
Abstract
The SARS-CoV-2 pandemic has confirmed that the ability to rapidly mutate may be extremely beneficial for a virus. Not long after the first wave, new variants emerged with altered infectivity, disease severity, and mortality. These new strains most notably had numerous mutations of the spike (S) protein, a surface protein responsible for binding to and entering the host cell. The Delta and Omicron strains demonstrated increased immune evasion and improved binding affinity to the host cell receptor, angiotensin-converting enzyme 2 (ACE2). This study examines the ability of wild-type SARS-CoV-2 IgG to bind Delta and Omicron antigens, as well as their functional binding capabilities to two different S-ACE2 complexes. Twenty SARS-CoV-2 positive samples from patients who had recovered from infection with ancestral SARS-CoV-2 in the first wave of COVID-19 and 10 pre-pandemic control samples were studied. SARS-CoV-2 exposed patients showed significantly higher levels of IgG to SARS-CoV-2 S1/RBD (p < 0.001), N protein (p < 0.001), and Omicron spike variant (p = 0.01), but not to Delta spike variant (p = 0.966) when compared with controls. Furthermore, patient samples showed significantly greater inhibition of SARS-CoV-2 S1/RBD and E484K spike to ACE2 binding (p < 0.001 and p = 0.015, respectively). Conversely, there was no correlation between the binding inhibition of S1/RBD and E484K spike to ACE2 receptor. This study shows there is considerable cross-reactivity of IgG generated by wild-type SARS-CoV-2 infection to the Delta and Omicron variants.
Introduction
Named after their solar corona-like appearance, coronaviruses are covered in 100 surface projections ∼18 kDa in length made up of spike (S) glycoproteins, which are responsible for virus-cell membrane binding through the angiotensin-converting enzyme 2 (ACE2) receptor (Kathiravan et al., 2021). The ACE2 cell receptor is a surface protein that is found on many different types of cells throughout the body, but especially in the lungs, kidneys, intestines, brain, and other capillary-rich organs (Samavati and Uhal, 2020).
The S protein is a type-1, surface-anchored membrane protein that forms a trimer crucial in host cell recognition and entry (Zhang et al., 2021). The discontinuous manner in which coronaviruses transcribe RNA results in the rapid accumulation of recombination events, insertions, deletions, and point mutations (Carabelli et al., 2023). These S protein mutations have repeatedly been shown to alter infection rates, vaccine effectiveness, affinity to ACE2, and disease severity and outcomes (Singh et al., 2021). In fact, all the SARS-CoV-2 variants that have been declared variants of concern by the WHO have at least one mutation of the S protein (Khamees et al., 2022).
The Delta variant was first detected during the second SARS-CoV-2 wave in India in October 2020. Also known as B.1.617.2, this variant had increased neutralizing antibody evasion and infectivity, attributable to the L452R, TK748K, and P681R mutations on the S protein (Dhawan et al., 2022). When compared with the original Wuhan strain, the Delta variant was 97% more transmissible and had both increased hospital admission and mortality rates (Campbell et al., 2021).
The Omicron variant, B.1.1.529, was first identified in November 2021 in South Africa and Botswana. As with the Delta variant, the L452R is present on the Omicron S protein, in addition to many other clinically important mutations, in particular, E484K (Das et al., 2022). This mutation reduced the immune response to the virus while simultaneously increasing the binding affinity of the receptor (Wang et al., 2021).
The present study primarily investigates the functional capabilities and cross-protection between SARS-CoV-2 IgG in individuals infected by the wild-type virus in the first wave of the pandemic and the Delta and Omicron variants. Second, we examine changes in the binding inhibitory function of the antibodies to the S-ACE2 complex formation of the wild-type S1/RBD and E484K spike variants.
Materials and Methods
Samples
An anonymized set of 30 serum samples was purchased from RayBiotech® (RayBiotech, Georgia, USA). Twenty samples were acquired from recovered patients who were infected during the first SARS-CoV-2 wave in 2020 with the wild-type virus, collected between April and August 2020. Ten samples from healthy adults taken prior to the pandemic were also included as a control group. Ethical approval for the project was obtained from the University College Dublin Human Research Ethics Committee (LS-E-22-53-Johnson-Hassan).
ELISA for determination of anti-SARS-CoV-2 IgG
The anti-SARS-CoV-2 QuantiVac enzyme-linked immunosorbent assay IgG (Euroimmum, Lübeck, Germany) quantitative assay was used to determine the concentration of human IgG to the SARS-CoV-2 S1/RBD antigen using 96 wells precoated with the recombinant domain of SARS-CoV-2S protein.
Samples were initially diluted 1:101 in sample buffer, though some samples required further dilution. First, 100 µL of each calibration control and sample were added to each well and incubated for 1 h at 37°±1°C. The plate was then washed three times using a BioTek ELx50 automated plate washer (Agilent, California, USA) and 450 µL working strength wash buffer, leaving the wash buffer in the wells for 30–60 sec per cycle. After washing, the plate was blotted dry on tissue. Following this, 100 µL of peroxidase-labeled antihuman IgG (enzyme conjugate) was added and incubated at 37°C ± 1°C for 30 min. The plate was washed again, and 100 µL tetramethylbenzidine (TMB)/hydrogen peroxide (H2O2) chromogen/substrate solution was added before a final incubation for 30 min at room temperature, protected from direct sunlight. Prior to reading at 450 nm (reference wavelength 630 nm) on a BioTek ELx808™ absorbance plate reader (Agilent, California, USA), 100 µL of 0.5M sulfuric acid stop solution was added, producing a color change that directly relates to the concentration of antibody in the sample.
Measurement of IgG to N protein, Delta spike and Omicron spike proteins of SARS-CoV-2
Quantification of the IgG present against SARS-CoV-2N, Delta, and Omicron spike variant proteins in human serum was determined using the RayBio COVID-19 N Protein, Delta, and Omicron Spike Variant Human IgG ELISA Kits, respectively (RayBiotech).
These indirect ELISAs utilize a 96-well plate and follow the same assay principle until the final incubation step. The samples were diluted 1:1500 in diluent, and 100 µL of sample and controls were added to each well and incubated at room temperature for 1 h with gentle shaking. The plate was then washed four times using 300 µL of wash buffer on the BioTek ELx50 automated plate washer (Agilent, California, USA) and blotted on paper. Next, 100 µL biotinylated antihuman IgG was added and incubated for 30 min at room temperature with gentle shaking. The plate was washed as before, and 100 µL horseradish peroxidase (HRP)-streptavidin solution was added to each well and incubated at room temperature with gentle shaking. The plate was washed again and 100 µL TMB one-step substrate was added and incubated at room temperature in the dark with gentle shaking for 15 min (N protein and Delta spike assays) or 30 min (Omicron spike assay). Finally, 50 µL of 0.2M sulfuric acid stop solution was added, and the plate was read at 450 nm immediately on the BioTek ELx808 absorbance plate reader (Agilent, California, USA).
SARS-CoV-2 S1/RBD binding inhibition assay to ACE2
To determine the neutralizing antibody levels in each sample, the semiquantitative Euroimmun SARS-CoV-2 NeutraLISA assay was used (Euroimmum, Lübeck, Germany).
The samples were diluted 1:5 in sample buffer containing soluble biotinylated ACE2 prior to use and 100 µL of each of the diluted samples, and controls were added to the recombinant S1/RBD domain-coated wells. Following incubation for 1 h at 37°±1°C, the plate was washed three times on the BioTek ELx50 automated plate washer (Agilent, California, USA) using 450 µL of wash buffer, leaving the wash buffer in the wells for 30–60 sec per cycle, and tapped dry on tissue. Subsequently, 100 µL of the peroxidase-labeled streptavidin enzyme conjugate was added and incubated for 30 min at room temperature. The plate was washed as above. Next, 100 µL of the TMB/H2O2 chromogen/substrate solution was added to each well and incubated, protected from direct sunlight, for 15 min at room temperature. Finally, 100 µL of 0.5 M sulfuric acid stop solution was added, and the plate was immediately read at 450 nm, with a reference wavelength of 630 nm, on the BioTek ELx808 absorbance plate reader.
SARS-CoV-2 E484K spike variant ACE2 interaction inhibition assay
The binding inhibition of SARS-CoV-2 E484K S/RBD and ACE2 interaction was established using the RayBio COVID-19 Spike Variant-ACE2 Binding Assay Kit I (RayBiotech). This ELISA utilizes a SARS-CoV-2S/RBD(E484K)-coated 96-well microplate and samples diluted in purified recombinant ACE2 protein.
Each sample was tested at three different serial dilutions, 1:25, 1:250, and 1:2500, in addition to a positive control and a blank, both of which were tested in triplicate. First, 100 µL of the control, blank, and each sample dilution were added to the wells and incubated for 2.5 h at room temperature with gentle shaking. The plate was washed four times using 300 µL of wash buffer using the BioTek ELx50 automated plate washer (Agilent, California, USA) and blotted against tissue. Then 100 µL of the ACE2 detection antibody was added and incubated for 1 h at room temperature with gentle shaking. The wells were washed as above, and 100 µL of HRP-conjugated anti-IgG were added and incubated for 1 h at room temperature with gentle shaking. The washing step was repeated, and 100 µL of TMB one-step substrate was added to each well and incubated in the dark for 30 min with gentle shaking at room temperature. Finally, 50 µL of stop solution was added, and the plate was read immediately at 450 nm using the BioTek ELx808 absorbance plate reader.
Statistical analysis
Data generated in this study were statistically analyzed using IBM® SPSS® Statistics [Version 28.0.1.1 for macOS (14)] and Microsoft® Excel [Version 16.54 for macOS (21101001)]. The half maximal inhibitory concentration (IC50) values for the binding inhibition assay were calculated on GraphPad© Prism [Prism 10 for macOS, Version 10.0.1 (170)] using nonlinear regression analysis. Spearman’s Rank Order Correlation was used to test for correlation between the control and patient groups for each assay. To compare the male and female patient groups, Mann–Whitney U tests were performed. A p < 0.05 indicates statistical significance.
Results
Patient demographics
In the control group, 70% of the samples were from males, versus 45% in the patient group. The mean age ± standard deviation (SD) for the control group was 39.0 ± 16.4 years (range = 18–65 years). The mean age ± SD for the patient group was 57.0 ± 14.5 years (range =21–81 years). In the patient cohort, the range of days post onset of symptoms was 24–34 days (mean ± SD = 32.0 ± 2.8 days).
IgG levels to the SARS-CoV-2 S1/RBD and N protein
The positive samples for both the S1/RBD and N protein assays all showed great variation in IgG concentration. The concentration of human IgG to the SARS-CoV-2 S1/RBD antigen in the patient samples ranged from 17.74 to 426.87 RU/mL, with all testing IgG positive. Interestingly, three patients had low to no IgG levels when tested on the N protein ELISA, with an overall range of 0 − 34.38 RU/mL. All 10 control samples were negative for both assays. The results for both assays showed significant differences when the control and patient groups were compared (p < 0.001), as shown in Figure 1.
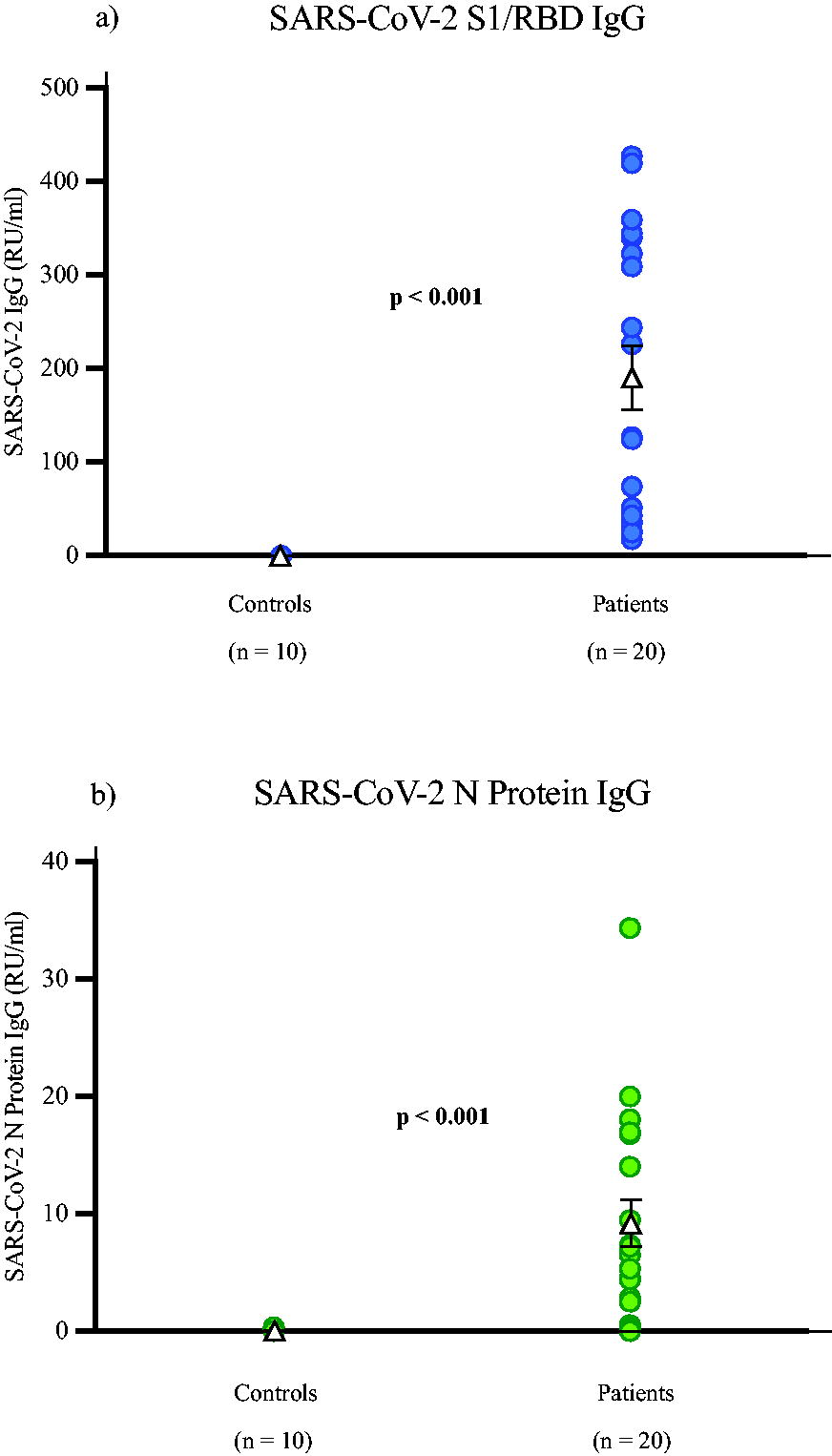
The concentration of SARS-CoV-2 IgG levels to the SARS-CoV-2
IgG levels to the SARS-CoV-2 Delta and Omicron spike variants
Of the control cohort, three were negative for the Delta variant, and four individuals had IgG levels over 100 RU/mL, as seen in Figure 2. The patient group showed 70% were positive; however, two of these patients had IgG concentration that were very weakly positive, below 16 RU/mL. The positivity cutoff point of the assay is >15 RU/mL. For the Omicron assay, only one control sample tested positive at 25.86 RU/mL. Of the patient group, 50% were positive for IgG against the SARS-CoV-2 Omicron spike variant. When comparing the control and patient cohorts, the results showed a significant correlation (p = 0.01) for the Omicron Spike Variant but not for Delta (p = 0.966).
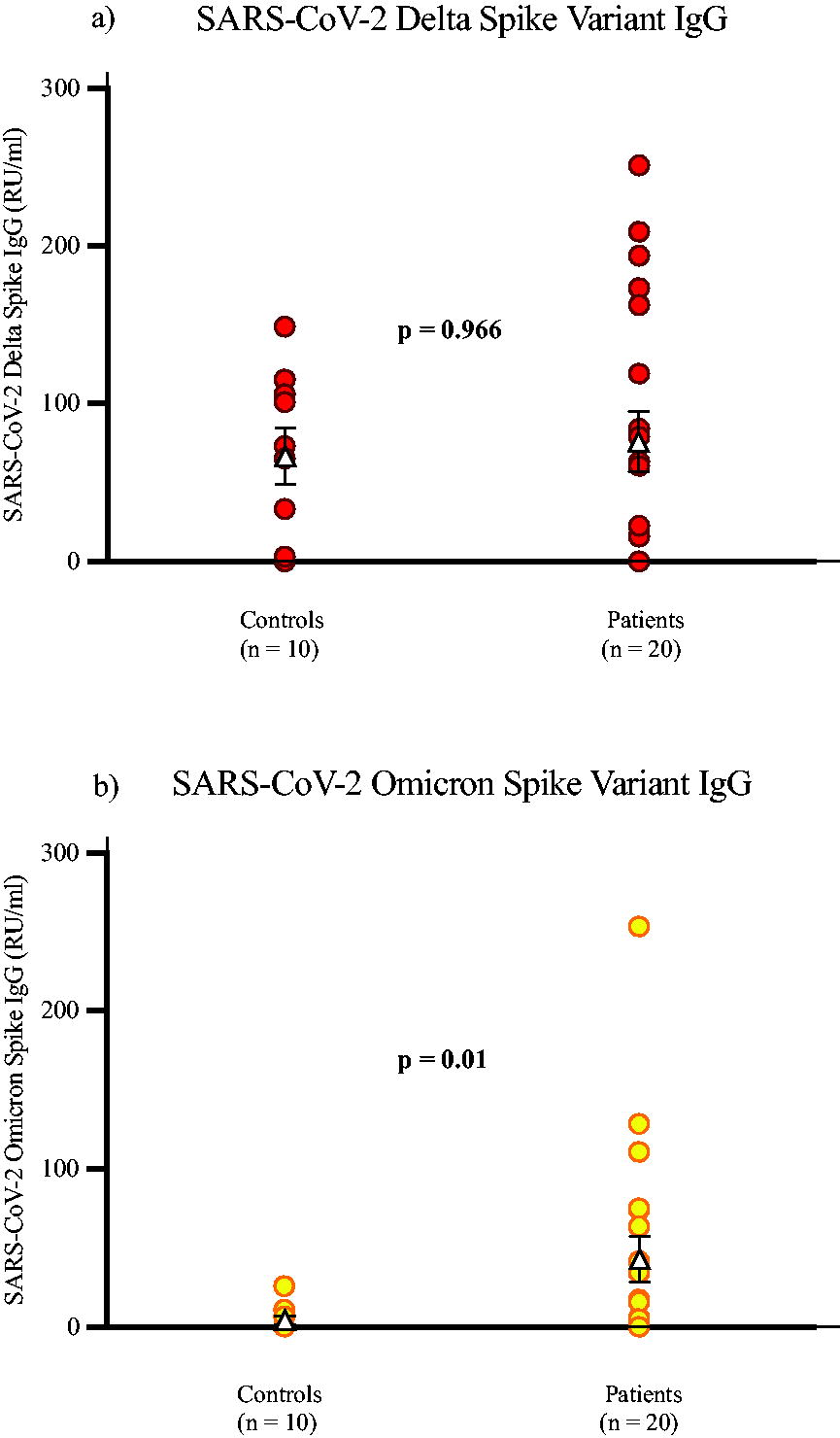
The concentration of control and patient samples’ respective SARS-CoV-2 IgG levels from the SARS-CoV-2
Additional Spearman’s Rank Order Correlation analysis was performed to compare the N protein, Delta spike, and Omicron spike to the SARS-CoV-2 S1/RBD reactivity in the convalescent patient group. No significant correlations were observed (p = 0.086, 0.2, and 0.405, respectively).
Determination of binding inhibition of SARS-CoV-2 E484K spike variant to ACE2
Nonlinear regression calculations were performed to generate the IC50 from the percentage binding inhibition values acquired from the E484K binding inhibition assay. An IC50 curve was acquired for each sample, showing the binding inhibition drop exponentially with each serial dilution. A representative result from a control and patient can be seen in Figure 3. These IC50 values were then used in the subsequent correlation analysis performed on the control and patient groups for each assay [mean = 1576.02, standard error of mean (SEM) ± 1878.55]. For the S1/RBD inhibition assay, all of the control samples were negative, 18 of the patient samples were positive, and 2 were equivocal. Among these positive samples, there was a wide range in the inhibition percentage, from 42.24% to 98.47% (mean ± SEM = 73.90 ± 25.94), as seen in Figure 4. However, the binding inhibition percentages from the assays evaluating binding capabilities were not statistically significant (p = 0.539). Correlation analysis of the SARS-CoV-2 S1/RBD and E484K variant binding inhibition assays showed a significant correlation between previous wild-type infection and inhibition of ACE2 binding (p < 0.001 and p = 0.015, respectively).

Nonlinear regression analysis of the percentage binding inhibition curves for a representative control sample (squares) and positive sample (circles), calculated using GraphPad© Prism. The serum dilution titers used in this assay were 1:25, 1:250, and 1:2500. The IC50 values for the control and patient shown were 22.81 and 341.30, respectively. IC50, half maximal inhibitory concentration.
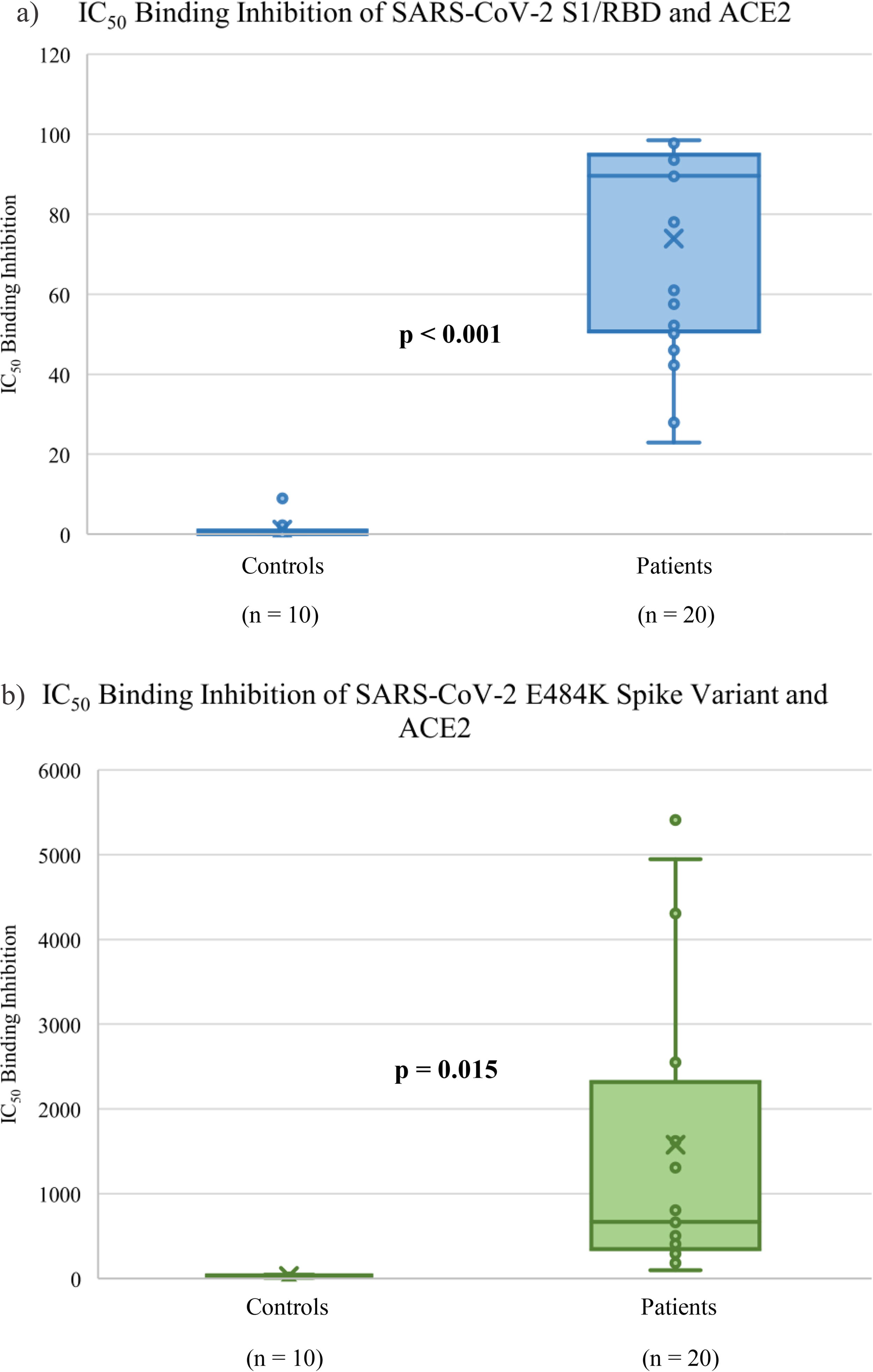
Box and whisker plot of the IC50 of control and patient samples’ respective SARS-CoV-2 binding inhibition levels to ACE2 from the SARS-CoV-2
Sex comparison
There was no significant correlation for biological sex for any of the assays performed in this study. The SARS-CoV-2 S1/RBD, N protein, Delta, and Omicron spike variant IgG assays had p values of 0.909, 0.224, 0.095, and 0.621, respectively, while the S1/RBD binding inhibition assay had a p-value of 0.470, and the E484K binding inhibition assay had a p-value of 0.271. When comparing the sex results for each quantitative assay, frequency analysis was performed on the patient groups. Notably, the male groups showed less cross-protection in every assay except for Omicron spike variant. The largest difference between male and female positivity was seen for the Delta spike variant, with 35.7% of males and 64.3% of females being positive. Interestingly, the S1/RBD and N protein assays had the same positivity rates for both sexes.
Discussion
Natural infection with SARS-CoV-2 is denoted by antibody reactivity to the N protein only, as it is the S protein that is used in the vast majority of SARS-CoV-2 vaccines. The N protein comprises the highest abundance in the SARS-CoV-2 virion and is essential for the packaging of the genomic RNA into the ribonucleotide complex (RNP) (Wu et al., 2023). In addition, a 2023 study showed that the N protein is capable of transferring RNA into cells through six-transmembrane epithelial antigen of the prostate-2 (STEAP2) receptor-mediated endocytosis, entirely independent of the ACE2 receptor (Wu et al., 2023). However, of the patient cohort with N protein assay results, three had undetectable IgG concentrations. This is interesting, as it is indicative of vaccine protection rather than natural infection, but the samples for this study were collected before SARS-CoV-2 vaccination began and thus cannot exhibit protection from a vaccine.
A significant correlation between previous natural infection with the wild-type virus and reactivity to the test antigen was present in all quantitative SARS-CoV-2 IgG assays except the Delta spike variant. This may be due to a number of reasons.
First, the specificity and performance of the RayBio Delta Spike Variant assay may not have been as good as the other assays of this study. Considerable positivity is evident in the control group for the Delta variant assay that is not evident in the Omicron and N protein. The Delta assay resulted in 70% of the control group being positive for SARS-CoV-2 Delta spike IgG. Second, the Delta variant may not gain any significant cross-protection from antibodies to the wild-type SARS-CoV-2. A 2022 study showed that there was a larger increase in Omicron neutralization in vaccinated individuals than Delta (Khan et al., 2022). However, a 2023 meta-analysis found that there was >85% cross-protection (on average) across the Alpha, Beta, and Delta spike variants, with the Omicron variant only showing <55% protection (Stein et al., 2023). Third, there is a possibility that members of the control group had been exposed to one of the other circulating seasonal coronaviruses prior to sample collection, offering some cross-protection and thus cross-reactivity in the test. Globally, four different seasonal coronaviruses circulate continuously that are capable of infecting humans, HCoV-HKU1, HCoV-NL63, HCoV-229E, and HCoV-OC43 (Sayama et al., 2023). It has been reported that coronavirus antibodies may be present in as much as 90% of adults (Theel et al., 2020).
It had been shown that males mount a significantly lower immune response to vaccines and viral infections (Jacobsen and Klein, 2021). In addition, males have also been shown to be three times more likely to be admitted to the intensive care unit and 16% more likely than females to suffer mortality from a SARS-CoV-2 infection (Chaturvedi et al., 2022). The change in severity and outcome of the infection is a result of the differences in the factors of innate immunity, hormones, and finally the sex chromosomes (Conti and Younes, 2020). Interestingly, in the present study, all quantitative IgG assays showed higher levels in the female patient group except for the Omicron spike variant, where the inverse of this pattern is evident. However, the small sample size of this study is the likely reason for this. In a 2020 study, it was shown that though men experience more severe disease outcomes, males and females are in fact equally susceptible to contracting COVID-19 (Jin et al., 2020). Furthermore, a 2023 study of Omicron infections in China also demonstrated greater disease severity in male patients (Miao et al., 2023).
Additionally, a 2021 study showed that recovered COVID-19 patients exhibit varying antibody levels and neutralizing titers, supporting the findings of the current study (Carrillo et al., 2021). These results strongly suggest that there is a relationship between previous SARS-CoV-2 infection and the neutralizing effect of the antibodies present. In contrast to the SARS-CoV-2 S1/RBD inhibition assay, the control group when tested using the E484K assay did show varying degrees of binding inhibition. Furthermore, 55% of the patient samples showed between 80% and 100% inhibition when using the S1/RBD assay, but the results for the E484K assay were generally much lower. This is likely due to poor assay specificity, as the E484K mutation did not emerge until the SARS-CoV-2 beta variant of September 2020 (Wolf et al., 2023). However, a significant correlation was still evident between the control and patient group. It has been shown that the presence of the E484K mutation reduces the neutralizing effects of antibodies in convalescent and vaccinated individuals (Jangra et al., 2021).
In conclusion, antibodies to the SARS-CoV-2 wild-type strain do offer significant cross-protection to the Omicron spike variant, but further analysis would be required to determine the levels of cross-protection for the Delta variant. This is due to the lack of specificity of the assay used to test for IgG against the Delta spike. In addition, this study showed that the presence of IgG in SARS-CoV-2 does increase inhibition of S-ACE2 binding to the wild-type virus. Finally, sex analysis showed that convalescent males exhibit less cross-protection to the Omicron variant than females, but not to the Delta variant.
Author Disclosure Statement
No competing financial interests exist.
Footnotes
Acknowledgments
The author would like to thank the National Virus Reference Laboratory for their support and funding of this project.
Authors’ Contributions
L.J.: Formal analysis (lead), investigation (lead), writing—original draft (lead), and visualization (lead). C.F.D.G.: Writing—review and editing (support). J.H.: Conceptualization (lead), methodology (lead), funding acquisition (lead), writing—review and editing (lead), and supervision (lead).
Funding Information
This project was funded by the National Virus Reference Laboratory.