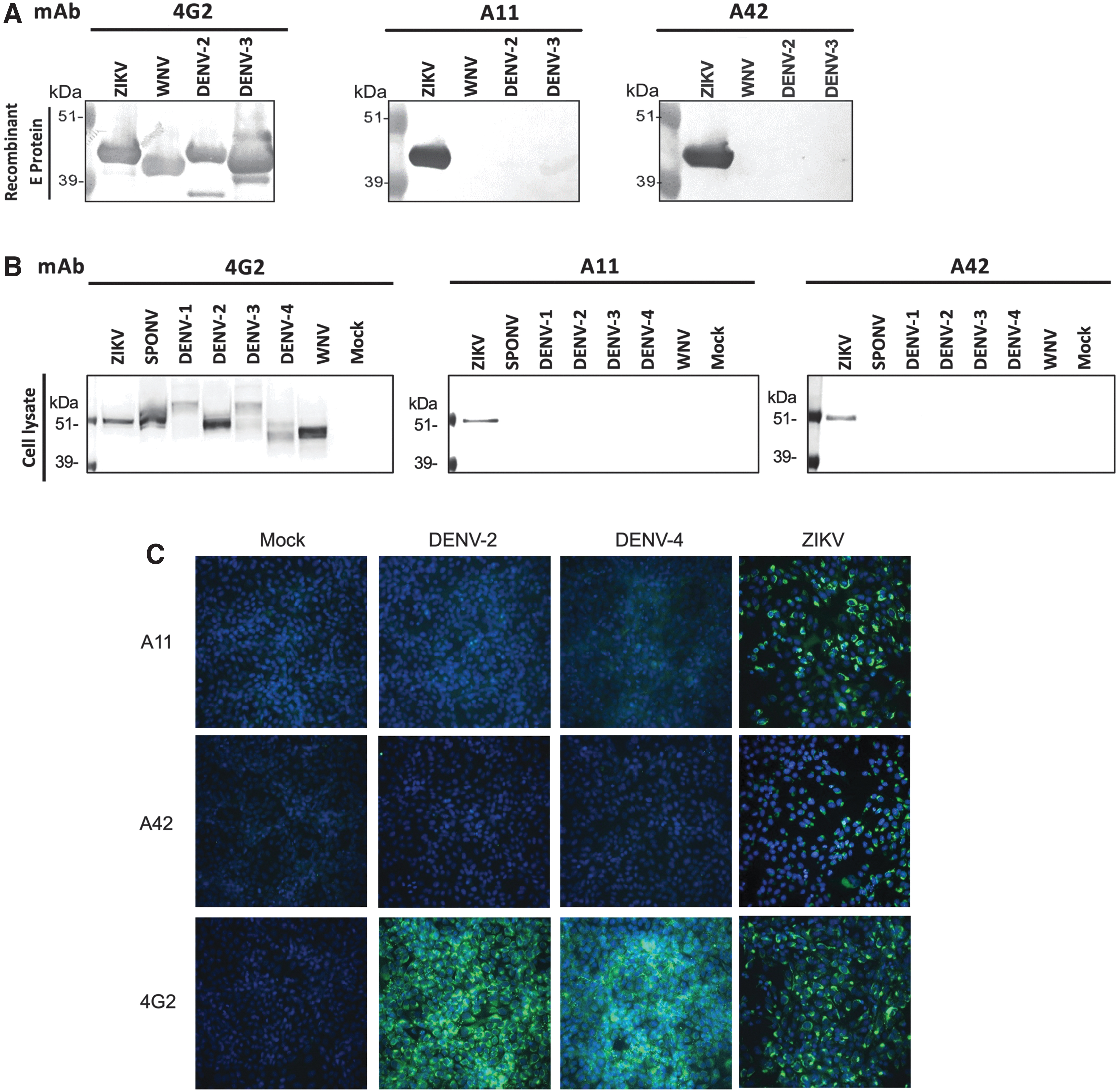
Binding specificities of mAbs A11 and A42 by Western blot analysis and immunofluorescence assay. Binding specificities of mAbs 4G2, A11 and A42, at 1 μg/mL, were determined by Western blot analysis using
Further, both mAbs recognized specifically ZIKV-infected Vero cell lysates but not E from other flaviviruse, including DENV-1-4, WNV, and SPONV, despite the latter being 72% homologous to ZIKV E (Fig. 2B). Next, we demonstrated that both mAbs A11 and A42 maintain their specificity when used for immunofluorescence staining with Vero cells infected with ZIKV, DENV-2, and DENV-4. Compared with mAb 4G2, which recognized cells infected with all three viruses, mAbs A11 and A42 only detected ZIKV-infected cells, but not cells expressing the E protein as a result of DENV-2 and DENV-4 infection (Fig. 2C).
Epitope mapping of mAb A11 supports binding to a unique epitope of the ZIKV envelope protein at the 150-loop
A peptide library containing 164 fifteen-mer peptides spanning the entire ZIKV E protein was grouped into pools of five and used to identify linear epitopes targeted by mAbs A11 and A42 using an ELISA format (Fig. 3A). Pool 19 was found to have the highest binding activity to mAb A11. Individually, peptide 51 (151-MIVNDTGHETDENRA-165), located in Domain I, displayed the highest reactivity with mAb A11 (Fig. 3B).
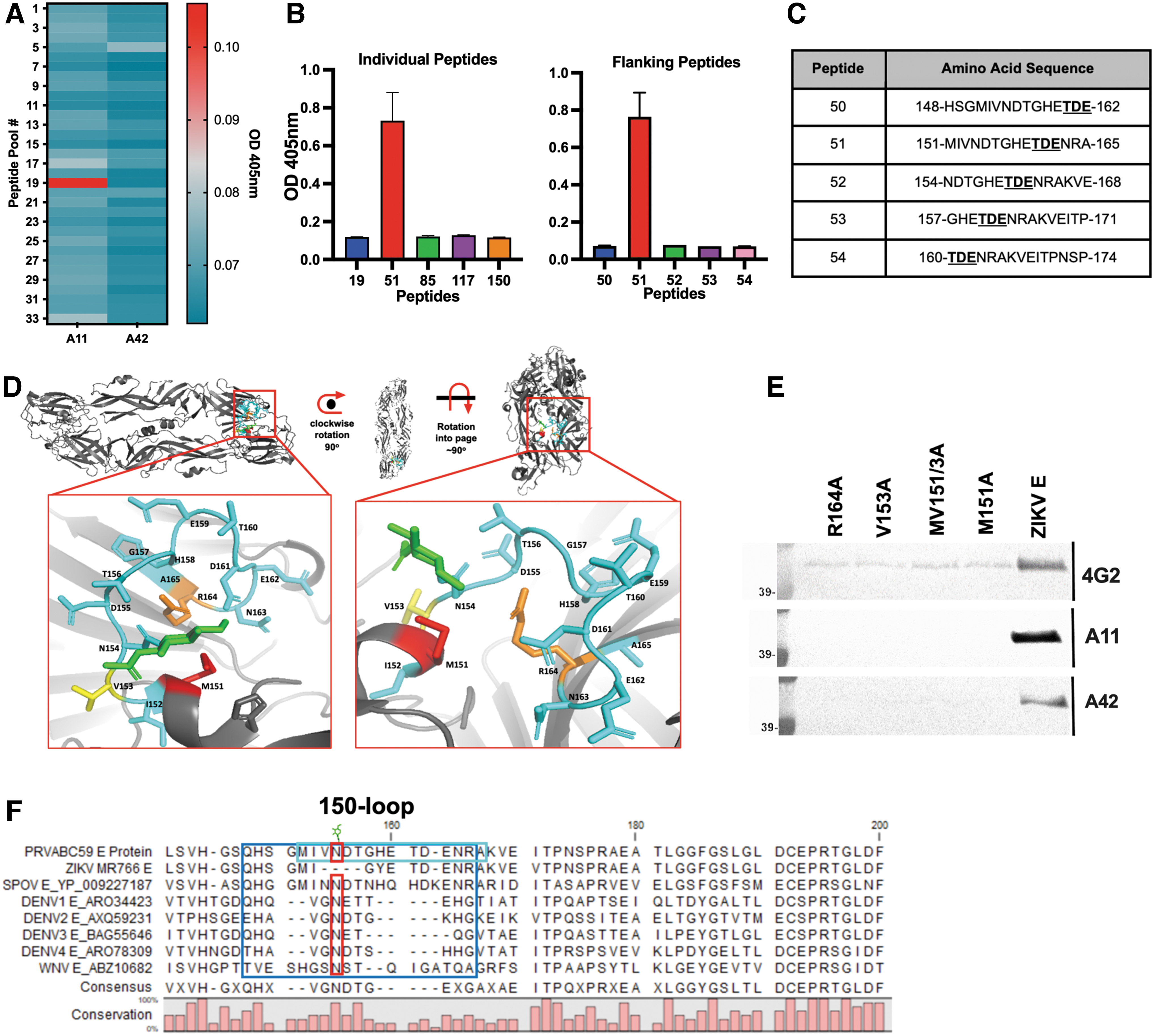
Epitope Mapping of mAb A11.
Interestingly, other peptides containing some of the same overlapping amino acids (151–165) (Fig. 3C) showed no reactivity to mAb A11 (Fig. 3B). The sequence of peptide 51 lies on the surface of ZIKV E. To reconfirm, three amino acids with bulky or positively charged functional groups (M151, V153, and R164) were mutated to alanine (Fig. 3D). E protein constructs with single- or double-mutations were transiently expressed as soluble proteins secreted into the cell culture supernatant. Western blots of the mutant proteins were probed using either mAbs 4G2, A11, or A42.
The lack of discernable reactivity of mAb A11 to all mutant ZIKV E supports our finding that the epitope is within the 150-loop or glycan-loop (Fig. 3E). Amino acid sequence alignments of various flavivirus envelope proteins reveal great diversity in the 150-loop region (Sirohi and Kuhn, 2017) and that the presence of the 151-MIVNDTGHETDENRA-165 peptide sequence is predictive of the interactions with mAb A11 (Fig. 3F).
Mouse/human chimeric cAb 42 maintains ZIKV specificity
Finally, for better utilization of the mAb A42, the variable domains of mAb A42 were cloned into HEK293T cells to produce a cmAb IgG. We determined that trans-species IgG switching did not affect the specificity of cmAb A42 with regards to reactivity to soluble WNV, DENV-2, and DENV-3 E by western blotting or our microsphere immunoassay (MIA) (Fig. 4A, B).
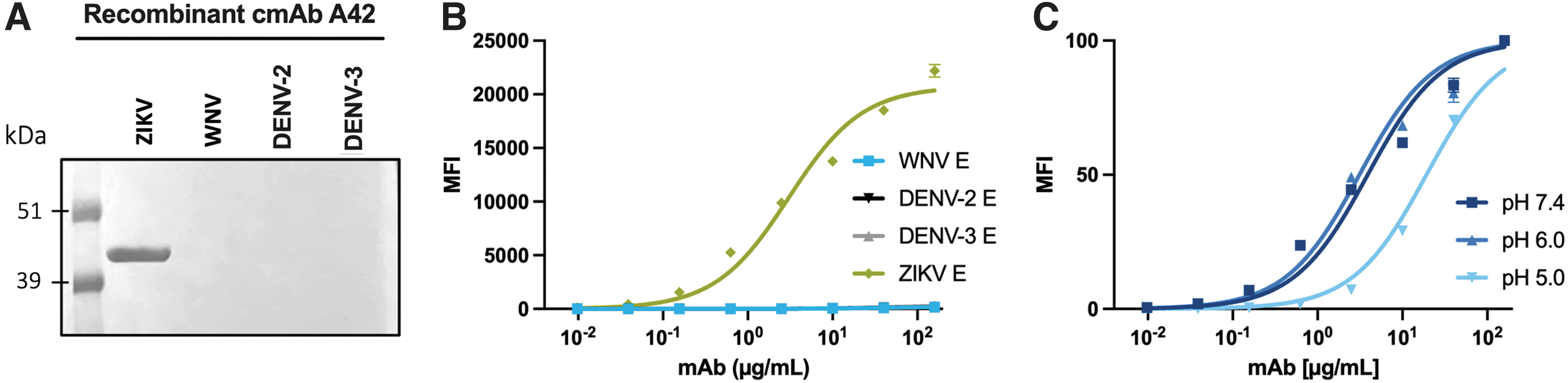
Binding Characteristics of Recombinant cmAb A42
However, the EC50 did increase to 3.0 (95% CI 2.0–4.6) μg/mL (Fig. 4B) and unlike the hybridoma-produced mAb A42, chimeric A42 had reduced affinity for ZIKV E at pH 5.0 (Fig. 4C), suggesting that chimeric expression changes the molecular interactions slightly. Regardless, the most unique aspect of mAb A42, the ZIKV E specificity, was preserved.
Discussion
We have generated two hybridoma cell lines that produce highly ZIKV E-specific mAbs, A11 and A42, recognizing a unique epitope in Domain I. Unlike most of the anti-flavivirus E mAbs, which are cross-reactive, both antibodies recognize only ZIKV E, highlighting their utility for cell culture-based diagnostic purposes, individually or alongside NS1 protein detection.
Both mAbs from our study are non-neutralizing, thus distinguishing them from other mAbs of the same class (Qu et al., 2020), and similarly were generated by immunizing soluble ZIKV E in mice, suggesting that recombinant monomeric E may expose vulnerable sites otherwise obscured from antibody recognition on mature virus particles. This may explain why this antibody class was not previously isolated from ZIKV-convalescent individuals (Barba-Spaeth et al., 2016; Stettler et al., 2016; Zhao et al., 2016).
In our analysis, mAb A11 has the stronger affinity; however, acidic environments decrease the affinity more for mAb A11 than for 4G2 or A42, indicating that mAb A11 interaction is pH dependent. Within the E protein epitope (151-MIVNDTGHETDENRA-165), only amino acid H158 is predicted to be protonated at low pH. Since this region is positively charged due to the R164 located in this region (Dai et al., 2016), protonation might inhibit mAb A11 from binding antigen in acidic environments (Haslwanter et al., 2017).
The lack of interaction with linear peptides and no binding to the glycan-loop regional mutants suggest that mAb A42's epitope is conformational. In addition, there is a reduction in binding at pH 5.0 with the recombinant cmAb A42 compared with hybridoma-produced antibody, which may be attributed to the differences in the constant regions between the native mouse mAb and the chimeric, human-derived antibody construct.
In conclusion, mAbs A11 and A42's specificity for ZIKV E highlights a unique epitope not commonly targeted by the humoral response to ZIKV infection. The specificity of both mAbs to discriminate against other flavivirus makes these antibodies highly useful for the development of diagnostic orthogonal virus-detection assays and a valuable resource for laboratory and molecular techniques.
Data Availability
The nucleotide and protein sequences of the cmAb A42 heavy and light IgG1 chains were deposited into GenBank under the accession numbers: OR260422 and OR260423.
Footnotes
Acknowledgments
The authors thank Wei-Kung Wang from the University of Hawaiʻi at Mānoa and David Clements from Hawaiʻi Biotech, Inc., for providing reagents used in this study. They also thank Michael Lieberman and Susan Hettenbach for their edits on this manuscript and acknowledge the WRCEVA for providing the SPONV. The following reagent was obtained through BEI Resources, NIAID, NIH: Peptide Array, ZIKV, PRVABC59 Envelope Protein, NR-50553.
Authors' Contributions
A.R.M., B.K.H., A.T., A.G., J.M.B., and A.T.L. conceived and designed the experiments. A.R.M., B.K.H., A.T., Y.-J.S.H., D.L.V., A.G., J.M.B., and A.T.L. coordinated the projects. A.R.M., B.K.H., A.T., L.M., L.O.M., T.A.S.W., E.N., D.A., and Y.-J.S.H. collected samples and performed the experiments. A.R.M., B.K.H., A.T., L.M., and C.-Y.L. performed the data analysis. A.R.M., B.K.H., A.T., L.M., and A.T.L. wrote the article. A.T. revised the article. All authors contributed to the article and approved the submitted version.
Author Disclosure Statement
The authors have no conflicts of interest to declare. All co-authors agree with the contents of the manuscript and there is no financial interest to report.
Funding Information
This project was supported by the National Institute of General Medical Sciences (NIGMS) of the NIH, IDeA Networks of Biomedical Research Excellence (INBRE) to support Undergraduate research (P20GM103466). Additional support was provided by the State of Kansas National Bio and Agro-defense Facility (NBAF) Transition Fund. A.T. was supported by the Fogarty International Center of the NIH under (D43TW009345) awarded to the Northern Pacific Global Health Fellows Program.